
茶树硒吸收及耐性机理研究进展
2017-11-23 05:20金孝芳龚自明马林龙刘艳丽
生物技术进展 2017年5期
曹 丹, 金孝芳, 龚自明, 马林龙, 刘艳丽, 郑 琳
湖北省农业科学院果树茶叶研究所, 武汉 430064
茶树硒吸收及耐性机理研究进展
曹 丹, 金孝芳*, 龚自明*, 马林龙, 刘艳丽, 郑 琳
湖北省农业科学院果树茶叶研究所, 武汉 430064
茶树对硒具有较强的富集能力,可为人体提供安全有效的有机硒。主要阐述了茶树不同部位、不同品种硒含量的差异,综述了不同形态硒的吸收以及硒耐受机制等方面的研究进展,并对后续研究进行了展望,以期为富硒茶新品种的选育及开发提供理论依据。
硒;茶树;吸收;耐性
硒是人和动物必需的微量元素,但其有益和毒性剂量阈值很小[1],适量的硒可以增强人和动物的免疫力,预防多种疾病的发生,缺硒会引起多种疾病,如克山病、大骨节病等,过量硒摄入则会引起中毒[2]。植物可将无机硒转化为安全有效的有机硒,是人体摄取硒的主要来源。茶树(CamelliasinensisL. O. Ktze.)是一种集硒植物,有机硒是茶叶中硒的主要形态[3]。故饮茶可作为人体摄入硒的方式之一。研究茶树对硒的吸收机理,对调控茶叶硒含量具有重要的指导意义。本文综述了茶树不同部位、不同品种硒含量的差异,并探讨了不同形态硒的吸收以及硒耐受机制等方面的研究进展,以期为茶树的育种及富硒茶产品的开发提供参考。
1 茶树硒含量的差异性
1.1茶树不同部位硒含量的差异
硒在茶树体内的分布,依据其部位不同而表现出极大的差异。通过土壤施硒发现,茶树不同部位总硒含量的增加幅度表现为:根部最高,2年生枝次之,叶部最低,且新叶高于老叶[4],说明硒可能靠浓度差由根系吸收,经茎向地上部迁移,而老叶中的硒有向新叶中转运的趋势[5]。另有研究发现,不同叶位间的硒含量也存在显著差异,呈现出老叶>4叶>3叶>2叶>1叶的规律,原因可能是随着茶树叶片的成熟老化,体内的硒向结合态转变,逐渐形成不易流动的大分子硒蛋白,最终导致老叶的含量高于嫩叶[6~8]。
对茶树叶片的亚细胞研究发现,春季茶叶中的硒初期主要累积在膜和细胞器(F2),随着时间的延长逐渐向可溶性部分(F3)和细胞壁(F1)转移,最终呈现F1>F2>F3的分布趋势,而夏秋两季的硒分布规律均表现为F1>F2>F3的特点。另外,在单硒和硒锌交互处理时,硒大多集中在膜和细胞器部分,而单锌处理时则主要分布在细胞壁中[9]。
1.2不同品种茶树硒含量的差异
我国茶树种质资源丰富,不同品种对硒的累积量及其富集能力差异显著。王雅玲等[10]依据茶树叶片和根部硒含量比值不同,将供试的11个品种分为富硒、中等富硒和非富硒品种。Zhao等[11]通过水培的方式对14个品种进行硒处理,发现乌牛早和龙井43的硒含量增加最多,说明其吸收转运硒的能力较强,而梅占、福云6号和中茶108等3个品种的硒含量变化不大;通过SSR分析发现该14个品种遗传多样性丰富,可为富硒品种的培育提供材料基础。因此,充分利用茶树种质资源进行富硒品种的筛选,结合现代育种技术手段和方法,在兼顾茶叶品质的基础上,培育优质富硒茶树新品种是完全可能的。
2 茶树对硒的吸收
植物吸收硒的主要形式为硒酸盐和亚硒酸盐。硒酸盐是碱性和氧化环境中的主要存在形式,亚硒酸盐是酸性和中性土壤中的主要存在形式,在还原性很强的条件下硒化物则占主导[12]。茶园土壤大多显酸性,因而硒的主要存在形式是亚硒酸盐[5]。植物吸收硒酸盐时,根系吸收的硒能快速向地上部转移,如果浓度控制不好较易产生毒害,而亚硒酸盐被植物吸收后,很容易转化为硒代氨基酸及其氧化物等,这些物质主要累积在根部,但是更易生产出地上部富集机硒的植物[13,14]。
2.1茶树根系对硒的吸收
硒与硫属于同一主族元素,物理性质和化学性质相似[15]。多数研究表明,植物通过硫转运子对硒酸盐进行吸收和转运[16],黄进等[17]研究发现低浓度硫酸盐促进茶树对硒酸盐的吸收,而高浓度硫酸盐对其具有抑制作用。目前已从茶树中克隆出CsSUL3.5的cDNA序列,生物信息学分析发现该基因属于硫酸盐转运蛋白家族,硒酸盐处理能显著诱导其表达[18]。另外,通过筛选拟南芥耐硒酸盐突变体,鉴定出了高亲和硫转运蛋白Sultrl;2,研究表明Sultrl;2除转运硫酸盐外,还可转运硒酸盐;而超富硒植物体内积累的硒含量远高于非积累型植物,可能由于它们的转运蛋白具有不同的选择性和转运能力[19]。
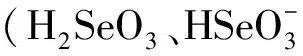
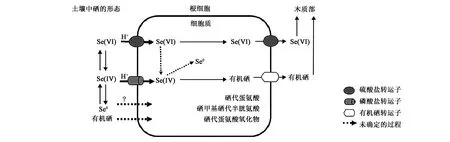
图1 植物对不同形态硒吸收转运示意图[20]Fig.1 Illustration of selenium uptake and translocation in plants[20].
通过对茶树不同部位韧皮部与木质部的硒含量分析发现,根系吸收的硒在两个部位间可能存在横向迁移,主要利用韧皮部向上运输[17]。另有研究认为,植物根系吸收的硒酸盐经木质部向地上部转运,而有机硒是通过韧皮部向上转移的[1]。造成这种差异的原因可能是植物根系吸收了不同形态的硒。
2.2茶树叶片对硒的吸收
叶片能够快速吸收营养物质并转运到植株其他部位,故叶面喷施是一种调控植株营养状况的快速有效方法[26]。有研究发现,叶片喷施Na2SeO3对夏茶品质提高最显著,主要表现为降低酚氨比,同时提高可溶性糖的含量[27]。同时,在春秋季茶叶采摘前喷施Na2SeO3也能显著提高硒含量,且与喷施浓度存在极显著相关性,但在秋茶生产季节喷硒时,虽然叶片中的硒含量能明显增加,但是茶多酚含量增加、氨基酸含量降低,导致茶汤滋味苦涩、香气下降[28]。硒在茶树体内的主要富集形态是硒蛋白,参与合成的硒代氨基酸有3种,分别是硒甲基半胱氨酸(SeMeCys)、硒代半胱氨酸(SeCys)和硒代蛋氨酸(SeMet),其中SeMeCys是主要形式。当茶树叶面喷施Na2SeO3时,随着浓度的逐渐增加,茶叶中的SeMeCys呈现出先升后降的趋势,这与总的硒代氨基酸的含量变化相同,而浓度过高则会导致叶片上出现褐色的斑点甚至焦化[29]。
叶面喷硒虽具有见效快、成本低等优点,但要兼顾富硒和品质优的平衡,既要严格把握好施肥时间,避免未被叶面吸收而残留在嫩梢上的硒进入加工环节,又要合理控制施用量,防止过量的硒对植株产生毒害。有研究发现人工施用亚硒酸盐生产的富硒茶具有与天然富硒茶具有相近的硒形态和抗氧化功能[30],而另有研究认为,人工施硒不仅对土壤中元素间的化学平衡有一定的影响,还会改变植物的正常生理周期,表现为缩短营养品质形成的关键时期,从而导致品质下降,故若非选择能够适应早萌发或早成熟的品种,是否需要采用人工手段富硒还需权衡[31]。
3 茶树硒吸收及耐受机制
3.1代谢关键酶
硒经植物吸收后发生了一系列的同化代谢反应,存在多个关键酶。ATP硫化酶(ATPS)是硫酸盐代谢途径中的第一个酶,也是限速酶[32],它可以促进硒酸盐转化为亚硒酸盐,进而形成有机硒;硒代半胱氨酸甲基转移酶(SMT)可将硒代半胱氨酸(蛋白质氨基酸)催化形成硒甲基硒代半胱氨酸(非蛋白质氨基酸),降低了硒蛋白的毒性,以此来增加植物对硒的耐受度[33]。目前已从茶树中分离出 ATP硫酸化酶和硒半胱氨酸甲基转移酶(CsSMT)的全长序列,并对其进行了结构分析及功能验证[33,34]。另外,硒的毒害作用主要源于在蛋白质位点硒半胱氨酸取代半胱氨酸,而硒代半胱氨酸裂解酶(SL)可以将硒代半胱氨酸裂解为硒和丙氨酸,减少硒进入蛋白的机会。Garifullina等[35]将该酶基因转入印度芥菜中进行表达发现,在提高转基因植物硒吸收能力的同时却降低了硒的耐受性,可能是SL介导产生了大量元素态硒,进一步参与了植物硫代谢途径;类似结果也表现在拟南芥过表达一种类似叶绿体Nifs蛋白(AtCpNifS),AtCpNifS转基因植物同样表现出硒积累能力增强的特点,同时该基因在植物修复中也有一定的应用价值[36]。
3.2抗氧化系统
茶树在正常生长代谢过程中,会在其细胞质膜上进行电子传递和有氧呼吸,产生大量的活性氧自由基,一旦过量将会造成氧化应激和代谢紊乱[37]。谷胱甘肽过氧化物酶(GSH-Px)在植物活性氧清除过程中起重要作用[38],它的主要功能是催化还原型谷胱甘肽(GSH)生成氧化型谷胱甘肽(GSSG),同时使H2O2被还原生成H2O,降低活性氧的含量,进而阻断其对细胞核组织的过氧化链式反应[39]。硒是GSH-Px催化活性中心的重要组成部分。研究发现,茶树在高硒环境下,其体内的GSH-Px仍能保持较高的活性,推测硒促进GSH-Px的合成,并维持其催化活性以抵御细胞膜质过氧化,使茶树表现出了较强的耐性[17]。另外,GSH也与植物硒吸收或耐受性有关。张联合等[40]研究发现,外源GSH能显著促进水稻根系对亚硒酸盐的吸收,吸收量是对照的2倍多,这是由于GSH可以通过参与亚硒酸盐的代谢进而促进硒的吸收,而L-cys利用其在根内的代谢产物GSH也能间接促进亚硒酸盐的吸收。
抗坏血酸过氧化物酶(APX)也是植物调节H2O2非常重要的一种酶,该酶属于Ⅰ型血红素过氧化物酶,对抗坏血酸具有高度的特异性[41]。对拟南芥等植物的研究发现,APX1基因参与调控硒耐受的分子机制,可能的途径有两条,一是外源硒参与GSH-Px的合成,加快体内活性氧的清除;二是外源硒可能通过活性氧参与细胞信号转导,增加体内GSH的含量,螯合过量的硒。这两种途径也可能共同调节植物的硒耐受和累积能力[42]。
3.3植物激素
植物激素对植物的生长发育和新陈代谢等具有重要的作用,同时也与营养元素的吸收、运输和转运密切相关[43]。研究发现,硒处理可以诱导超富集植物Stanleyapinnata体内的茉莉酸(JA)、茉莉酸甲酯(MeJA)、乙烯(ETH)和水杨酸(SA)等植物激素生物合成相关基因的上调表达,推测这些激素促进了植物体内硫的吸收同化,间接调控了硒的吸收与代谢,从而对植物的富硒及耐硒性发挥积极的作用[44]。另有研究表明,JA和ETH也与拟南芥的硒耐受性相关[45,46],而细胞分裂素(CTK)和一氧化氮(NO)互作与其耐受性呈负相关,含量过多会降低植株的硒敏感性[47]。 周维等[48]试验发现,春季喷施MeJA可有效提高茶叶对Na2SeO4的吸收,而对Na2SeO3作用不显著,其机理有待于深入研究。
4 展望
目前,人们对拟南芥、水稻、紫云英等植物硒吸收的研究已经取得了重要的突破,然而对茶树硒吸收的研究尚属初级阶段,还有诸多方面有待深入探讨,建议从以下几方面进行:①加大硒在茶树亚细胞中的分布、存在形式以及结合位点等的研究力度;②茶树吸收硒酸盐的研究已经有了一定基础,但对亚硒酸盐的吸收包括吸收方式、影响因素、吸收相关基因的挖掘和功能验证等仍需探究;③抗氧化系统对茶树耐硒性发挥了重要的作用,但是抗氧化基因的挖掘也有待于深入研究;④植物激素对元素的吸收、运输和分布起着重要作用。茶树在对硒吸收的过程中,植物激素含量和合成相关基因的转录水平发生了何种变化,是单一激素影响还是多种激素交互作用,这些都亟待探讨。
[1] Zhu Y G, Pilonsmits E A H, Zhao F J,etal.. Selenium in higher plants: Understanding mechanisms for biofortification and phytoremediation[J]. Trends Plant Sci., 2009, 14(8):436-442.
[2] Winkel L H, Johnson C A, Lenz M,etal.. Environmental selenium research: from microscopic processes to global understanding.[J]. Environ. Sci. Technol.,2012,46(2):571-579.
[3] 龚晓钟, 欧阳政, 蔡端仁. 富硒茶叶和富硒大蒜中硒的有机形态[J]. 天然产物研究与开发, 1996(1):59-62.
[4] 许春霞, 李向民, 肖永绥. 土施硒肥与茶叶含硒量和产量的关系[J]. 西北农业学报, 1996, 5(1):71-75.
[5] 朱 林, 房婉萍, 叶爱华,等. 植物硒代谢与茶树硒营养研究进展[A]. 见:安徽食品安全博士科技论坛论文集[C]. 安徽食品安全博士科技论坛, 2005.
[6] 杜琪珍, 方兴汉. 茶树累积硒的动态分布和主要形态[J]. 中国茶叶, 1991(3):8-9.
[7] 顾 谦, 赵慧丽, 童梅英,等. 茶叶中总硒含量及其影响因素的研究[J]. 生物数学学报, 1994(S1):108-113.
[8] 周 超. 土壤因子对茶树硒吸收特性的影响[D]. 重庆:西南大学,硕士学位论文, 2015.
[9] 杜 倩. 硒锌交互对茶叶亚细胞中硒锌分布及多酚氧化酶(PPO)活性的影响[D]. 四川雅安:四川农业大学, 硕士学位论文, 2009.
[10] 王雅玲. 皖南茶区土壤中铜、铅、硒含量及其茶树吸收、分配与品种差异[D]. 南京: 南京农业大学, 硕士学位论文, 2005.
[11] Zhao H, Huang J, Li Y,etal.. Natural variation of selenium concentration in diverse tea plant (Camelliasinensis) accessions at seedling stage[J]. Sci. Hortic., 2016, 198:163-169.
[12] Elrashidi M A, Adriano D C, Workman S M,etal.. Chemical equilibria of selenium in soils: A theoretical development1[J]. Soil Sci., 1987, 144(2):274-280.
[13] Mehdawi A F E I, Pilon-Smits E A H. Ecological aspects of plants selenium hyperaccumulation[J]. Plant Biol.,2012,14:1-10.
[14] Longchamp M, Angeli N, Castrec-Rouelle M. Selenium uptake inZeamayssupplied with selenate or selenite under hydroponic conditions[J]. Plant Soil, 2013, 362:107-117.
[15] Wessjohann L A, Schneider A, Abbas M,etal.. Selenium in chemistry and biochemistry incomparison to sulfur[J]. Biol.Chem., 2007,388(10):997-1006.
[16] Shibagaki N, Rose A, Mcdermott J P,etal.. Selenate-resistant mutants ofArabidopsisthalianaidentify Sultr1;2, a sulfate transporter required for efficient transport of sulfate into roots[J]. Plant J., 2002, 29(4):475-486.
[17] 黄 进. 硒对茶树抗氧化系统的影响及其在品种间富集特性研究[D]. 武汉:华中农业大学, 硕士学位论文, 2014.
[18] 胡玉荣, 岳 川, 周 超,等. 茶树硫酸盐转运蛋白基因CsSUL3.5的克隆与表达分析[J]. 园艺学报, 2015, 42(11):2306-2314.
[19] Galeas M L, Zhang L H, Freeman J L,,etal.. Seasonal fluctuations of selenium and sulfur accumulation in selenium hyper-accumulators and related nonaccumulators[J]. New Phytol., 2007, 173: 517-525.
[20] 王晓芳, 陈思杨, 罗 章,等. 植物对硒的吸收转运和形态转化机制[J]. 农业资源与环境学报, 2014(6):539-544.
[21] 张联合,李友军,苗艳芳,等. pH对水稻离体根系吸收亚硒酸盐生理机制的影响[J]. 土壤学报,2010,47(3):523-528.
[22] Zhang L H, Yu F Y, Shi W M,etal.. Physiological characteristics of selenite uptake by maize roots in response to different pH levels[J]. J. Plant Nutr. Soil Sci.,2010,173(3):417-422.
[23] Zhang L, Shi W, Wang X. Difference in selenite absorption between high- and low-selenium rice cultivars and its mechanism[J]. Plant Soil, 2006, 282(1):183-193.
[24] Zhao Z X, Mitani N, Yamaji N,etal.. Involvement of silicon influx transporter OsNIP2;1 in selenite uptake in rice[J]. Plant Physiol.,2010,153:1871-1877.
[25] 叶 飞, 龚自明, 高士伟,等. 湖北恩施茶园土壤及茶叶硒元素调查研究[J]. 四川农业大学学报, 2015(3):275-278.
[26] Gupta U C, Winter K A, Mcrae K B. Selenium enrichment of crops through follar applications [J]. Can. J. Soil Sci., 1988, 68(3):519-526.
[27] 李 静, 夏建国, 巩发永,等. 外源硒肥对茶叶硒含量及化学品质的影响研究[J]. 水土保持学报,2005,19(4):104-106.
[28] 王 磊, 黄婷婷, 杨 春,等. 叶面喷硒对台茶12号所制红绿茶含硒量及品质的影响[J]. 西南农业学报, 2016, 29(11):2578-2582.
[29] 秦冰. 茶树硒代氨基酸的赋存形态及外源硒肥对其形态和含量影响的研究[D]. 合肥:安徽农业大学, 硕士学位论文, 2013.
[30] 胡秋辉, 潘根兴, 安辛欣,等. 天然和人工富硒茶叶的抗氧化功能比较[J]. 营养学报, 2001, 23(3):242-245.
[31] 罗 杰, 温汉辉, 吴丽霞,等. 自然富硒与人工施硒肥的比较[J]. 中国农学通报, 2011, 27(33):90-97.
[32] Schiavon M, Pilon M, Malagoli M,etal.. Exploring the importance of sulfate transporters and ATP sulphurylases for selenium hyperaccumulation-a comparison ofStanleyapinnataandBrassicajuncea(Brassicaceae).[J]. Front Plant Sci., 2015, 6(6):2.
[33] Zhu L, Jiang C J, Deng W W,etal.. Cloning and expression of selenocysteine methyltransferase cDNA fromCamelliasinensis[J]. Acta Physiol. Plant, 2008, 30(2):167-174.
[34] 王雅楠. 茶树ATP硫化酶和硒代半胱氨酸甲基转移酶的基因克隆和启动子结构分析[D]. 合肥:安徽农业大学, 硕士学位论文, 2013.
[35] Garifullina G F, Owen J D, Lindblom S D,etal.. Expression of a mouse selenocysteine lyase inBrassicajuncea, chloroplasts affects selenium tolerance and accumulation[J]. Physiol. Plant, 2003,118(4):538-544.
[36] Douglas V H, Gulnara F G, Ashley R A,etal.. Over-expression of AtCpNifS enhances selenium tolerance and accumulation inArabidopsis[J]. Plant Physiol., 2005, 139: 1518-1528.
[37] 陈瑾歆, 唐聪明. 氧自由基的研究进展[J]. 海南医学院学报, 2004, 10(3):206-208.
[38] Upadhyaya C P, Venkatesh J, Gururani M A,etal.. Transgenic potato overproducing L-ascorbic acid resisted an increase in methylglyoxal under salinity stress via maintaining higher reduced glutathione level and glyoxalase enzyme activity.[J]. Biotechnol. Lett., 2011, 33(11):2297-2307.
[39] 林源秀, 顾欣昕, 汤浩茹. 植物谷胱甘肽还原酶的生物学特性及功能[J]. 中国生物化学与分子生物学报, 2013, 29(6):534-542.
[40] Zhang L, Yu F, Shi W,etal.. L-cysteine stimulates selenite uptake through GSH involving in selenite reduction in rice roots[J]. J. Plant Nutr., 2015, 38(5):768-779.
[41] 李泽琴, 李静晓, 张根发. 植物抗坏血酸过氧化物酶的表达调控以及对非生物胁迫的耐受作用[J]. 遗传, 2013, 35(1):45-54.
[42] 陈子平. 一个调控拟南芥硒耐受及硒积累的基因功能研究[D]. 合肥:合肥工业大学, 硕士学位论文, 2014.
[43] 汪 洪, 金继运, 周 卫,等. 植物营养元素与激素之间相互关系的研究[A]. 见: 全国青年植物营养科学工作者学术讨论会论文集[C]. 全国青年植物营养科学工作者学术讨论会,2000.
[44] Freeman J L, Tamaoki M, Stushnoff C,etal.. Molecular mechanisms of selenium tolerance and hyperaccumulation inStanleyapinnata[J]. Plant Physiol., 2010, 153(4):1630-1652.
[45] Inoue E. Transcriptome analyses give insights into selenium-stress responses and selenium tolerance mechanisms inArabidopsis[J]. Physiol. Plant., 2008, 132(2):236-253.
[46] Tamaoki M, Freeman J L, Pilon-Smits E A. Cooperative ethylene and jasmonic acid signaling regulates selenite resistance inArabidopsis[J]. Plant Physiol., 2008, 146(3):1219-1230.
[47] Lehotai N, Feigl G, Koós,etal.. Nitric oxide-cytokinin interplay influences selenite sensitivity inArabidopsis[J]. Plant Cell Rep., 2016, 35(10):1-15.
[48] 周 维, 魏成熙, 韦 伟. 茉莉酸甲酯对富硒春茶硒含量及品质的影响[J]. 山地农业生物学报, 2012, 31(6):514-518.
龚自明研究员团队介绍
本研究团队多年来一直从事茶树种质资源收集保存与鉴定利用、茶树新品种选育等方面的研究工作。在硒学研究方面,目前主要开展茶树响应硒的分子机制以及富硒品种选育等方面的研究。迄今为止,先后选育出无性系优良茶树品种6个,其中国家级品种2个(鄂茶1号、鄂茶5号),并收集保存国内外茶树种质资源1 200余份;获得省(部)级科技成果奖励10余项,国家专利20余项,制订省级地方标准10余项。
ProgressonSeleniumAbsorptionandToleranceMechanismofTeaPlant
CAO Dan, JIN Xiaofang*, GONG Ziming*, MA Linlong, LIU Yanli, ZHENG Lin
FruitandTeaResearchInstitute,HubeiAcademyofAgriculturalSciences,Wuhan430064,China
Camelliasinensiswith strong enrichment ability of selenium (Se), can provide effective organic Se for human body. The latest advances were summarized in this review, which mainly focused on the Se content in different types and parts of tea plant, absorption of different forms and the mechanism of tolerance. Moreover, a prospect for future research was made, which was expected to provide a theoretical basis for breeding and development of Se-rich tea.
selenium; tea plant; absorption; tolerance
2017-06-21;接受日期2017-07-13
湖北省农业科技创新中心项目(2016-620-000-001-032)资助。
曹 丹,助理研究员,主要从事茶树资源与育种研究。E-mail:skyiswide@163.com。*通信作者:金孝芳,助理研究员,博士,主要从事茶树资源与育种研究。E-mail:jxf1130@126.com;龚自明,研究员,主要从事茶叶加工及综合利用技术等研究。E-mail:ziminggong@163.com
10.19586/j.2095-2341.2017.0065
猜你喜欢
国际放射医学核医学杂志(2020年2期)2020-05-30
乡村地理(2018年2期)2018-09-19
现代检验医学杂志(2016年5期)2016-08-20
湖南农业(2016年3期)2016-06-05
音乐天地(音乐创作版)(2016年11期)2016-02-05
东北电力大学学报(2015年4期)2015-11-13
中国当代医药(2015年22期)2015-03-01
应用化工(2014年8期)2014-08-08
安徽医专学报(2014年6期)2014-03-20
无机化学学报(2014年3期)2014-02-28
