
硒对农业植物病害防控的研究进展
2017-11-23 05:20明佳佳郑亚伟胡承孝赵小虎
生物技术进展 2017年5期
贾 玮, 明佳佳, 郑亚伟, 贾 芬, 胡承孝, 赵小虎
华中农业大学资源与环境学院, 农业部长江中下游耕地保育重点实验室; 新型肥料湖北省工程实验室, 武汉 430070
硒对农业植物病害防控的研究进展
贾 玮, 明佳佳, 郑亚伟, 贾 芬, 胡承孝, 赵小虎*
华中农业大学资源与环境学院, 农业部长江中下游耕地保育重点实验室; 新型肥料湖北省工程实验室, 武汉 430070
硒是植物的有益元素之一,有助于促进植物生长、增强植物抗逆能力。总结了国内外关于硒提高植物抗逆能力的生物学基础研究,重点综述了硒在农业植物病害防控领域的研究进展,以期为深入开展硒在农业生态安全领域的研究与应用提供理论和实践依据。
硒;农业;植物病害;防控
硒是人和动物必需的微量元素之一,也是对植物生长有益的微量元素[1]。适量的硒有助于促进植物叶绿素的合成、增强新陈代谢、提高光合效率及对矿质元素的转运与积累等[2~4]。此外,硒有助于清除植物体内过量的自由基、延缓细胞衰老,进而增强植物的抗氧化能力,缓解干旱、病害、盐害、低温、重金属毒害等逆境条件对植物的胁迫与损伤[5~7]。
近些年,有关硒及其在农业植物抗逆生物学领域的研究逐渐成为热点[8,9]。其中,重金属污染下硒增强植物抗逆能力的研究较为深入。其他逆境条件下,硒的生物学功能研究尚处于起步阶段[10~13]。尤其是在病害胁迫下,硒对植物抗病能力的提升及其对农业植物病害防控研究较少。鉴于此,本文概述了国内外近些年在硒与农业植物病害防控领域的研究进展,为深入开展硒在农业生态安全领域的研究与应用提供参考。
1 硒提高植物抗逆能力的生物学基础
1.1硒提高植物抗性的生物化学基础
硒促进植物发育及增强对逆境胁迫的耐受能力主要是通过增强植物抗氧化能力、激发植物免疫系统得以实现的。植物在逆境胁迫下会产生大量自由基。植物体内谷胱甘肽过氧化物酶(GSH-Px)的主要功能是加速胞内过氧化物的分解、清除有机氢化物、阻断膜脂质过氧化的连锁反应等。硒可通过提高植物体GSH-Px的活性,进而优化胞内环境,调控胞内系列生物化学反应[14]。同时,硒的存在也可抑制GSH-Px基因转录子的降解,提高DNA修复酶的活力,促进DNA与RNA的合成[15,16]。除GSH-Px外,硒还能够改变胞内超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等活性氧清除相关酶的活性,调控胞内各种抗氧化系统处于动态平衡的状态[17]。
植物体内适量的硒可清除机体内的活性氧自由基。然而,当硒浓度过低或过高时,活性氧自由基会因为得不到及时清除或新的自由基不断产生而增加,可能导致生物效应降低甚至死亡[18]。
1.2硒提高植物抗性的细胞学基础
机体内适量的硒有助于提高植物线粒体呼吸速率,增强线粒体呼吸强度,维持正常的呼吸酶系统活力水平;维持植物线粒体的数量、内嵴的密度、外膜的完整性等[19~22]。硒参与植物线粒体呼吸链辅酶A与辅酶Q的合成,促进丙酮酸脱羧、提升α-酮戊二酸氧化酶系统的活性,在三羧酸循环和呼吸链的电子传递过程中发挥着重要的生物学作用[23,24]。此外,硒还可通过增强GSH-Px酶的活力,进而保护细胞膜结构和功能的完整性。
2 硒在农业植物病害防控领域的研究与应用
2.1硒对植物病原微生物生长的抑制
离体培养条件下,硒对植物病原菌生长的抑制作用已被证实。Razak等[25]采用分离培养的方式研究了硒对镰孢菌、链格孢菌、曲霉菌生长的抑制效果,发现10 mg/kg的亚硒酸钠可使得这3种真菌停止生长。赵献军等[26]研究指出:当培养基中硒的浓度达到20 mg/L时,可显著抑制禾谷镰刀菌、木贼镰刀菌、三线镰刀菌、拟枝孢镰刀菌、串珠镰刀菌、半裸镰刀菌等病原菌的生长。Erika等[27]的研究表明亚硒酸钠能够改变白腐病菌孢子的形态,诱导孢子表面收缩。Wu等[28]研究发现24 mg/L的硒能够显著抑制灰霉病病菌孢子的萌发和菌丝细胞质的流失。
硒对离体条件培养的病原微生物生长的影响主要表现为抑制,并未完全致死。贾芬等[29]以油菜菌核病病原-核盘菌为研究对象,采用PDA培养的形式,研究了不同浓度硒对核盘菌菌丝生长的影响。结果表明:硒显著降低了PDA培养基上核盘菌菌落的直径,抑制了菌丝的生长。随着硒浓度的增加,对核盘菌生长的抑制作用逐渐增大,最大抑制率达80%。将被硒抑制生长的菌丝移至正常培养基上,24 h后菌丝恢复生长,表明外源硒的添加仅抑制了核盘菌的生长,并未使其致死。
2.2硒预处理增强作物抗病能力
目前,硒在农业植物病害防控方面研究和应用较多的是采用硒溶液或含硒药物、生长调理剂等对植物进行预处理[30~33]。主要包括:①以硒溶液的形式在作物叶片进行喷施;②以肥料的形式在作物根部土施。通过硒的预处理,调控植物的抗性生理,诱导植物抗病系统的建立,进而增强作物对病原菌的抗性,降低其发病率。
向果树注射含硒营养液,可显著降低苹果和梨树腐烂病、轮纹病、根腐病等病害的发病率;1.0 mg/L的亚硒酸钠预处理梨树的叶片,3天后接种轮纹病菌,其发病率约为未经硒预处理组发病率的30%[30,31];在经20 mg/L亚硒酸钠预处理过的番茄上接种尖孢镰刀菌,番茄枯萎病的发病率大大降低[32]。
土壤施用适量硒(0.1~0.5 mg/kg土壤)有助于提高油菜对菌核病的抗病性,降低其发病率和病斑的大小[33]。
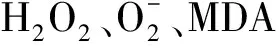
综合已有的研究报道可看出,适量的硒可能从以下几个方面增强植物对病原菌的抵御和对病原生长的抑制能力:①形态结构方面。硒可通过增强植物对矿质元素的吸收、促进细胞结构蛋白的合成、维持细胞膜的稳定性等方面提高寄主细胞的素质,阻断或抵御病原菌的侵入;②生理方面。硒可通过提高植物体内叶绿素的合成,促进线粒体呼吸和叶绿体的电子传递,增强寄主细胞的活力,提高其耐受能力;③生化方面。硒参与并调节GSH-Px、CAT、POD、SOD等酶的活性,直接清除胞内活性氧自由基,维持胞内各种抗氧化系统处于动态平衡的状态,诱导木质素及植保素的合成,提高多酚和醌类物质的含量,并适时诱导寄主的防御反应;④防卫基因表达方面。硒改变了侵染细胞中活性氧的含量、种类及次生代谢产物含量等,在“病原菌-植物”互作体系中诱导植物防卫基因的差异表达。
2.3硒改变作物根际土壤微生物区系特征及病原微生物的丰度
当前,关于硒的施用对土壤微生物区系特征研究较少,尤其是病原微生物群落结构影响的研究报道更是罕见。中国科学院地球化学研究所对湖北恩施渔塘坝富硒碳质泥岩中微生物群落结构特征的调查结果表明:硒显著影响到泥岩中微生物的群落结构,高硒碳质泥岩中具有丰富的微生物多样性[35]。
郑亚伟等采用16S rDNA扩增子测序技术研究了外源硒对油菜根际土壤微生物多样性的影响,并深入分析了硒对土壤中油菜相关病原微生物丰度的影响。物种注释和丰度聚类分析结果表明,油菜根际土壤样品的微生物群落结构由9个属的细菌组成。随土壤硒浓度的增加,其根际土壤的物种丰度降低,分布越不均匀,群落结构越简单。土壤外源施硒1.0 mg/kg处理的油菜根际土壤中酸杆菌门的相对丰度高于对照组,放线菌门的细菌种类随硒施用浓度的增加而降低。0.5 mg/kg硒处理显著提高了油菜根际土壤黑斑病病原菌链格孢属的多样性(未发表数据)。表明土壤施硒影响了油菜病害黑斑病的发生与形成。硒处理显著改变了根际土壤微生物的多样性,也降低了部分土传病害病原的丰度。相关研究结果可为植物硒营养与抗病的根际微生物过程的研究提供参考。
近些年,随着植物病害防控措施的不断发展,通过调控微生物群落结构多样性及其分布特征控制作物病害已成为实现“绿色植保”极有希望的途径之一[34]。因此,深入研究硒-根际微生物区系特征-作物病原的关系,有助于从微生物分子生态的角度寻找绿色、生态的农业植物病害防治措施。
2.4硒预处理提高作物秸秆提取物的抑菌能力
硒预处理可增强植物的抗病能力,这是否与硒处理改变了作物提取物的成分及其生化特征,进而增强了植物的抗病能力有关,目前罕见相关报道。有研究以硒处理植物生长介质,培养并获得富硒大蒜,从大蒜中提取大蒜汁用于抑菌试验,发现经硒预处理获得的大蒜汁对金黄色葡萄球菌、黄曲霉、黑曲霉、啤酒酵母的抑制作用显著增强[17]。贾玮等通过向土壤中施用硒处理,进一步提取并研究了秸秆溶解性有机质(DOM)对核盘菌生长的抑制效果,发现由土壤施用0.5 mg/kg硒预处理获得的油菜秸秆DOM具有显著的抑菌效果,且由此处理获得的油菜秸秆DOM提取率最高。进一步分析DOM的光谱特征,发现油菜秸秆DOM中的腐殖质为富里酸,硒的施用显著降低了油菜秸秆DOM的相对分子量。从荧光光谱图得出,不同浓度硒处理获得的DOM荧光指数随着土壤施硒浓度的增加呈现先升高后降低的趋势,硒浓度为0.1 mg/kg 和0.5 mg/kg处理显著提高了荧光指数。从傅里叶红外光谱图得出,不同浓度硒处理的DOM含有较多的-C=O、-CH2、-OH、-NH2等官能团,随着土壤施硒浓度的增加,这些官能团的含量也随之增加(未发表数据)。这些成分的变化是否为硒预处理增强油菜对核盘菌抗性的内在因素还有待进一步研究。
3 展望
国内外大量研究已证实,硒在植物生长发育、提高农产品产量和品质、促进叶绿素合成、增强植物抗性等方面发挥了重要的作用。硒增强植物抗性、抑制植物病原微生物生长已是不争的事实。但硒在农业植物病害防控过程中的生物学机制的研究仍处于起步阶段。因此,有关硒对农业植物病害防控的研究还需综合利用多种生物学、土壤化学的技术手段,联合微生物生态调控理念,分别对硒调控植物抗病生物学应答机制和对作物微生态环境的优化等方面进行深入探讨。
此外,硒是一种两性元素,其生物正常营养与中毒剂量阈值范围狭窄。在一定浓度范围内有助于增强植物的抗病能力,施用过量反而降低植物抗病性,增大了病害发生的风险。因此,在对作物补硒时要综合考虑土壤的含硒状况、植物的耐硒性及病原菌的生物学特征,充分发挥硒在土壤-植物-病原微生物系统中的积极作用。
[1] Rotruckj T, Popeal, Ganther H E,etal.. Selenium: Biochemical role as a component of gluthathione peroxidase [J]. Science, 1973, 179(5):88-90.
[2] Khaliq A, Aslam F, Matloob A,etal.. Seed priming with selenium: Consequences for emergence, seedling growth, and biochemical attributes of rice [J]. Biol. Trace Elem. Res., 2015, 166: 236-244.
[3] Xue T, Hartikainen H, Piironen V. Antioxidative and growth promoting effect of selenium in senescing lettuce [J]. Plant Soil, 2001, 237: 55-61.
[4] 赵 巍. 硒促进水稻种子萌发的生理机制初探[D]. 河南洛阳: 河南科技大学,硕士学位论文, 2011.
[5] Roqueiro G, Maldonado S, Rios M C,etal.. Fluctuation of oxidative stress indicators inSalixnigraseeds during priming [J]. J. Exp. Bot., 2012, 63: 3631-3642.
[6] 李 彦, 史衍玺, 张英鹏, 等. 盐胁迫条件下硒对小白菜抗氧化活性及膜脂过氧化作用的影响[J]. 植物营养与肥料学报, 2008, 14(4): 749-753.
[7] Hu K L, Zhang L, Wang J T,etal.. Influence of selenium on growth, lipid peroxidation and antioxidative enzyme activity in melon (CucumismeloL.) seedlings under salt stress[J]. Acta Soc. Bot. Polon., 2013, 82: 193-197.
[8] 贾 玮, 吴 隽, 屈婵娟, 等. 硒增强植物抗逆能力及其机理研究进展[J].中国农学通报, 2015, 31(14): 171-176.
[9] 吴之琳, 尹雪斌, 袁林喜, 等. 硒在植物抗逆境胁迫耐受中的作用[J]. 粮食科技与经济,2014, 39(3): 26-31.
[10] Yigit E. The effects of organic selenium on some physiological and biochemical parameters inHordeumvulgareL. andTriticumL. exposed to salt stress [J]. Fresenius Environ. Bull., 2012, 21: 743-747.
[11] Zembala M, Filek M, Walas S,etal.. Effect of selenium on macro- and microelement distribution and physiological parameters of rape and wheat seedlings exposed to cadmium stress [J]. Plant Soil, 2010, 329: 457- 468.
[12] Kumar M, Bijo A, Baghel R S,etal.. Selenium and spermine alleviate cadmium induced toxicity in the red seaweedGracilariaduraby regulating antioxidants and DNA methylation [J]. Plant Physiol. Biochem., 2012, 51: 129-138.
[13] Qing X J, Zhao X H, Hu C X,etal.. Selenium alleviates chromium toxicity by preventing oxidative stress in cabbage (BrassicacampestrisL. ssp.Pekinensis) leaves [J]. Ecotox. Environ. Safe., 2015(114): 179-189.
[14] 李志玉, 郭庆元. 大豆施硒效应和硒素积累特性的初步研究[J]. 中国油料, 1994, 16(1): 41-44.
[15] 麦维军, 王 颖, 梁承邺, 等. 谷胱甘肽在植物抗逆中的作用[J]. 广西植物, 2006, 25(6): 570-575.
[16] 赵中秋, 郑海雷, 张春光, 等. 土壤硒及其与植物硒营养的关系[J]. 生态学杂志, 2003(1): 23-26.
[17] 段咏新, 傅庭治. 硒在大蒜体内的生物富集及其抗氧化作用[J]. 园艺学报, 1997, 4:343-347.
[18] 徐辉碧, 张罗平, 范华汉, 等. 硒化合物与脂质过氧自由基作用的研究[J]. 化学学报, 1989, 47: 901-906.
[19] 薛泰麟, 候少范, 谭见安, 等. 硒在高等植物体内的抗氧化作用—Ⅱ. 非酶机制的探讨[J]. 科学通报, 1993, 38(4): 356-358.
[20] 杜 倩, 王昌全, 李 冰, 等. 硒、锌及其交互作用对夏茶多酚氧化酶(PPO)活性的影响[J]. 植物营养与肥料学报, 2009, 15(4): 930-935.
[21] Djanaguiraman M, Prasad P V V, Seppanen M,etal.. Selenium protects sorghum leaves from oxidative damage under high temperature stress by enhancing antioxidant defense system [J]. Plant Physiol. Biochem., 2010, 48, 999-1007.
[22] Dziubinska H, Filek M, Krol E,etal.. Cadmium and selenium modulate slow vacuolar channels in rape (Brassicanapus) vacuoles [J]. J. Plant Physiol., 2010, 167, 1566-1570.
[23] Filek M, Koscielniak J, Labanowska M,etal.. Selenium-induced protection of photosynthesis activity in rape (Brassicanapus) seedlings subjected to cadmium stress fluorescence and EPR measurements [J]. Photosyn. Res., 2010, 105(1): 27-37.
[24] Zheng W, Tan X L, Zhang Z Z,etal.. Defense toSclerotiniasclerotirumin oilseed rape is associated with the sequential activations of salicylic acid signaling and jasmonic acid signaling [J]. Plant Sci., 2012, 184: 75-82.
[25] Razak A, El-Tantawy H, El-Sheikh H,etal.. Influence of selenium on the efficiency of fungicide action against certain fungi [J]. Biol. Trace Elem. Res., 1991, 28(1):47-56.
[26] 赵献军,汪昭贤,谢毓芬. 硒与霉菌相互影响的研究[J]. 西北农业学报, 2005, 14(5): 49-52.
[27] Erika J E, Graciela G G, Pascal E,etal.. Effects of selenium oxyanions on the white-rot fungusPhanerochaetechrysosporium[J]. Appl. Microbiol. Biot., 2015, 99: 2405-2418.
[28] Wu Z, Yin X, Bauelos G S,etal.. Effect of Selenium on control of postharvest gray mold of tomato fruit and the possible mechanisms involved [J]. Front. Microbiol., 2016, 6(732): 1441.
[29] 贾 芬, 朱 婷, 赵小虎, 等. 硒对油菜菌核病菌的抑制作用[J]. 中国农学通报, 2015, 31(25): 176-181.
[30] 刘延隆. 富硒植物营养注射液的应用推广初探[D]. 陕西杨凌: 西北农林科技大学, 硕士学位论文, 2007.
[31] 吴玉星, 周宗山, 王孝娣, 等. 生长期喷施氨基酸硒叶面肥对苹果树腐烂病的控制作用研究[J]. 中国果树, 2012, 5: 51-53.
[32] Companioni B, Medrano J, Torres J,etal.. Protective action of sodium selenite against fusarium wilt in tomato: Total protein contents, levels of phenolic compounds and changes in antioxidant potential [C]. Proceedings of the II International Symposium on Soilless Culture and Hydroponics, 2011.
[33] 明佳佳. 硒增强油菜对菌核病抗性及其生理机制[D]. 武汉: 华中农业大学, 硕士学位论文, 2016.
[34] 明佳佳, 胡承孝, 赵小虎, 等. 硒对油菜各部位矿质元素含量及其迁移特征的影响[J]. 浙江农业学报, 2016, 28(9): 1564-1571.
[35] 雷 磊, 朱建明, 王风平, 等. 湖北恩施鱼塘坝富硒碳质泥岩中的微生物多样性初探[J]. 地球与环境, 2011, 39(4): 517-522.
胡承孝教授团队介绍
胡承孝研究团队将“植物微量元素营养”与“生态安全”结合,致力于以元素硒增强植物抗逆能力、维护农业生态安全的机理研究。目前,在本领域已发表代表性论文20多篇,申请专利4项,获得广东省科技进步三等奖、湖北省优秀博士论文等奖励。主持硒相关科研项目包括:国家自然科学基金2项、国家科技支撑计划1项、湖北省自然科学基金1项、其他省部级项目3项。
ProgressonPreventionandControlofSeleniumtoAgriculturalPlantDiseases
JIA Wei, MING Jiajia, ZHENG Yawei, JIA Fen, HU Chengxiao, ZHAO Xiaohu*
KeyLaboratoryofArableLandConservationinMiddleandLowerReachesofYangtzeRiver,MinistryofAgriculture;HubeiProvincialEngineeringLaboratoryforNew-typeFertilizer,CollegeofResourcesandEnvironment,HuazhongAgriculturalUniversity,Wuhan430070,China
Selenium (Se) is one of the beneficial elements for plant, which could improve the growth of plants and enhance the ability of plants to resist the stress of adversity. In the present review, the research and application literatures refer to biological basis of Se improving plant resistance to stress were summarized. This review focused on the new findings of the prevention and control of Se to agricultural plant diseases. The purpose was to provide theoretical basis for the study and application of Se in the field of agricultural ecological safety.
selenium; agriculture; plant diseases; prevention and control
2017-07-10;接受日期2017-07-31
国家自然科学基金项目(41571321;31201501);国家科技支撑计划项目(2014BAD14B02);湖北省自然科学基金项目(2015CFB582)资助。
贾 玮,博士研究生,主要从事硒与生态安全研究。E-mail: jiaweihzau@qq.com。*通信作者:赵小虎,副教授,硕士生导师,主要从事微量元素与农业生态安全研究。E-mail: 50654069@qq.com
10.19586/j.2095-2341.2017.0078

猜你喜欢
今日农业(2021年21期)2021-11-26
今日农业(2021年14期)2021-10-14
湖北农机化(2020年4期)2020-07-24
娃娃乐园·综合智能(2018年3期)2018-03-22
数学小灵通(1-2年级)(2017年10期)2017-11-08
制导与引信(2017年3期)2017-11-02
课程教育研究·学法教法研究(2017年12期)2017-07-12
工业设计(2016年11期)2016-04-16
现代农业(2016年6期)2016-02-28
天津农业科学(2015年12期)2015-12-03
