
青藏高原高寒草甸群落特征对氮沉降和增水的响应
2017-11-17 00:53许庆民郭小伟曹莹芳杜岩功青海省环境监测中心站青海西宁80008威海市农业局山东威海644中国科学院西北高原生物研究所青海西宁8000
草原与草坪 2017年5期
许庆民,周 赓,郭小伟,曹莹芳,杜岩功(.青海省环境监测中心站,青海 西宁 80008; .威海市农业局,山东 威海 644;.中国科学院西北高原生物研究所,青海 西宁 8000)
青藏高原高寒草甸群落特征对氮沉降和增水的响应
许庆民1,周 赓2,郭小伟3,曹莹芳3,杜岩功3
(1.青海省环境监测中心站,青海 西宁 810008; 2.威海市农业局,山东 威海 264411;3.中国科学院西北高原生物研究所,青海 西宁 810001)
试验选择高寒草甸为研究对象,通过连续两年定位观测,采用模拟氮沉降与增水及其交互作用对高寒草地群落特征的影响,为高寒草地生产力功能提升提供科学依据。结果表明:模拟氮沉降和增水及其交互处理,当年均能够显著增加高寒草甸地上总生物量、禾本科植物、豆科植物生物量,但显著降低莎草科植物生物量;而在第2年,增水处理并未明显增加地上生物量,氮沉降与增水交互作用可以稳定地提高高寒草地生产力,各处理均能够明显提高禾本科和莎草科生物量、降低杂类草生物量。增水当年能够显著提高高寒草甸物种丰富度和生物多样性指数,但氮沉降和增水交互降低物种多样性指数;而在第2年,各处理物种数目和多样性指数均降低,且氮沉降与增水的交互作用影响效应达到极显著性检验水平。综合分析,氮沉降和增水及其交互作用在提升草地生产力、优良牧草生物量的同时,也可能造成高寒草甸物种丰富度和多样性指数降低,在草地的适应性管理方面需要引起足够的关注。
氮沉降;增水;高寒草甸;地上生物量;物种多样性
氮元素是所有生命体的基本营养物质之一,是蛋白质的关键组成成分,氮也是维持生物体结构组成和运转生物化学循环过程的基础,是草地植物生长的关键限制因素[1]。近年来,人类活动导致大气氮沉降量明显增加,我国境内大气干湿沉降总量平均值为12.9 kg/(hm2·a),部分地区高达63.5 kg/(hm2·a),已成为继欧洲、北美之后的第三大氮素沉降区[2-3],氮输入可能缓解草地生态系统氮素限制,氮素有效性增加能够提高地上生物量和凋落物量,减少光合产物向地下碳分配数量,提高草地生态系统初级生产力,但也有研究发现氮有效性增加能提高资源消耗型或酸适应物种多度[4-5]。
受全球CO2浓度和平均气温不断增高的影响,全球水分时空格局正在发生巨大改变,近50年来青藏高原地区冰川退缩加剧,降水呈现逐渐增加趋势,其中,冬季降水每10年约增加14%[6]。水热条件及其组合是决定植物群落分布的主要因素,降水通过改变土壤肥力、地表温度和通气状况,影响草地群落物种分布、物种多样性与初级生产力关系密切[7]。
以往研究较多关注高寒草甸不同物种对短期的生长季施氮肥或增雨的单一处理响应差异、氮素吸收能力、根系氮素含量和根冠比等功能性状的种间差异[8-9],结果表明氮沉降显著提高草地地上生物量,但随着施氮量增加,地上生产力逐渐降低,物种丰富度和多样性下降[10-12]。莎草科植物对施肥处理响应不敏感,说明其对环境的波动适应性较强,而杂类草和禾本科牧草敏感性高于莎草科牧草[9]。也有研究发现模拟增水处理与对照相比,地上生物量变化不显著[9]。目前有关施肥及增水处理,对高寒草甸生物量、多样性特征的短期及较长时间的影响研究较少。
依据连续两年观测结果,分析青藏高原高寒草甸群落特征对较长时期氮沉降和增水及其交互作用的响应规律,为高寒草地生产力提升与可持续利用提供科学指导,同时也是在全球变化背景下,丰富高寒草地氮素循环研究的理论基础。
1 材料和方法
1.1研究地区概况
试验设置在中国科学院海北高寒草甸生态系统定位站(海北站),地处祁连山北支冷龙岭东段南麓的大通河谷,N 37°29′~37°45′,E 101°12′~101°23′,海拔3 280 m。属典型高原大陆性气候,年均气温-1.7℃,最冷月1月平均气温-14.8℃,最热月7月平均温度9.8℃。年均降水量560 mm,主要集中于5~9月,占年降水量总和的80%,植物生长季,雨热同期。2012年和2013年生长季降水量(5~9月)分别为352.6 mm和381.8 mm,占全年降水量的89.1%和92.1%。年平均日照时数2 462.7 h,土壤类型为草毡寒冻雏形土,平均厚度0.65 m。样地为冬季牧场,自1982年以来均保持3只羊单位/hm2的放牧强度,生长季围栏封育,休眠季放牧[13]。
优势植物种为矮嵩草(Kobresiahumilis)、小嵩草(Kobresiapygmaea)、紫羊茅(Festucarubra.)、垂穗披碱草(Elymusnutans)、矮火绒草(Leontopodiumnanum)、雪白委陵菜(Potentillanivea)、线叶龙胆(Gentianafarreri)、早熟禾(Poaannua)、麻花艽(Gentianastraminea)、美丽风毛菊(Saussureapulchra)等,土壤基本理化性状见表1。

表1 矮嵩草草甸土壤基本理化性质
注:同列不同小写字母表示各土层间差异性显著(P<0.05)
1.2试验设计与方法
采用双因素交互试验设计,参照青藏高原东部大气氮沉降量数据[3][0.87~1.38 g/(m2·a)],分别设置施入2 g N/(m2·a)(NH4NO3)、增加20%降水、以及增氮和增水交互控制试验,各试验处理分别简写为N、Pre、N×Pre、CK(对照),分别模拟在氮沉降、降水增加气候情景下[6],高寒草甸主要群落动态潜在变化特征。在海北站地区,采用样线法,选择3个1 km×1 km样地,样地重复3次,每样地设置4个大小为1 m× 1 m的样方,分别为N、Pre、N×Pre、CK的样方,样方间隔带宽2 m;各处理随机进行设置,以消除微地形等环境异质性对试验结果的可能影响。3个样地,每处样设置(试验处理和对照)1个样方,3次重复。
试验于2012年生长季(5~9月)利用喷雾器在月初将氮肥均匀喷洒于样方内,对照样方则喷洒相同数量的水,以减少因外源添加水对草地生物地球化学循环的影响,生长季每次喷施水量不足0.5 mm。每年10月中旬将非生长季氮肥一次性施入土壤(因为模拟氮沉降情景,各月份所占比例相同)。降水量参考海北站地区以往30年平均降水量数据,分生长季和休眠季增加,其中,生长季按照月份在各月初添加,休眠季在10月一次性添加。模拟氮沉降和增水处理同期进行。
生物量采用标准样方法测定[13]。在2012和2013年生长季盛期(8月下旬),利用0.25 m×0.25 m样方,分种收获植物地上部分,于65℃烘干至恒重。
丰富度指数R=S
物种多样性指数的计算采用Shannon-Wiener(香农-威纳)指数表征:
式中:Pi为物种i的相对重要值;Pi=(相对高度+相对盖度)/2;S为种i所在样方的物种总数。
1.3数据分析
高寒草甸物种丰富度、地上生物量、物种多样性指数、功能群生物量对氮沉降、增水处理的响应间差异采用双因素方差分析。统计分析均利用SPSS 16.0(SPSS,Chicago,USA)实现,统计图形在Sigmaplot 10.0中绘制。
2 结果与分析
2.1氮沉降和增水对草地地上生物量的影响
模拟氮沉降和增水及其交互作用,当年均能够显著提高高寒草甸地上生物量(P<0.05),增加幅度分别为13.8%,22.0%和25.4%。与氮沉降处理相比,模拟增水对草地生产力提升的贡献稍高。对照样地生物量仅为(261.3±3.7)g/m2,交互作用处理样地地上生物量最高,达到(327.7±5.6)g/m2(P<0.01)。
在第2年,氮沉降、以及交互作用均提高了高寒草甸地上生物量,后者增加幅度为22.7%,达到极显著性检验水平(P<0.01),生物量为(314.2±6.9)g/m2。模拟增水处理并未明显增加地上生物量。与第1年相反,模拟氮沉降对草地生物量的提升效果优于增水处理。连续两年试验结果表明,模拟氮沉降与增水交互作用可以稳定地提升高寒草地生物量(图1)。
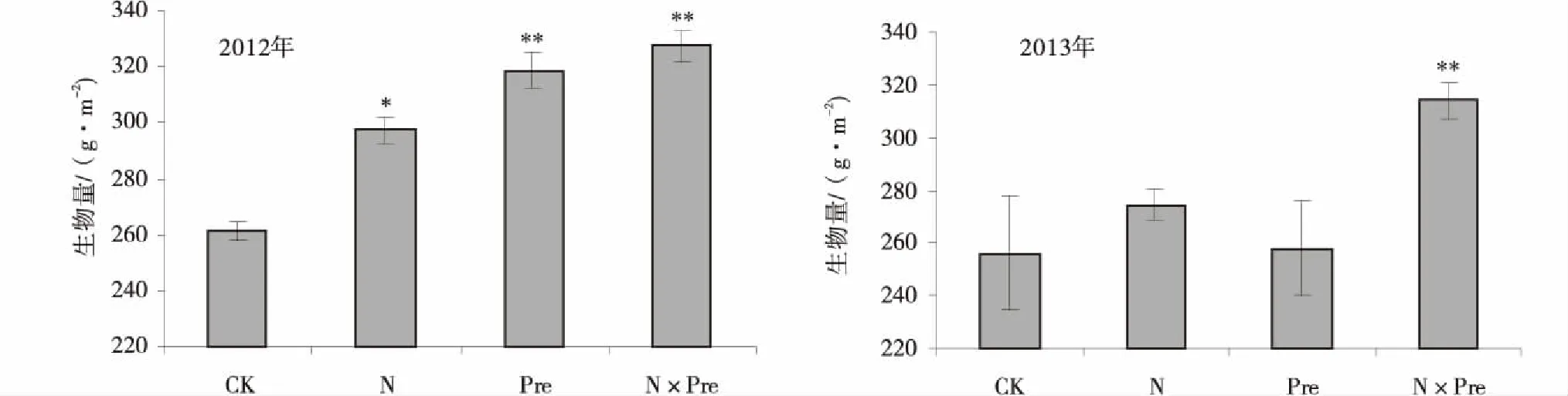
图1 高寒草甸地上生物量对氮沉降和增水的响应规律Fig.1 Effect of nitrogen deposition and adding precipitation on aboveground biomass in alpine meadow 注:*表示P<0.05;**表示P<0.01
2.2氮沉降与增水及交互作用对高寒草地不同功能群生物量的影响
氮沉降、增水及其交互作用处理当年均能够显著提高禾本科、豆科植物生物量(P<0.05),其中,增水处理禾本科植物生物量最高,为(215.2±6.57) g/m2,施肥与增水交互处理草地豆科植物生物量最高,达到(39.04±1.05)g/m2;显著的降低莎草科植物生物量(P<0.05),增水处理样地降低幅度最大,比对照降低1.66倍。氮沉降显著提高杂类草植物生物量,相比对照增加1.16倍(P<0.05),而增水显著降低杂类草生物量(P<0.05),降幅为26.4%。氮沉降处理显著降低凋落物输入量。
第2年氮沉降和增水及其交互作用均能够明显提高禾本科和莎草科植物生物量,禾本科植物生物量增加幅度分别为78.6%,84.6%和89.7%(P<0.05),氮沉降和交互作用促使莎草科植物生物量分别增加2.6和3.4倍(P<0.05),增水能够提高莎草科植物生物量约27.7%。氮沉降及交互作用略微降低了豆科植物生物量,但降水能够显著增加豆科生物量,比对照增加1.06倍(P<0.05)。各个处理均显著降低杂类草植物生物量,降低幅度依次为33.7%、48.3%和44.3%(P<0.05)。

图2 氮沉降和增水处理下高寒草甸不同功能群的生物量Fig.2 Effect of nitrogen deposition and adding precipitation on biomass of different function groups
2.3高寒草地物种丰富度对氮沉降与增水及交互作用的响应特征
增水能够显著提高高寒草甸物种丰富度,物种数目从18种增加到20种(P<0.05),对照样地中的小米草(Euphrasiapectinata)、发草(Deschampsiacaespitosa)、异叶米口袋(Tibetiahimalaica)、黄花棘豆(Oxytropisochrocephala)在增水样地中消失,而羊茅、早熟禾、黑穗薹草(Carexatrata)、湿生扁蕾(Gentianopsispaludosa)、地丁(Corydalisbungeana)和二裂委陵菜(Potentillabifurca)均有分布。第2年相比于对照样地,各处理物种数目均减少,氮沉降和交互作用显著降低了物种丰富度,物种数目从19种降低为13种(P<0.01)。对照样地分布的黄芪(Astragalusmembranaceus)和异叶米口袋,在氮沉降和交互作用样地消失。垂穗披碱草、羊茅、繁缕(Stellariamedia)、雅毛茛(Ranunculuspulchellus)、二裂委陵菜、矮火绒草、兰石草(Fructuslancea)和鹅绒委陵菜(Potentillaanserine)出现分布。
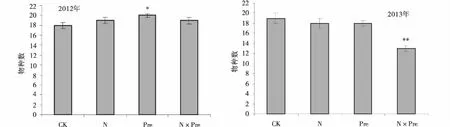
图3 氮沉降和增水处理下高寒草甸的丰富度Fig.3 Effect of nitrogen deposition and adding precipitation on plant richness in alpine meadow
2.4氮沉降与增水及交互作用对高寒草地生物多样性的影响
增水显著增加高寒草甸物种多样性指数,从2.63±0.18增加到2.83± 0.34(P<0.05),氮沉降和增水交互处理降低其多样性指数。而在第2年时,各处理均能引起高寒草地物种多样性降低,并且相比于对照,氮沉降和增水交互作用样地物种多样性指数从2.63±0.52降低到2.33±0.18,与其他处理达到极显著差异(P<0.01),氮沉降和增水及其交互作用在提升生产力的同时也造成物种丰富度和多样性降低。

图4 氮沉降和增水处理下高寒草甸的物种多样性指数Fig.4 Effect of nitrogen deposition and precipitation adding on biodiversity index in alpine meadow
3 讨论
通过连续两年观测,发现模拟氮沉降及其与增水的交互作用均能够明显增加高寒草地生产力,氮沉降和增水当年能够显著提高草地生物量,而在第2年其增产效率降低,尤其是增水处理效果较弱。已有研究报道,高剂量氮素添加能够缓解青藏高原高寒草甸植物生长的营养限制,提高植物地上生物量,幅度达到24%[4]。合理施肥能够促进草地植物根系发育,拓展了其对土壤水分的利用空间,在生长发育旺盛期能较好利用较深层土壤水分以满足对水分的需求[9]。降水量的变化直接导致了克氏针茅草原初级生产力的年度波动[14]。增加降水量明显提高高寒草地地上生物量,主要以禾草科生物量增加为主,禾本科植物如垂穗披碱草、异针茅等,其植株较高,处于群落的上层,可获得更丰富的光资源,根系主要为须根系,对水分、营养元素的竞争在群落中处于优势地位,从而其生长受到明显的促进[4],增加高寒草地牧草产量[15]。
研究发现,当年氮沉降、增水及交互作用能够提高高寒草甸物种丰富度,而第2年各处理物种丰富度均降低,氮沉降和交互作用显著降低了物种丰富度,消失和出现的物种均为禾本科和杂类草。已有研究发现,施肥导致资源供应的不对称性,改变地上、地下竞争强度,引起高寒草地物种丰富度下降17.3%,群落结构趋于简单[12,16-18],但也有研究发现,施肥提高高寒草地物种丰富度[18],前3年施肥未引起荒漠草地物种数目降低,但第4年显著降低物种丰富度[17]。高寒草甸不同物种存在氮素吸收能力和资源分配策略的权衡[8],物种之间对群落下层光的竞争也是资源添加导致物种丰富度降低的主要原因之一[10]。生长季降水丰富的羌塘东部地区物种丰富度最高约为24种,而在生长季降水贫乏的羌塘西部地区平均物种丰富度为7种[7]。
在海北站地区,增水能够显著增加高寒草甸生物多样性指数,氮沉降和增水交互处理降低其多样性指数。而在第2年各处理均能引起高寒草地物种多样性降低,氮沉降和增水交互作用样地物种多样性指数从2.63±0.52降低到2.33±0.18。已有研究表明达日县和曲麻莱县高寒草甸物种多样性最高,分别为2.74和2.27;唐古拉山乡和杂多县最低,约为1.5[13]。杨元武等[19]在祁连山高寒草甸研究结果报道多样性为0.65~3.73。肥水交互作用降低荒漠草地生物多样性,而养分和水分添加对其影响不显著[17]。内蒙古典型草原生态系统植物物种多样性随着氮素添加而降低,是由于杂类草的多样性降低所引起[20]。藏北高原高寒草地香农威纳多样性指数分布于0.92±0.05到2.19±0.07之间,随生长季降水减少而降低[7]。植物生物多样性与年降水量显著正相关,但当水资源过剩时,可能抑制其他物种的侵入和生长[17]。
高寒草甸生态系统群落特征对连续两年氮沉降和增水及交互作用的不同响应特征规律,可能也表现出高寒草地生态系统对外界干扰反应比较敏感,抵抗力偏低,但恢复力和稳定性较强。已有研究发现高寒草甸生态系统主要环境因子以3-4年的主周期随机低频振荡,生态系统的行为会呈现同主周期、振幅比较稳定的随机波动,高稳定性是长期处于较稳定的环境和系统适应环境的进化演替结果[21-23]。
4 结论
连续两年试验结果表明,模拟氮沉降与增水交互作用均可以稳定地提升高寒草地生物量,分别达到327.7和314.2 g/m2。连续两年进行氮沉降和增水及其交互处理,均能够明显提高禾本科生物量,增加幅度分别为78.6%,84.6%和89.7%,氮沉降和交互作用促使莎草科植物生物量分别增加2.6和3.4倍,为草地功能提升的重要途径。增水能够显著提高高寒草甸物种丰富度,物种数目从18种增加到20种,第2年各处理均降低物种数目,且莎草科植物(矮嵩草和小嵩草)抵抗外界干扰能力较强,可能是其成为优势植被的重要原因。
[1] 贺纪正,张丽梅.土壤氮素转化的关键微生物过程及机制[J].微生物学通报,2013,40(1):98-108.
[2] Liu X,Zhang Y,Han W.Enhanced nitrogen deposition over China[J].Nature,2013,494:459-462.
[3] L C,Tian H.Spatial and temporal patterns of nitrogen deposition in China:Synthesis of observational data[J].Journal of Geophysical Research:Atmospheres,2007,112:534-567.
[4] 杨晓霞,任飞,周华坤.青藏高原高寒草甸植物群落生物量对氮,磷添加的响应[J].植物生态学报,2013,38(2):159-166.
[5] Liu L,Greaver T L A.Global perspective on belowground carbon dynamics under nitrogen enrichment[J].Ecology Letters,2010,13(7):819-828.
[6] Piao S,Ciais P,Huang Y,etal.The impacts of climate change on water resources and agriculture in China[J].Nature,2010,467:43-51.
[7] 武建双,李晓佳,沈振西.藏北高寒草地样带物种多样性沿降水梯度的分布格局[J].草业学报,2012,21(3):17-25.
[8] 邓建明,姚步青,周华坤.水氮添加条件下高寒草甸主要植物种氮素吸收分配的同位素示踪研究[J].植物生态学报,2014,38(2):116-124.
[9] 沈振西,周兴民,陈佐忠.高寒矮嵩草草甸植物类群对模拟降水和施氮的响应[J].植物生态学报,2002,26(3):288-294.
[10] 张杰琦,李奇,任正炜.氮素添加对高寒草甸植物群落物种丰富度及其与生产力关系的影响[J].植物生态学报,2010,34(10):1125-1131.
[11] 杨中领,苏芳龙,苗原.施肥和放牧对青藏高原高寒草甸物种丰富度的影响[J].植物生态学报,2014,38(10):1074-1081.
[12] Wu Y B,Wu J,Deng Y C,etal.Comprehensive assessments of root biomass and production in aKobresiahumilismeadow on the Qinghai-Tibetan Plateau[J].Plant and soil,2011,338(1):497-510
[13] 杜岩功,崔骁勇,葛劲松.三江源地区高寒草地群落特征研究[J].草业科学,2010,27(3):9-14.
[14] 白永飞.降水量季节分配对克氏针茅草原群落初级生产力的影响[J].植物生态学报,1999,23(2):155-160.
[15] 王长庭,王启基,沈振西.模拟降水对高寒矮嵩草草甸群落影响的初步研究[J].草业学报,2003,12(2):25-29.
[16] 江小蕾,张卫国,杨振宇.不同干扰类型对高寒草甸群落结构和植物多样性的影响[J].西北植物学报,2004,23(9):1479-1485.
[17] 赵新风,徐海量,张鹏.养分与水分添加对荒漠草地植物群落结构和物种多样性的影响[J].植物生态学报,2014,38(2):167-177.
[18] 郑华平,陈子萱,王生荣.施肥对玛曲高寒沙化草地植物多样性和生产力的影响[J].草业学报,2007,16(5):34-39.
[19] 杨元武,李希来,祁盛仓.江河源地区不同荒漠化草地物种多样性研究[J].青海大学学报(自然科学版),2005,23(3):42-45.
[20] 白永飞,邢雪荣,许志信.内蒙古高原针茅草原群落β多样性研究[J].应用生态学报,2000,11(3):408-412.
[21] 李亚娟,孙灿灿,曹广民.三江源区不同利用方式草地生物量及土壤养分特征[J].草原与草坪,2016(4):48-53.
[22] 周华坤,周立,赵新全.青藏高原高寒草甸生态系统稳定性研究[J].科学通报,2006,51:63-69.
[23] 聂学敏,芦光新,鲁子豫.不同沙化高寒草地植物群落结构特征及土壤理化特性研究[J].草原与草坪,2016(4):73-77.
ResponseofplantcommunityonnitrogendepositionandaddingprecipitationinalpinemeadowonQinghai-TibetanPlateau
XU Qing-min1,ZHOU Geng2,GUO Xiao-wei3,CAO Ying-fang3,DU Yan-gong3
(1.CentreStationofQinghaiEnvironmentMonitoring,Xining810008,China;2.BureauofWeihaiAgriculturalBureau,Weihai264411,China3.NorthwestInstituteofPlateauBiology,ChineseAcademyofScience,Xining810008,China)
The effects of nitrogen deposition and adding precipitation,and its interaction on characteristics of plant community were studied for two years in alpine meadow at Haibei station of Chinese Academy of Sciences on Qinghai-Tibetan Plateau.The result indicated that the total aboveground biomass,biomass of gramineae and leguminosae were significantly increased in the first year.However,the biomass of sedge family was significantly decreased.Furthermore,the nitrogen deposition and its interaction with adding precipitation steadily raised the grassland production in the second year,but not for adding precipitation.All treatments could enhance the biomass of gramineae and sedge family but not for forbs.In addition,the precipitation increasing enhanced conspicuously species richness and biodiversity of alpine meadow in first year,but nitrogen deposition and its interaction decreased the biodiversity.The species richness and biodiversity index under all treatments decreased in second year.It could be concluded that nitrogen deposition and adding precipitation and its interaction increase the grassland production,and decrease the species richness and biodiversity.
nitrogen deposition;adding precipitation;alpine meadow;aboveground biomass;biodiversity
S 812
A
1009-5500(2017)05-0008-06
2016-12-20;
2017-01-03
国家自然科学基金(31470530和31200379);青海省自然科学基金(2016-ZJ-918Q)资助
许庆民(1983-),男,山东聊城人,在读硕士。
E-mail:403672585 @qq.com
杜岩功为通讯作者。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
活力(2019年21期)2019-04-01
河北林业科技(2016年5期)2016-11-08
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
