
不同干扰类型栲类次生林天然更新研究
2017-11-12 10:48孟家敏湖北省襄阳市林业局胡永辉湖北省国有鹿门寺林场
中国林业产业 2017年5期
孟家敏(湖北省襄阳市林业局) 胡永辉(湖北省国有鹿门寺林场)

福建省将乐国有林场的常绿阔叶林大多数是1958年皆伐后自然恢复生长的次生林,栲类次生林是当地常绿阔叶林主要森林类型之一。壳斗科植物是研究区栲类次生林的主要成分,樟科植物仅次于壳斗科。在人为干扰下,栲类次生林的演替或加速或暂停或逆向进行,可以通过对其主要建群树种天然更新状况的研究加以确定。
森林的天然更新是生态系统进行恢复和群落演替的重要手段之一。目前针对于森林天然更新的研究单一地集中在不同环境因子对更新幼苗多样性、数量以及分布格局等的影响,鲜有综合分析栲类次生林更新幼苗的分布格局并进行更新评价的研究。因此,本研究根据乔木层的物种多样性测度因子,结合研究区实际情况划分栲类次生林不同干扰类型,通过方差均值比法结合5种聚集强度参数测定更新幼苗的空间分布格局,并通过因子分析法对栲类次生林主要建群树种更新状况进行评价,旨在为研究区栲类次生林的恢复演替经营提供依据。
数据与方法
1.1 数据来源
于2013年7月在研究区内通过踏查选择立地条件相同或相似且具有代表性的栲类次生林典型地段,采用样地调查方法,对各样地进行每木检尺和更新调查。样地面积为20m×30m,共15个,对样地内乔木(胸径≥5cm)进行每木检尺,分别调查乔木的树种、树高、胸径、冠幅、枝下高等;分别在样地的四个角和中心选取更新样方(5m×5m),共75个,分别调查更新幼苗(胸径<5cm)的树种、苗高、地径(胸径)、密度以及盖度等。各样地基本情况如表1。

表1 栲类次生林样地基本情况表Table1 Basic situation of each plot of Castanopsis secondary forest
1.2 数据处理
1.2.1 物种多样性测度指标
统计各树种的株数、胸高断面积以及频度,计算重要值(Pi)。然后选取Shannon-Wiener物种多样性指数(SW)、Simpson多样性指数(D)、Simpson优势度指数(C)、Pielou群落均匀度(R)以及种间机遇机率Hurlbert指数(H)作为物种多样性测度指标,其计算公式如下:

式中,S为研究系统中物种总数;Ni指种i的个体数;N为所在群落全部种的个体总数。
1.2.2 更新幼苗的等级划分
依据幼苗的株高将其划分为5个等级:Ⅰ级(h≤30cm)、Ⅱ级(30cm<h≤60cm)、Ⅲ级(60cm<h≤100cm)、Ⅳ级(100cm<h≤200cm)、Ⅴ级(h>200cm,d≤7.5cm)。
2.2.3 幼苗的空间分布格局
运用方差均值比(V/m)法对更新幼苗的分布格局进行测定,并结合负二项分布参数(K)、丛生指标(I)、Cassie指数(Ca)、聚块性指标(m*/m)、扩散指标(Iδ)5种聚集强度参数加以验证,判别标准见表2。

表2 聚集度指标法各指标的判别标准Table2 Discriminating standard of each index in aggregation index method
1.2.4 幼苗的更新状况评价
首先,对15块样地数据做KMO和Bartlett检验,确定数据是否适合做因子分析;然后,用软件SPSS18.0通过“分析→降维→因子分析”处理15块样地数据,对每公顷幼苗株数、平均地径和平均苗高3个指标进行降维,用天然更新综合得分因子(F)对上述3个指标综合表示;最后,每个树种的得分情况反映其天然更新状况的优劣程度。
结果与分析
2.1 栲类次生林乔木层的物种多样性
分别计算15块样地乔木层的Shannon-Wiener物种多样性指数(SW)、Simpson多样性指数(D)等5种物种多样性测度指标(见表3)。
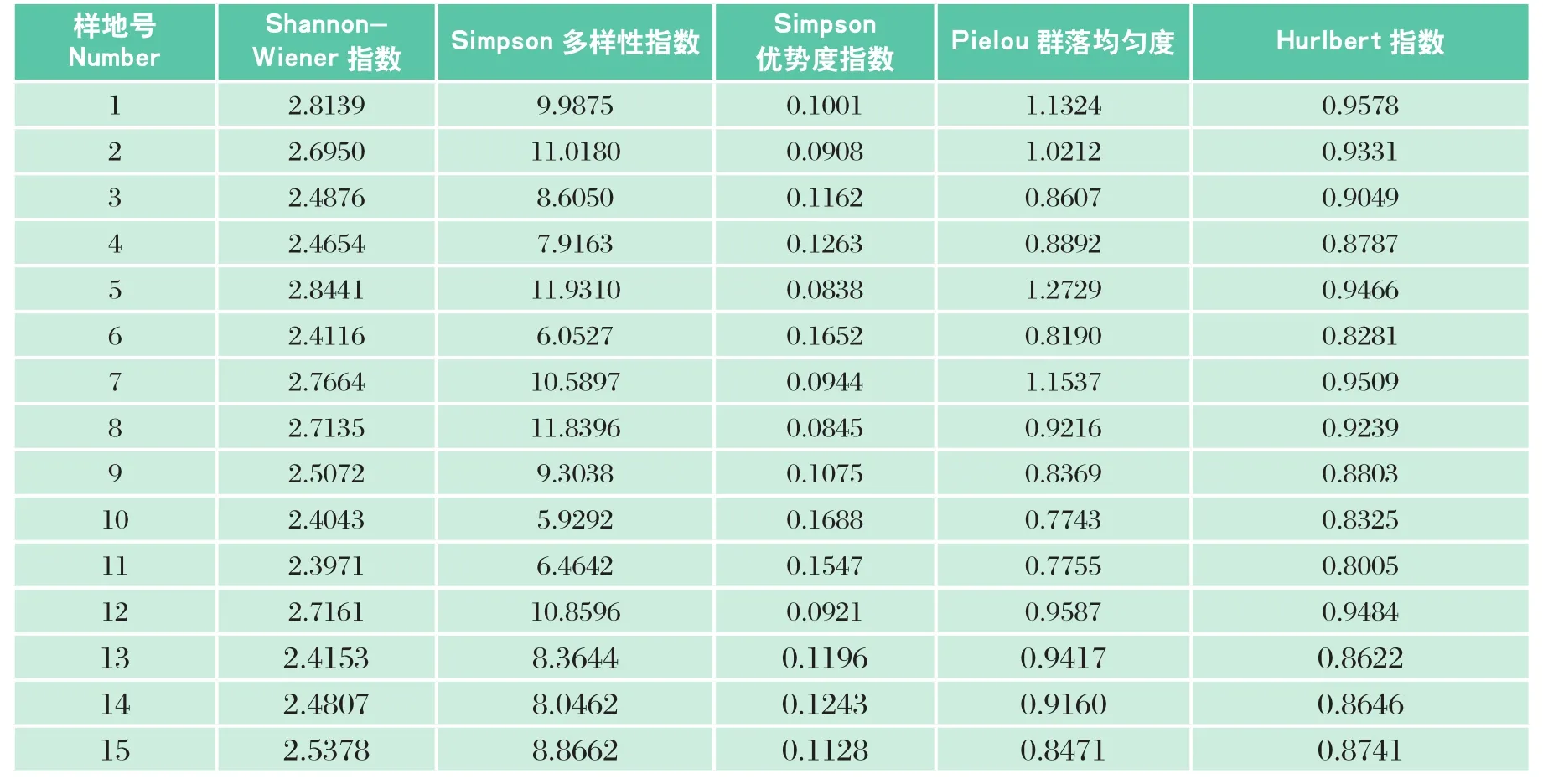
表3 栲类次生林乔木层的物种多样性Table3 Tree species diversity of Castanopsis secondary forest
研究区15块样地中,Shannon-Wiener多样性指数在 2.3971~2.8441,而福建省栲树顶级群落的Shannon-Wiener指数值是5.14,说明研究区栲类次生林尚未达到顶级状态;Simpson多样性指数在5.9292~11.9310,表明各样地的物种数目及个体分配的均匀度相差较大; Simpson优势度指数0.0838~0.1688,其中样地5最小,样地10最大,说明样地5物种数量分布较其它样地均匀,样地10的优势种地位最突出;Pielou群落均匀度刚好与之相反;Hurlbert指数在 0.8005~0.9578,各样地情况变化不大。
2.2 栲类次生林各样地干扰类型的划分
根据表3的物种多样性测度指标结果对15块样地进行数据进行K-均值聚类分析,并进行方差检验,结果见表4和表5。
根据故障原语描述[12],在低电压SRAM中注入写破坏耦合故障和读破坏耦合故障,仿真结果分别如图9、图10所示,写破坏耦合故障和读破坏耦合故障也能被March-Like算法检测出,而不能被March C+算法检测。

表4 基于Shannon-Wiener指数、Simpson多样性指数、Simpson优势度指数、Pielou群落均匀度指数和Hurlbert指数的栲类次生林各样地的K-均值聚类结果Table4 K-means clustering results of each plot of Castanopsis secondary forest based on Shannon-Wiener index、simpson diversity index、simpson dominance index、Pielou index and Hurlbert index

表5 栲类次生林各样地K-均值聚类结果的方差分析Table 5 Variance analysis of K-means clustering results of each plot in Castanopsis secondary forest
由表4可知,将乐林场栲类次生林15块样地可划分为三类,即{1,2,5,7,8,12 },{3,4,9,13,14,15 }和{6,10,11}。由表5可知,5个分类变量在3个类别中差异显著,因此分类结果具有可信度。
综上所述,15块栲类次生林样地可以划分为以下三个类型。
轻度干扰——类型Ⅰ(样地1,2,5,7,8,12):样地栲类树种的总胸高断面积所占比例为6成及以上。林分偶有人进入林内收集枯枝,乔木未被采伐,人为干扰轻微。
中度干扰——类型Ⅱ(样地3,4,9,13,14,15):样地栲类树种的总胸高断面积所占比例为4~5成。林分中有少量乔木被采伐,形成小面积林窗,人为干扰程度中度。
重度干扰——类型Ⅲ(样地6,10,11):样地栲类树种的总胸高断面积所占比例为3~4(不含)成。林分曾遭受大量的人为采伐,有较大面积的林窗,人为干扰程度严重。
从划分结果看,属于轻度和中度干扰类型的林分较多,与研究区实际情况相符。林分中优势树种所占的断面积比例可以作为划分不同干扰类型的主要标准之一。
2.3 不同干扰类型栲类次生林主要建群树种更新幼苗的平均密度分布
分别统计不同干扰类型各样地内主要建群树种不同等级幼苗的株数,计算不同等级的幼苗密度(表6)。由表6可以看出,轻度干扰林分中,壳斗科和樟科更新的平均幼苗株数密度为5320株/hm2,中度和重度干扰林分更新的幼苗株数密度分别为4364株/hm2和1856株/hm2,说明随着干扰强度的增加,栲类次生林主要建群树种更新幼苗株数呈不断减少的趋势。
随着干扰强度的增加,林分中各树种的株数密度大致呈现逐渐减少的趋势。与轻度干扰相比,中度干扰下栲树更新幼苗株数密度稍有增加,可能是由于栲树种子在林窗下的萌发率和幼苗成活率高于林下,而在重度干扰的林分中,栲树更新幼苗密度骤降,可能是因为大面积林窗下,过强的光线灼伤幼苗导致其大量死亡。此外,重度干扰林分中没有出现茅栗的更新幼苗以及米槠和刨花润楠的Ⅰ级、Ⅱ级和Ⅲ级的幼苗,除了受人为收集枯枝落叶等的影响外,还有可能是大面积的林窗导致的幼苗死亡或者无法正常完成天然更新。

表6 不同干扰类型栲类次生林主要建群树种天然更新各等级幼苗的株数密度Table6 Number density of domination species various grades seedling in different interference types of Castanopsis secondary forest单位:株/hm²

栲树Castanopsis fargesii 189 97 89 72 31 478 25.75青冈栎Cyclobalanopsis glauca 73 46 40 31 14 204 10.99苦槠Castanopsis sclerophylla 96 58 49 47 17 267 14.39壳斗科Fagaceae 米槠Castanopsis carlesii 0 0 0 48 11 59 3.18甜槠Castanopsis eyrei 49 30 26 25 13 143 7.70多穗石栎Lithocarpus polystachya 51 28 26 24 9 138 7.44茅栗Castanea seguinii 0 0 0 0 0 0 0重度干扰华南木姜子Litsea greenmaniana 34 21 20 20 6 101 5.44 Severe disturbance樟科Lauraceae黄润楠 Phoebe zhennan 78 46 40 37 12 213 11.48黄樟 Cinnamomum porrectum 63 33 24 20 2 142 7.65绒毛山胡椒Lindera nacusua 25 18 17 15 3 78 4.20刨花润楠Machilus pauhoi Kanehira 0 0 0 26 7 33 1.78总计Total 658 377 331 365 125 1856比例(%)Proportion 35.45 20.31 17.83 19.67 6.73 1
2.4 不同干扰类型栲类次生林更新幼苗分布格局
聚集强度参数测定结果(表7)表明,不同干扰类型栲类次生林更新幼苗的V/m均大于1,其他5种聚集度强度参数计算的结果分别为:K>0、I>0、Ca>0、m * /m>1、 Iδ>1,说明栲类次生林更新幼苗都处于聚集分布状态。且随着干扰强度的增加,更新幼苗的聚集强度呈现逐渐增大的趋势。较高的聚集程度表明种群处在侵入扩散阶段,当种群定居成功并不断繁殖使种群增长,种群的聚集程度就会减弱。可以表明,随着干扰强度的增加,栲类次生林的演替开始停滞或逆向进行。

表7 不同干扰类型栲类次生林主要建群树种更新幼苗的分布格局Table7 Spatial distribution of domination species seedlings of Castanopsis secondary forest in different interference types
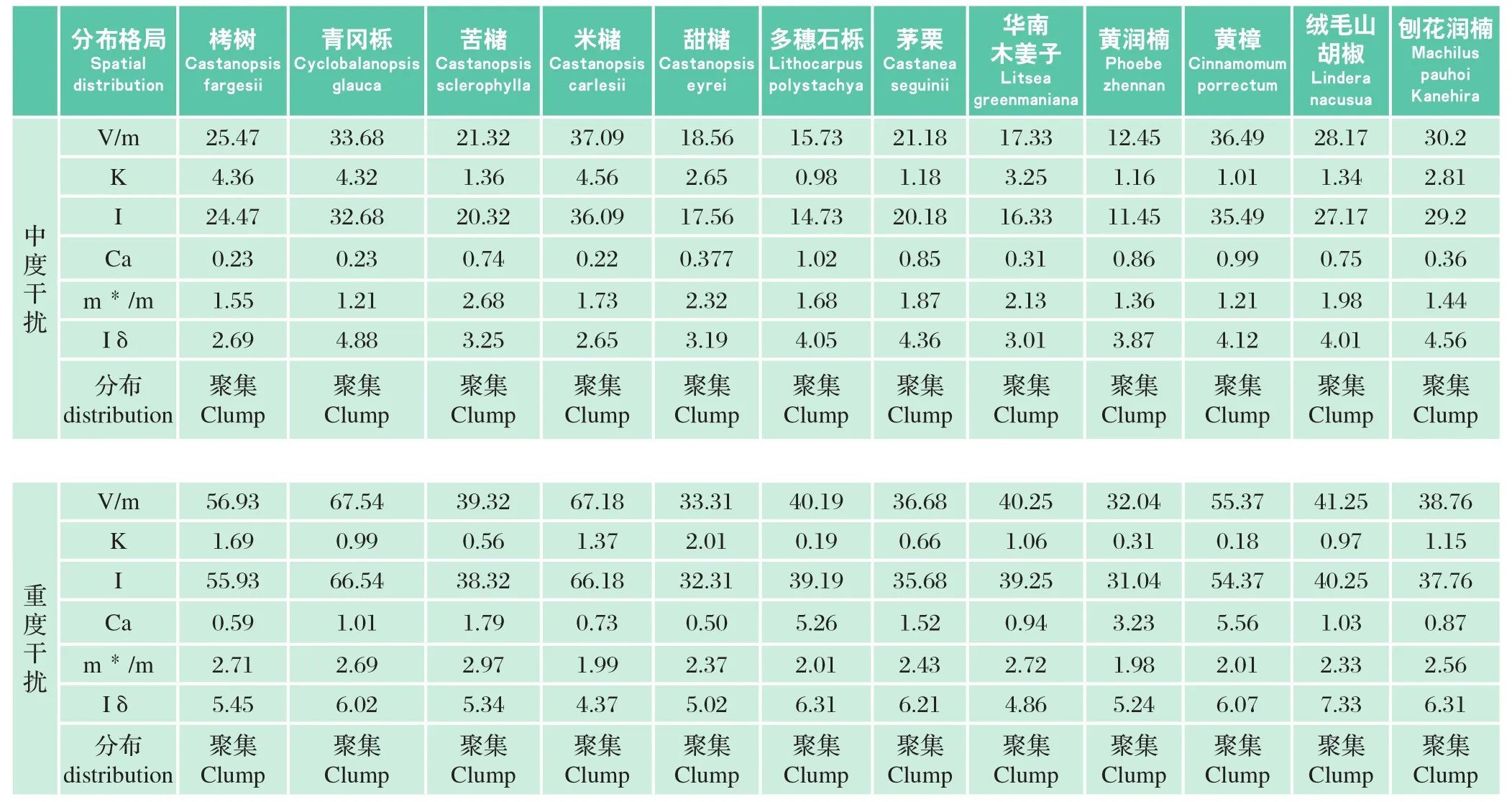
分布格局栲树青冈栎苦槠米槠甜槠多穗石栎茅栗黄润楠黄樟刨花润楠Spatial distribution Castanopsis fargesii Cyclobalanopsis glauca Castanopsis sclerophylla Castanopsis carlesii Castanopsis eyrei Lithocarpus polystachya Castanea seguinii华南木姜子绒毛山胡椒Litsea greenmaniana Phoebe zhennan Cinnamomum porrectum Lindera nacusua Machilus pauhoi Kanehira V/m 25.47 33.68 21.32 37.09 18.56 15.73 21.18 17.33 12.45 36.49 28.17 30.2 K 4.36 4.32 1.36 4.56 2.65 0.98 1.18 3.25 1.16 1.01 1.34 2.81 I 24.47 32.68 20.32 36.09 17.56 14.73 20.18 16.33 11.45 35.49 27.17 29.2 Ca 0.23 0.23 0.74 0.22 0.377 1.02 0.85 0.31 0.86 0.99 0.75 0.36 m*/m 1.55 1.21 2.68 1.73 2.32 1.68 1.87 2.13 1.36 1.21 1.98 1.44 Iδ 2.69 4.88 3.25 2.65 3.19 4.05 4.36 3.01 3.87 4.12 4.01 4.56分布distribution中度干扰聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump V/m 56.93 67.54 39.32 67.18 33.31 40.19 36.68 40.25 32.04 55.37 41.25 38.76 K 1.69 0.99 0.56 1.37 2.01 0.19 0.66 1.06 0.31 0.18 0.97 1.15 I 55.93 66.54 38.32 66.18 32.31 39.19 35.68 39.25 31.04 54.37 40.25 37.76 Ca 0.59 1.01 1.79 0.73 0.50 5.26 1.52 0.94 3.23 5.56 1.03 0.87 m*/m 2.71 2.69 2.97 1.99 2.37 2.01 2.43 2.72 1.98 2.01 2.33 2.56 Iδ 5.45 6.02 5.34 4.37 5.02 6.31 6.21 4.86 5.24 6.07 7.33 6.31分布distribution重度干扰聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump聚集Clump
2.5 不同干扰类型栲类次生林主要建群种更新状况评价
对15块样地数据做KMO和Bartlett检验,结果显示KMO统计量为0.54,Bartlett检验显著性水平为0.00,表明数据适合做因子分析。由因子分析结果(表8)可以看出,前两个成分的累积方差贡献率达到99.62%,前2个主成分包含了预设指标体系中的绝大多数信息。因此,采用主成分分析法提取前两个因子,分别定义为 F1,F2。

表8 栲类次生林主要建群树种的因子分析Table8 Factor analysis method for domination species in Castanopsis secondary forest
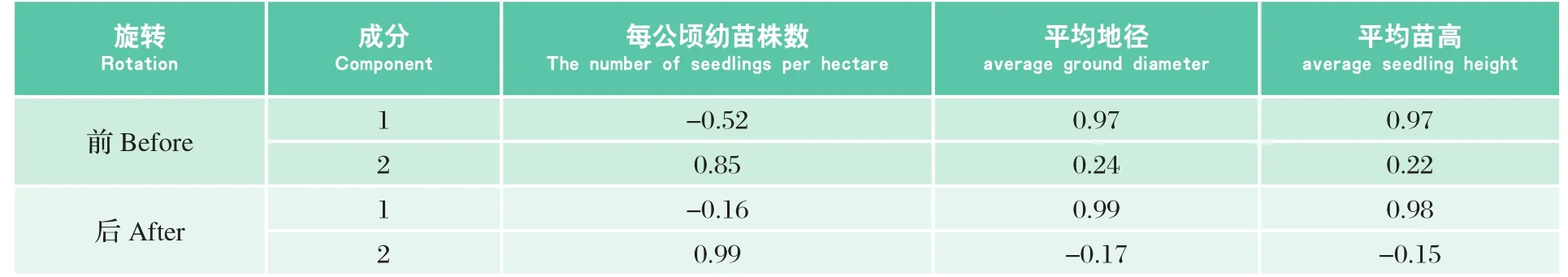
表9 各评价指标旋转前后的因子载荷矩阵Table9 Factor loading matrix of evaluation index before and after rotation
从旋转前后的因子载荷矩阵(表9)可以看出,采用最大方差法进行旋转以后,F1和F2的因子载荷更为集中。其中,F1包含了平均地径的0.99的信息,平均树高的0.98的信息,因此将F1定义为幼苗生长因子;F2包含了每公顷林木株数的0.99的信息,因此将F2定义为幼苗密度因子。再以旋转后提取因子方差贡献率为权重,可得到天然更新综合得分因子(F)(公式(7))。为了便于比较,在各数值大小关系不变的前提下,将天然更新综合得分因子(F)换算成评价指数(P)(公式(8)),范围在0~10之间。P值越大表明更新状况越好。
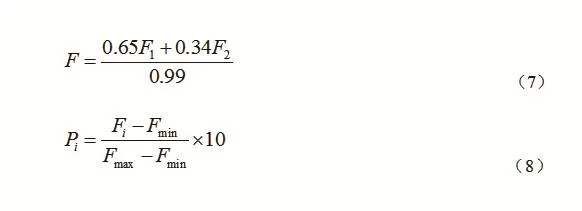

分别计算不同干扰类型栲类次生林主要建群树种的更新评价指数(P),生成柱状图(图1)。由图1可以看出,中度干扰下栲树更新评价指数最大,为10;重度干扰下刨花润楠更新评价指数最小,为0。就每个树种而言,轻度干扰的P值最大,中度干扰次之,重度干扰最小。而栲树在中度干扰时的更新状况优于轻度干扰,是由于小面积的林窗增加了群落下层的光照,有利于栲树更新幼苗的成活。
结论与讨论
天然更新是次生林持续发展的主导因子,木本植物幼苗更新状况直接影响着植物种群的数量及其分布格局,最终可能影响到森林群落的演替过程。栲类次生林是研究区常绿阔叶林的主要森林类型之一,研究其在不同干扰条件主要建群树种的幼苗更新状况对于确定栲类次生林的演替方向具有重要意义。
1)福建将乐林场栲类次生林乔木层的Shannon-Wiener多样性指数为2.3971~ 2.8441;Simpson多 样 性 指 数在5.9292~11.9310;Simpson优势度指数为0.0838~0.1688;Pielou群落均匀度 为 0.7743~ 1.2729;Hurlbert指 数 在0.8005~0.9578。研究区栲类次生林尚未达到顶级群落状态。
2)以上述5个物种多样性测度指标对研究区栲类次生林进行K-均值聚类,最终将研究区15块样地划分为3个类型:轻度干扰{样地 1,2,5,7,8,12 },中度干扰{样地 3,4,9,13,14,15 }和重度干扰{样地6,10,11},划分结果与研究区实际情况相符。
3)研究区栲类次生林壳斗科和樟科的幼苗幼树株数密度随着幼苗等级的增加而逐渐减少,呈现明显的倒“J”型。随着干扰强度的增加,各树种更新幼苗呈现逐渐减少的趋势,栲树除外。中度干扰林分中栲树更新幼苗的株数密度多于轻度干扰,这与中度干扰林分中形成的小面积林窗有关。
4)运用方差均值比(V/m)法对更新幼苗的分布格局进行测定,并结合负二项分布参数(K)、丛生指标(I)等5种聚集强度参数加以验证。不同干扰类型栲类次生林更新幼苗的V/m均大于1,其他5种聚集度强度参数计算的结果分别为:K>0、I>0、Ca>0、m*/m>1、 Iδ>1,说明栲类次生林更新的幼苗都处于聚集分布状态。且随着干扰强度的增加,更新幼苗的聚集强度呈现逐渐增大的趋势。
5)采用因子分析法对不同干扰类型栲类次生林更新状况进行评价。中度干扰林分中的栲树更新状况最优,重度干扰林分中的刨花润楠更新状况最差。就每个树种而言,轻度干扰时更新较优,中度干扰次之,重度干扰最差。由于小面积林窗等的影响,栲树在中度干扰时的更新状况优于轻度干扰。
研究区栲类次生林干扰主要来自于人为不合理的采伐,随着干扰强度的增加,栲类次生林主要建群树种更新幼苗的株数密度、空间分布格局以及更新状况呈现明显的规律性,即随着干扰强度的增加,主要建群树种的株数密度逐渐减少,空间分布聚集程度逐渐增加,更新状况逐渐变差。重度干扰条件下,主要建群树种更新幼苗的总株数仅为1856株/ hm2,远不能满足天然更新的需求。在以后的经营活动中,在减少人为不合理采伐的前提下,可结合人工辅助更新,使林分能够保持正向演替。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
农业科技与信息(2022年9期)2022-06-18
中国麻业科学(2021年6期)2022-01-21
防护林科技(2020年6期)2020-08-12
临床检验杂志(电子版)(2020年1期)2020-04-03
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
现代农业研究(2017年11期)2018-01-12
现代园艺(2017年20期)2017-02-03
学苑创造·B版(2015年12期)2016-06-23
