
覆土控鞭高品质竹笋栽培对高节竹叶片形态和养分化学计量特征的影响
2017-11-27 07:22:00江志标陈双林郭子武杨清平李明良
浙江农林大学学报 2017年6期
江志标,陈双林,郭子武,杨清平,朱 玲,李明良
(1.浙江省桐庐县农业和林业技术推广中心,浙江 桐庐311500;2.中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400)
覆土控鞭高品质竹笋栽培对高节竹叶片形态和养分化学计量特征的影响
江志标1,陈双林2,郭子武2,杨清平2,朱 玲1,李明良1
(1.浙江省桐庐县农业和林业技术推广中心,浙江 桐庐311500;2.中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400)
高节竹Phyllostachys prominens是优良的笋材兼用竹种,生态适应性强。采取覆土控鞭经营措施能显著提高高节竹笋外观品质、营养品质和食味品质及竹林经济效益。为摸清覆土控鞭栽培对高节竹生长的影响,为高节竹林高效可持续经营提供参考,测定了覆土控鞭栽培与对照高节竹林1~3年生立竹叶片的主要叶性因子和碳(C),氮(N),磷(P)质量分数,分析高节竹叶片形态和养分化学计量特征对覆土控鞭栽培的响应规律。结果表明:覆土控鞭栽培2 a后,高节竹1~3年生立竹叶片变细长,单叶面积和比叶面积略有增大,单叶质量和叶片厚度稍有下降,叶片形态指标覆土控鞭栽培和不覆土栽培竹林间差异均不显著(P>0.05)。覆土控鞭栽培的高节竹林1~3年生立竹叶片碳质量分数显著升高(P<0.05),氮略有升高(P>0.05),磷稍有下降,C/N和N/P均有所升高(P>0.05),而C/P显著升高 (P<0.05)。覆土控鞭栽培对高节竹叶片形态并未产生明显的影响,对高节竹光合碳同化能力和氮、磷养分利用效率有促进作用,可见高节竹林采取科学的覆土控鞭高品质竹笋栽培措施能达到高效可持续经营的目标。表3参21
森林培育学;高节竹;覆土控鞭栽培;叶片性状;化学计量
叶片性状包括叶片的结构性状和功能性状,是由遗传因素和环境条件共同决定的。不同生境的植物叶片性状间的差异体现了植物对环境的适应性[1-2]。叶片重要的性状指标能反映出植物偿还叶片养分和干物质投资时间的快慢[3],如比叶面积较小的植物叶片会将能量和物质用来构建叶片而非投资到光合器官,因而含有更多的厚壁组织和细胞壁成分[4-6],这势必引起叶片碳同化能力的下调。然而,叶片碳的储存在一定程度上受控于氮、磷可获得的量[7-8],碳/氮(C/N)和碳/磷(C/P)的比值更是在一定程度上反映了植物的养分利用效率,氮/磷(N/P)比则可以作为对植物生长起限制性作用的养分元素的指示剂[9-10]。因此,研究植物叶片性状和养分化学计量与环境和人工经营干扰的关系,有助于理解植物对环境的适应机制及对资源的利用和分配过程等[11],具有重要的科学价值和生产指导意义。高节竹Phyllostachys prominens俗称钢鞭哺鸡竹,是优良的笋材兼用竹种,具有竹笋产量高、品质佳、加工性能好,竹材径级较大、材质坚硬,生态适应性强,地下鞭系粗壮、延伸生长能力强等特点,在浙江省杭州市、湖州市等地广为栽培。高节竹出笋期4月中下旬至5月中旬,较雷竹Phyllostachys violascens和毛竹Phyllostachys edulis出笋迟,由于竹笋消费疲劳的原因,高节竹林经济效益一直以来都表现不佳。为顺应市场对高品质竹笋的大量需求,根据高节竹的生物学和生态学特性,浙江省杭州市桐庐县高节竹主产区开展了高节竹林地覆土控鞭高品质竹笋栽培措施的规模化应用。覆土控鞭栽培措施实施后高节竹笋期推迟约15 d,竹笋外观品质、营养品质和食味品质明显改善,可食率提高,竹笋香甜味增加,酸涩味及粗糙度降低[12],深受上海、杭州等地的消费者欢迎,竹笋供不应求,竹林经营效益显著提高,推广规模日趋增大。高节竹林地覆土控鞭栽培后,林分结构可以通过留笋养竹和伐竹等人工调控措施来维持丰产结构要求,但竹林地下部分的生长环境发生了变化,这是否会对高节竹立竹叶片形态和养分化学计量特征产生影响,进而影响高节竹生长呢?为此,开展了覆土控鞭栽培2 a和不覆土栽培的高节竹林1~3年生立竹叶片主要形态指标和碳、氮、磷质量分数及化学计量比的比较研究,旨在为高节竹林高效可持续经营提供参考。
1 材料与方法
1.1 试验地概况
试验地位于浙江省杭州市桐庐县(29°35′~30°05′N, 119°11′~119°58′E)莪山乡, 属亚热带季风气候区,四季分明,年平均气温为16.6℃,极端高温为41.7℃,极端低温为-9.5℃,全年≥10℃的积温为5 262.0℃,年平均无霜期为252.0 d,年平均降水量为1 462.0 mm,年平均蒸发量为1 385.0 mm,年平均相对湿度为81%。土壤为红壤,土层厚度80 cm以上。高节竹资源丰富,全乡有高节竹林0.14万hm2,是浙江省 “高节竹之乡”。以高节竹资源为主的竹产业已成为莪山乡农业支柱产业和特色产业。近年来,为提高高节竹竹笋品质和经济效益,规模化推广应用高节竹覆土控鞭栽培技术措施,具体方法为9月至翌年2月利用建房、林道修建等的土方,在具有丰产林分结构的高节竹林中均匀地添加团聚体结构好、土壤黏粒50%~70%,容重1.2 g·cm-3左右,pH 4.5~5.5,去除了土中石块、树蔸等的黄壤和红壤客土30 cm左右,覆土后前2 a的6月、10月挖除覆土层土壤中的竹鞭,覆土后第3年覆土层留鞭,实行季节性施肥、林地垦复和林分结构调控。1次覆土可维持3 a的高节竹高品质竹笋生产,经济效益较不覆土栽培提高4倍以上。
1.2 试验方法
试验林立地条件基本一致,土壤为红壤,pH 5.2,有机质质量分数为10.71 g·kg-1,全氮1.37 g·kg-1, 全磷 0.839 g·kg-1, 全钾 8.93 g·kg-1。 2012年 6-7月结合林地垦复, 撒施复合肥 450 kg·hm-2, 9-10月覆土控鞭栽培前撒施复合肥750 kg·hm-2。2014年6月分别选择面积不少于0.2 hm2的覆土控鞭栽培(覆土2 a)和不覆土栽培高节竹试验林各1块。在每块试验林内,距离林缘10 m处设置10 m×10 m样地3个,样地间距10 m,并调查试验林立竹密度、立竹胸径、立竹年龄结构等林分结构状况。覆土控鞭栽培和不覆土栽培高节竹试验林立竹密度分别为(11 475±465)株·hm-2和(11 310±405)株·hm-2,胸径分别为(5.85 ± 0.43)cm 和(5.75 ± 0.37)cm, 1年生竹∶2年生竹∶3年生竹数量比分别为 3.9∶3.8∶2.3和3.8∶3.7∶2.5。
随机选取1~3年生标准立竹各6株·样方-1,分别取竹冠上部、中部和下部叶片各20片的均匀混合后取20片,作为一标准竹叶片样品,然后将6株标准竹叶片混合后,从中取20处作为该样方该标准竹叶片样品,即1~3年生立竹叶片共180片·处理-1,用于测量叶片的叶长、叶宽、厚度、单叶面积、单叶质量, 计算比叶面积[比叶面积(cm2·g-1)=单叶面积(cm2)/单叶质量(g)]和叶形指数(叶长/叶宽)。 另将每标准竹剩余叶片混合作为一个样品,共计18个样品,用于测定叶片碳、氮、磷质量分数,并计算养分化学计量比。叶片碳采用重铬酸钾氧化法测定,氮采用凯氏定氮法测定,磷采用钼锑抗比色法测定[13]。
1.3 数据分析
试验数据在Excel 2003统计软件中整理和图表制作,在SPSS 17.0统计软件中进行单因素方差分析和0.05水平的最小显著差法(LSD)多重比较,分析覆土控鞭栽培和不覆土栽培高节竹林1~3年生立竹叶片主要形态指标和碳、氮、磷质量分数及化学计量比之间的差异。试验数据均表示为平均值±标准差。
2 结果与分析
2.1 覆土控鞭栽培对高节竹叶片形态特征的影响
由表1可知:随立竹年龄增加,除比叶面积外,高节竹主要叶性因子均总体呈增加趋势,且2年生、3年生立竹显著高于1年生立竹(P<0.05),而前两者间并显著差异;比叶面积则相反,为1年生立竹显著高于2年生、3年生立竹,且后两者间仅略有差异(P>0.05)。
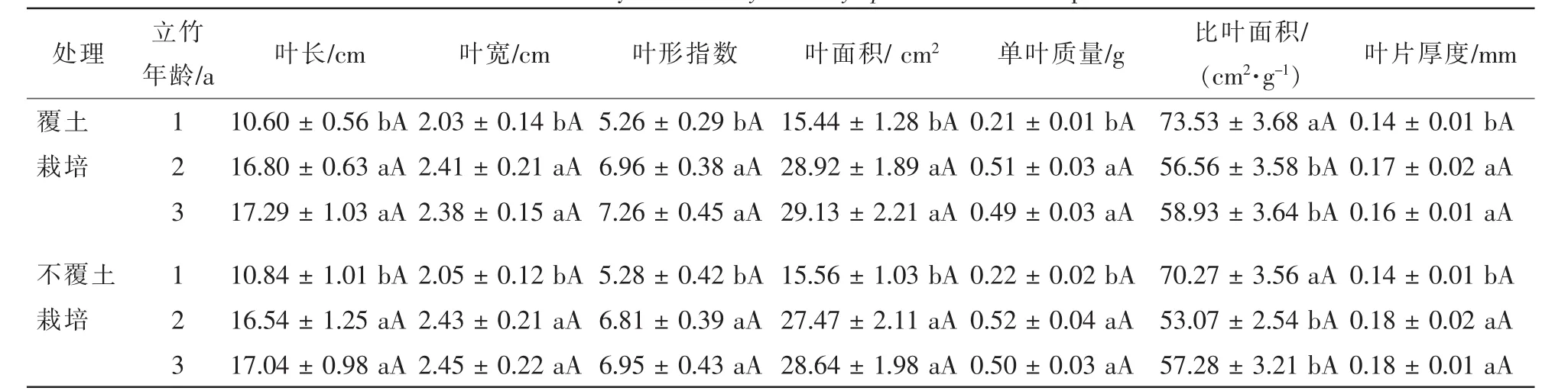
表1 覆土控鞭栽培高节竹叶片形态特征Table 1 Leaf traits of 1-3-years-old Phyllostachys prominens from experimental stand
覆土栽培2 a后,高节竹1年生立竹叶长稍有降低,而2~3年生立竹叶长有小幅度增加,1~3年生立竹叶宽均有小幅度下降,1~3年生立竹叶片叶长和叶宽的变化使叶形指数总体上有小幅度升高,叶长、叶宽和叶形指数覆土控鞭栽培和不覆土栽培竹林间均无显著差异(P>0.05);覆土栽培2 a后高节竹1~3年生立竹叶片单叶面积和比叶面积总体上均有一定幅度的升高,而单叶质量和叶片厚度总体上下降,单叶面积、单叶质量、比叶面积和叶片厚度覆土控鞭栽培和不覆土栽培竹林间也均无显著差异(P>0.05)。
上述分析表明:虽然覆土控鞭栽培后高节竹的地下鞭系生长环境发生了明显的变化,特别是温度和土壤水分,但对不同年龄立竹的叶片形态特征并没有产生明显影响,体现出高节竹很强的生态适应性。
2.2 覆土控鞭栽培对高节竹叶片碳、氮、磷质量分数的影响
随立竹年龄增加,高节竹叶片碳含质量分数总体升高,1年生立竹碳显著低于2年生和3年生立竹(P<0.05),且后两者间并无显著差异,而氮、磷总体下降,1年生立竹氮、磷显著高于2年生和3年生立竹,且后两者间亦无显著差异(P>0.05)(表 2)。覆土控鞭栽培2 a后,高节竹1~3年生立竹叶片碳质量分数均显著升高,增幅分别为6.62%,9.25%和6.09%,差异达显著水平(P<0.05);覆土栽培2 a后高节竹1~3年生立竹叶片氮质量分数也有不同幅度的升高,增幅分别为4.12%,2.51%和1.99%,而叶片磷质量分数均有一定幅度的下降,降幅分别为0.55%,2.89%和0.75%,但1~3年立竹叶片氮、磷质量分数覆土控鞭栽培与不覆土栽培高节竹林间均无显著差异(P>0.05)。可见,高节竹林覆土控鞭栽培后对叶片光合产物碳会产生明显的影响,竹林光合碳同化能力可能得到提高,但从土壤中吸收转运氮、磷养分并没有明显的变化。
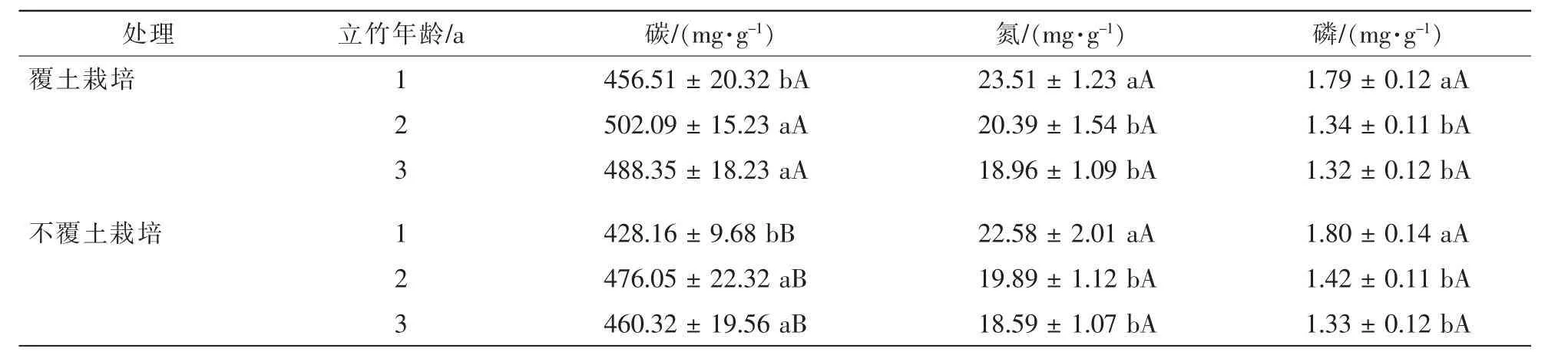
表2 覆土控鞭栽培高节竹叶片碳、氮、磷质量分数Table 2 Leaf C,N,P content for 1-3-year-old Phyllostachys prominens of experimental stand
2.3 覆土控鞭栽培对高节竹叶片碳、氮、磷化学计量比的影响
由表3可知:随立竹年龄增加,高节竹1~3年生立竹叶片碳/氮(C/N)比和碳/磷(C/P)比均总体呈升高变化趋势,2年生和3年生立竹叶片C/N和C/P匀显著高于1年生立竹(P<0.05),且2年生和3年生立竹的C/N和C/P并无显著差异(P>0.05);叶片氮/磷(N/P)比虽亦呈升高变化规律,但立竹年龄间差异不显著(P>0.05)。覆土控鞭栽培2 a后,高节竹1~3年生立竹叶片C/N和N/P均有所升高,增幅分别为2.43%,2.90%,3.99%和4.87%,5.55%,2.79%,但与不覆土栽培高节竹林比较并无显著差异,而高节竹林1~3年生立竹叶片C/P覆土栽培后均有显著提高,增幅分别为7.32%,8.32%和11.06%,覆土控鞭栽培和不覆土栽培高节竹林之间差异均达显著水平(P<0.05)。说明覆土控鞭栽培并没有影响到高节竹的土壤氮、磷养分吸收转运,而且一定程度上提高了高节竹的氮、磷养分利用效率。
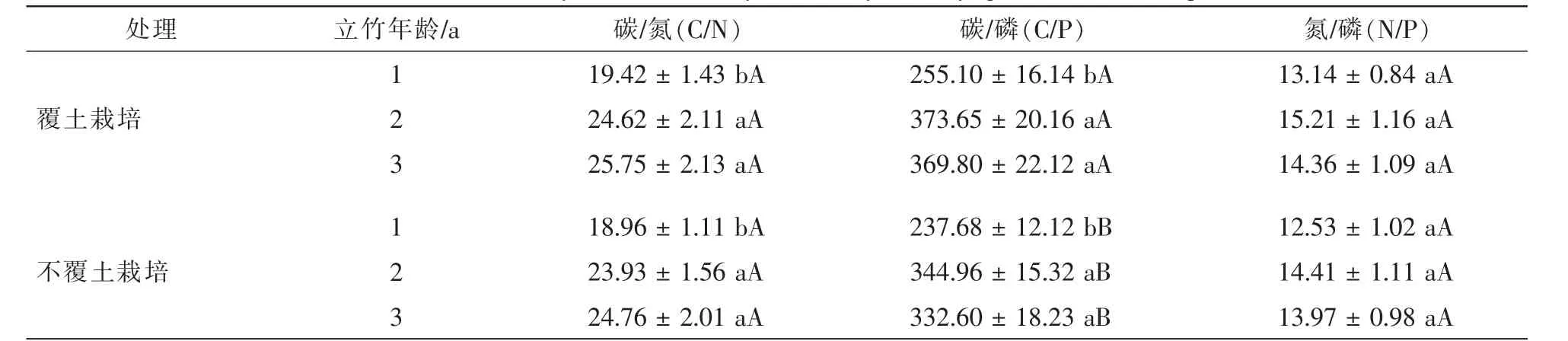
表3 覆土控鞭栽培高节竹叶片碳、氮、磷化学计量比Table 3 The C,N,P stoichiometry for leaf of 1-3-year-old Phyllostachys prominens from experimental stand
3 结论与讨论
叶片是植物进行光合作用的主要器官,其功能性状与植物生物量积累与分配、生长策略和资源的获取与利用密切相关。叶片功能性状由是遗传因素和环境条件共同决定的。同时,也受人工干扰与经营措施的影响[14-15],反映了植物的生态适应对策与资源获取策略[16-18]。本研究发现:覆土控鞭栽培后,高节竹1~3年生立竹叶长、叶宽均有所变化,叶形指数总体升高,虽然与不覆土栽培高节竹林间差异均未达显著水平,但叶片有趋于狭长的趋势;而在适宜林分密度和邻体竞争效应较低情况下,叶片趋于狭长,既可在一定程度上减少自身叶片间的空间竞争和相互干扰,也在一定程度上拓展了生长空间,利于叶片的形态建成和功能发挥。通常,较高的比叶面积意味着具有较高光捕获能力和净光合速率[19-21]。覆土控鞭栽培后,高节竹单叶面积升高,而单叶质量和叶片厚度略有降低,致使比叶面积总体升高,则进一步说明叶片减少了用于构建保卫构造或叶肉细胞密度的投入,将更多物质与能量用于光能捕获与碳同化。这表明覆土控鞭栽培后,高节竹叶片碳同化能力有所增加,叶片厚度与干质量下降,将更多的能量与物质投入到光合结构的构建上,从而形成大而薄的叶片。综上分析表明:虽然覆土控鞭栽培导致林地环境,特别是鞭系分布区的水热条件明显变化,但叶片功能性状与不覆土栽培并无明显差异,这说明高节竹具有很强的生态适应性,能够适应覆土控鞭栽培引起的林地环境扰动与水热条件变化,从而维持较高的竹笋产量,实现可持续经营的目标。
本研究发现,覆土控鞭栽培后,高节竹1~3年生立竹叶片碳质量分数显著升高,氮略有升高,而磷则总体下降,并且C/N略有升高,C/P则显著升高,也即光合碳同化能力与氮、磷利用效率提高[9-10],而N/P亦有小幅升高,即磷素限制作用有所增强,这与磷素利用效率明显提高的研究结果一致。也即,覆土控鞭栽培后,高节竹叶片碳质量分数明显提高,而氮、磷差异不大,但养分利用效率提高,特别是磷素利用效率与限制作用增强。其原因可能是:①覆土控鞭栽培过程中,林地干扰和断鞭刺激致使高节竹产生明显补偿性生长。覆土控鞭栽培前2 a通常会清除覆土层竹鞭(20 cm以内),强烈的土壤扰动和断鞭刺激,致使立竹产生明显的补偿生长,对碳的需求明显增大;而断鞭后,地下鞭根系统结构破坏,生物量下降,导致养分吸收功能降低,这可能是氮磷升幅较小,乃至下降及利用效率升高的重要原因之一。②覆土栽培后,立竹地下鞭根系统分布层下移30 cm以上,鞭根分布区水热条件发生较大改变,对鞭根生长产生一定的非生物胁迫,致使鞭根用于防御的含碳次生代谢物需求量大增,这也是叶片光合固碳能力提升的原因之一。③鞭根分布区下移,高节竹笋出土路程和时间明显延长,生长阻力增大,也势必消耗大量的碳水化合物,从而拉动叶片光合能力的进一步提升。因此,需要进一步研究覆土控鞭栽培后,高节竹叶片光合生理、荧光特性与碳组分的变化规律,探索覆土控鞭栽培高节竹叶片碳含量升高的生理生态学机制。同时,随着覆土控鞭栽培年限的延长,高节竹鞭根系统分布层上移,技术效果会有所弱化。故而,应该进一步研究覆土控鞭栽培年限延长对高节竹生长发育的影响,特别是鞭根系统构型与叶片功能性状的协同变化规律、笋芽萌动与碳积累动态的研究,探索覆土控鞭栽培技术长效稳定维持机制。
[1] PRIOR L D,EAMUS D,BOWMAN D M J S.Leaf attributes in the seasonally dry tropics:a comparison of four habitats in northern Australia [J].Funct Ecol,2003,17(4):504-515.
[2] SUÁREZ N.Leaf construction cost in Avicennia germinans as affected by salinity under field conditions [J].Biol Plant,2005,49(1):111-116.
[3] WRIGHT I J,REICH P B,WESTOBY M.Strategy shifts in leaf physiology,structure and nutrient content between species of high and low rainfall,and high and low nutrient habitats [J].Funct Ecol,2001,15(4):423-434.
[4] 张国平,周伟军.植物生理生态学[M].杭州:浙江大学出版社,2003.
[5] LAMBERS H,POOTER H.Inherent variation in growth rate between higher plants:a search for physiological causes and ecological consequences [J].Adv Ecol Res,1992,23(6):187-261.
[6] 温达志,叶万辉,冯惠玲,等.外来入侵杂草薇甘菊及其伴生种基本光合特性的比较[J].热带亚热带植物学报, 2000, 8(2): 139-146.WEN Dazhi,YE Wanhui,FENG Huiling,et al.Comparison of basic photosynthetic characteristics between exotic invader weed Mikania micrantha and its companion species [J].J Trop Subtrop Bot,2000,8(2):139-146.
[7] GÜSEWELL S,KOERSELMAN W,VERHOEVEN J T A.Biomass N∶P ratios as indicators of nutrient limitation for plant populations in wetlands [J].Ecol Appl,2003,13(2):372-384.
[8] GÜSEWELL S.N∶P ratios in terrestrial plants:variation and functional significance [J].New Phytol,2004,164(2):243-266.
[9] TESSIER J T,RAYNAL D J.Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation [J].J Appl Ecol,2003,40(3):523-534.
[10] KEOERSELMAN W,MEULEMAN A F M.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation [J].J Appl Ecol,1996,33(6):1441-1450.
[11] KERKHOFF A J,FAGAN W F,ELSER J J,et al.Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plant[J].Am Nat,2006,168(4):103-122.
[12] 郭子武,江志标,陈双林,等.覆土栽培对高节竹笋品质的影响[J].广西植物,2015,35(4):515-519.GUO Ziwu,JIANG Zhibiao,CHEN Shuanglin,et al.Influence of soil covered cultivation on shoot quality of Phyllostachys prominens [J].Guihaia,2015,35(4):515-519.
[13] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000.
[14] 王萌,徐冰,张大勇,等.内蒙古克氏针茅草地主要植物叶片功能性状对氮素添加的响应[J].北京师范大学学报(自然科学版), 2016, 52(1): 32-38.WANG Meng,XU Bing,ZHANG Dayong,et al.Leaf functional trait response to nitrogen addition of abundant species in an Inner Mongolia Stipa steppe [J].J Beijing Norm Univ Nat Sci,2016,52(1):32-38.
[15] 刘文亭,卫智军,吕世杰,等.内蒙古荒漠草原短花针茅叶片功能性状对不同草地经营方式的响应[J].生态环境学报, 2016, 25(3): 385-392.LIU Wenting,WEI Zhijun,LÜ Shijie,et al.Response of grassland using modes to leaf trait of Stipa breviflora in desert steppe of Inner Mongolia [J].Ecol Envir Sci,2016,25(3):385-392.
[16] 宝乐,刘艳红.东灵山地区不同森林群落叶功能性状比较[J].生态学报,2009,29(7):3692-3703.BAO Le,LIU Yanhong.Comparison of leaf functional traits in different forest communities in Mt.Dongling of Beijing[J].Acta Ecol Sin,2009,29(7):3692-3703.
[17] CHOWN S L,GASTON K J,ROBINSON D.Macrophysiology:large-scale patterns in physiological traits and their ecological implications [J].Funct Ecol,2004,18(2):159-167.
[18] DIZA S,HODGSON J G,THOMPSON K,et al.The plant traits that drive ecosystems:evidence from three continents[J].J Veg Sci,2004,15(3):295-304.
[19] WRIGHT I J,WESTOBY M,REICH P B.Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span [J].J Ecol,2002,90(3):534-543.
[20] WANG Congyan,ZHOU Jiawei,XIAO Hongguang,et al.Variations in leaf functional traits among plant species grouped by growth and leaf types in Zhenjiang,China [J].J For Res,2017,28(2):241-248.
[21] SCOFFONI C,RAWLS M,McKOWN A,et al.Decline of leaf hydraulic conductance with dehydration:Relationship to leaf size and venation architecture [J].Plant Physiol,2011,156(2):832-843.
Leaf morphology and C,N,and P stoichiometry of Phyllostachys prominens under soil cover with rhizome controlling cultivation
JIANG Zhibiao1,CHEN Shuanglin2,GUO Ziwu2,YANG Qingping2,ZHU Ling1,LI Mingliang1
(1.Agriculture and Forestry Technology Extension Central Station of Tonglu County,Tonglu 311500,Zhejiang,China;2.Research Institute of Subtropical Forestry,Chinese Academy of Forestry,Hangzhou 311400,Zhejiang,China)
To determine the effect of soil cover with rhizome controlling cultivation on growth of Phyllostachys prominens,an excellent shoot and timber used bamboo species ecologically well adapted to its habitat,and to provide guidance for sustainable bamboo management,leaf traits as well as the content of leaf carbon,nitrogen,and phosphorus for 1-3 year-old Ph.prominens of soil cover with rhizome controlling cultivation and normal cultivation were studied.Furthermore,leaf morphology and nutrient stoichiometry were also analyzed.Results showed that 1-3 year-old Ph.prominens leaf of soil cover with rhizome controlling cultivation stand tended to be narrow and long with leaf area and specific leaf area increasing (P>0.05);whereas,leaf mass and thickness decreased slightly (P>0.05).Between soil cover with rhizome controlling cultivation and normal cultivation,there were no differences (P>0.05) for the same above mentioned leaf traits.However,leaf C content greatly increased (P<0.05),N content increased slightly (P>0.05),and P content decreased a little (P>0.05).Also,C/N and N/P increased slightly (P>0.05) with C/P being greatly enhanced (P<0.05).These results indicated that soil cover with rhizome controlling cultivation resulted in slight variation of leaf morphological plasticity for Ph.prominens and showed that with photosynthetic C fixation and nutrient utilization efficiency increasing,high and stable production of superior quality bamboo shoots could be promoted. [Ch,3 tab.21 ref.]
silviculture;Phyllostachys prominens;soil cover with rhizome controlling cultivation;leaf traits;stoichiometry
S795.7
A
2091-0756(2017)06-1155-06
10.11833/j.issn.2095-0756.2017.06.025
2017-01-18;
2017-03-23
国家重点研发计划项目(2016YFD0600903);中央财政林业科技推广示范资金项目(2015TS09);浙江省林业科技推广项目(2012B14)
江志标,高级工程师,从事林业技术研究与推广。E-mail:tlljzx@sohu.com。通信作者:郭子武,副研究员,博士,从事竹林生态与培育等研究。E-mail:hunt-panther@163.com
猜你喜欢
检察风云(2024年11期)2024-06-13 04:00:59
检察风云(2024年7期)2024-05-13 13:05:25
检察风云(2024年9期)2024-05-10 08:54:31
江西农业大学学报(2020年6期)2021-01-08 02:35:50
小天使·一年级语数英综合(2018年4期)2018-06-22 10:33:26
现代园艺(2018年1期)2018-03-15 07:56:21
小天使·一年级语数英综合(2017年11期)2017-12-05 07:18:15
小布老虎(2017年4期)2017-08-10 08:22:40
空中之家(2017年7期)2017-07-20 10:08:06
小学生作文(低年级适用)(2017年3期)2017-07-06 12:06:53
