
自然越冬过程中扁桃枝条生理特性变化与抗寒性评价
2017-11-01 12:01王绪春王振磊徐崇志王新建林敏娟
新疆农业科学 2017年10期
欧 欢,王绪春,王振磊,3,徐崇志,3,王新建,3,林敏娟,3
(1.塔里木大学植物科学学院/新疆生产建设兵团南疆特色果树生产工程实验室,新疆阿拉尔 843300;2.南疆特色果树高效优质栽培与深加工技术国家地方联合工程实验室,新疆阿拉尔 843300;3.新疆生产建设兵团塔里木盆地生物资源保护与利用重点实验室,新疆阿拉尔 843300)
自然越冬过程中扁桃枝条生理特性变化与抗寒性评价
欧 欢1,2,王绪春1,2,王振磊1,2,3,徐崇志1,2,3,王新建1,2,3,林敏娟1,2,3
(1.塔里木大学植物科学学院/新疆生产建设兵团南疆特色果树生产工程实验室,新疆阿拉尔 843300;2.南疆特色果树高效优质栽培与深加工技术国家地方联合工程实验室,新疆阿拉尔 843300;3.新疆生产建设兵团塔里木盆地生物资源保护与利用重点实验室,新疆阿拉尔 843300)
目的研究6个扁桃品种的枝条在自然越冬过程中生理特性变化,综合评价不同扁桃枝条抗寒性的大小,为扁桃引种栽培及推广提供理论基础。方法测定越冬过程中各扁桃枝条膜透性、保护酶活性和渗透调节物质含量,分析生理指标在越冬期间的变化及相关性,比较不同品种在越冬期间的差异。结果在自然越冬过程中,扁桃枝条电解质渗透率、MDA含量、SOD活性、POD活性、可溶性蛋白质含量、可溶性性糖含量及脯氨酸量均呈先上升后下降的趋势,且与冬季温度均为负相关关系;温度变化对电解质渗透率、MDA含量、SOD活性、可溶性蛋白质和可溶性糖含量变化影响较大,对POD活性和脯氨酸含量变化影响较小,且抗寒性强的品种较抗寒性弱的品种增幅大,其中浓帕烈在越冬期间各项生理指标变化显著高于其它品种。结论通过测定扁桃枝条在越冬过程中各生理指标,发现电导率、SOD、可溶性糖和蛋白质均可作为评价抗寒强弱的指标,并结合隶属函数关系综合评价6个扁桃品种抗寒性的大小依次为浓帕烈>桃巴旦>矮丰>苦巴旦>晚丰>小软壳。
扁桃;自然越冬;抗寒性;生理指标;综合评价
0 引 言
【研究意义】扁桃(AmygdalusCommunisL.)又称巴旦杏系蔷薇科李亚科桃属扁桃亚属落叶乔木。其营养价值高,其中蛋白质含量最高可达 28%,油脂含量在47%~61%左右[1],还含有人体必需氨基酸、糖、无机盐、维生素和钙、镁、钠、铁、硒、锌等营养元素。药用方面,巴旦杏仁具有明目、健脑、健胃和助消化的功能,能治疗多种疾病,尤其是在治疗肺炎、支气管炎等呼吸道疾患上疗效显著,苦杏仁还用于制成镇静剂和止痛剂等[1,2]。扁桃仁可鲜食,炒食,制糕点,糖果,干果罐头等[3],与核桃、阿月浑子、榛子并称为“世界四大干果”。 扁桃种植在世界分布较广,主要分布在中国、美国、俄罗斯、西班牙、意大利、葡萄牙、法国、土耳其、伊朗等30多个国家和地区,其中美国栽植面积及产量居世界之首[4]。我国扁桃的种植已有1300年的历史,主要分布在新疆维吾尔自治区,青海、宁夏、甘肃、陕西、河北、山东等省也有少量引种栽培[5],2014年,新疆扁桃栽培面积为7.5 hm2,总产量5.4×104t,总产值14.18×108元[6]。在冬季越冬中,扁桃能够抵抗-20℃的低温,但是-20~-24℃时,会出现的冻梢现象,花期只能耐4℃的短时低温[7,8],因此扁桃的栽培推广受到了限制,只有新疆喀什有少量种植。【前人研究进展】见王琳等[9]在人工低温胁迫下对几个扁桃品种的抗寒生理变化进行的研究,在自然越冬过程中曾见杨晓静等[10]对扁桃有相关报道,但对扁桃在自然越冬过程中进行系统的膜透性、保护酶、渗透物的生理特性研究较少。【本研究切入点】目前,有关扁桃抗寒性方面研究报道较少。研究自然越冬过程不同扁桃品种枝条的膜透性、保护酶活性及渗透调节物质的变化,结合隶属函数关系,综合评价其抗寒性的大小。【拟解决的关键问题】分析6个扁桃品种枝条在越冬过程中各生理指标的变化及各指标与抗寒之间的关系,综合评价不同扁桃枝条抗寒性的大小,为扁桃引种栽培及推广提供理论基础。
1 材料与方法
1.1 材 料
试验材料采集于塔里木大学园艺试验站扁桃种质资源圃中,材料为均为莎车引进品种,树龄均为12a生树。于2016年11月15日至2017年3月1日每18 d采一次样。供试品种为桃巴旦,矮丰,苦巴旦,小软壳,晚丰,浓帕烈。采集选择生长健壮、发育正常、充分成熟的1a生休眠枝,尽量在选定植株的不同方位不同层次选取粗细均匀的枝条,每个品种剪取8~10 根,剪取的枝条长15~20 cm,而后分别贴好标签,装入聚乙烯袋内带回实验室。在实验室内用自来水冲洗,用蒸馏水冲洗3次后,用干净干燥的纱布擦干水分,分装于塑料袋中,用于测定电导率的样品置于冰箱4 ℃中保存备用,生理指标测定样品用液氮速冻后置于-82℃超低温冰箱保存。
1.2 指标测定
电导率测定采用电导法[11];试验生理指标测定参照李合生[12]实验指导,方法略有改动。丙二醛(MDA)含量的测定采用采用硫代巴比妥酸(TBA)法,过氧化物酶(POD)活性测定采用采用愈创木酚法,超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)法,蛋白质含量测定采用考马斯亮蓝G-250染色法测定,可溶性糖含量采用硫酸蒽酮法,脯氨酸含量采用茚三酮比色法,每个指标平行测定3次。
1.3 数据处理
利用DPS7.05数据处理软件处理;综合评价采用隶属函数值法[13]。
2 结果与分析
2.1 采样地环境温度变化
研究表明,采样地环境的最高温度、最低温度从2016 年 11 月15至 2017年1月7日呈下降趋势。2017年1月7日至1月24日处于低温状态,温度变化不大。自然越冬期最低气温出现在 2017年 1月 7 日左右,最低温度为 -15 ℃。从 2017 年 1 月24日之后温度开始逐渐回升, 3 月 1 日上升至 -4℃。图1

图1 采样时间及日最高和最低温度
Fig.1 The highest and lowest day temperature of sampling date
2.2自然越冬过程中不同扁桃枝条膜透性变化
2.2.1 电解质渗透率变化
研究表明,6个扁桃品种在整个越冬期间同自然冬季温度变化相反,呈先上升后下降的趋势。桃巴旦、苦巴旦、矮丰、浓帕烈和小软壳从11月15日到1月7日,随着温度的降低,电解质渗透率增加,可能是由于温度下降细胞膜组织受到伤害;且均在1月7日出现峰值,较休眠初期分别增加了33.22%、23.72%,26.67%、29.77%、20.53%。1月24日至3月,温度有缓慢回升的趋势,相应的电解质渗透率下降。但是晚丰峰值出现在1月24日,较休眠初期期增加了20.53%。在温度度最低时期浓帕烈与桃巴旦差异不显著,与其他品种间存在显著性差异,桃巴旦与苦巴旦差异不显著,矮丰与小软壳差异不显著。温度对其影响较小可能是其保护酶活性及渗透调节物质含量的增加相关。图2

注:不同字母表示相同时期内不同品种在P<0.05 水平上差异显著
Note: different letters indicated significant difference inP<0.05 level at the same time
图2 越冬过程中扁桃枝条电导率变化
Fig.2 Changes of relative conductivity of six almond cultivars during
2.2.2 MDA含量变化
研究表明,6个扁桃品种枝条在自然越冬过程中MDA含量与自然冬季气温相对应,呈现上升后下降的趋势。矮丰、苦巴旦、浓帕烈和小软壳随温度变化而变化,并在1月7日出现峰值值,较休眠初期分别增加了61.10%、44.96%、80.80%和44.71%;其中含量最高及变化最大的均为浓帕烈。而晚丰在1月24日出现峰值,较休眠初期增加了49.00%。但是桃巴旦在刚入冬期间其MDA含量的变化与其它品种不同,从11月15日到12月3日自然气温下降,其MDA含量下降;从12月3到2月8日MDA含量逐渐增加,较休眠初期增加了114.34%。随后随着温度的增加其含量下降,可能是受前期低温影响较大,突然温度升高细胞膜受损,MDA含量增加。在冬季最低温度时期各品种间存在显著性差异,其中浓帕烈含量最高,其含量受低温度影响最大,晚丰和苦巴旦差异不显著。图3
2.3自然越冬过程中不同扁桃品种保护酶变化
2.3.1 SOD活性变化
研究表明,6个扁桃品种枝条在越冬过程中SOD活性的变化与冬季自然温度相对应,SOD酶活性大致呈先上升后下降的趋势。晚丰、矮丰、桃巴旦、浓帕烈随温度的变化而变化,晚丰、矮丰、浓帕烈在1月7日出现峰值,较休眠初期分别增加了38.65%、57.78%、52.96%。而桃巴旦则在1月24日出现峰值,较休眠初期增加了56.53%。但是苦巴旦和小软壳在从11月15日至12月21日呈上升趋势,且在12月21日出现峰值,较休眠初期分别增加了23.36%和34.265,但是到1月7日SOD活性下降,可能是持续低温增加了膜脂过氧化程度,导致植物体内的酶促防御系统受到危害,造成未得到及时清除活性氧的积累,从而降低了 SOD 活性。在最低温期间,矮丰与浓帕烈差异不显著,与其它品种间差异显著,晚丰与苦巴旦、小软壳差异不显著,但与桃巴旦差异显著;桃巴旦与苦巴旦、小软壳差异不显著;其中SOD酶活性及变化最高的是矮丰。图4
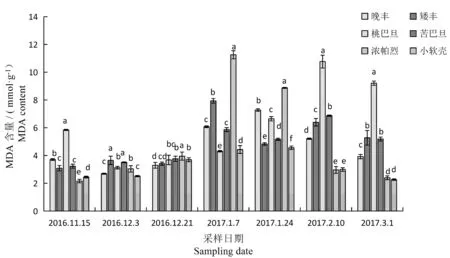
图3 越冬过程中6个扁桃品种MDA含量变化
Fig.3 Changes of the MDA content of six almond cultivars during natural overwintering

图4 越冬过程中6个扁桃品种SOD活性变化
Fig.4 Changes of the SOD activity of six almond cultivars during natural overwintering
2.3.2 POD活性变化
研究表明,POD活性变化与自然温度变化相对应,变化趋势也呈先上后下降的趋势。在整个自然越冬间,晚丰、矮丰、桃巴旦、苦巴旦、小软壳随自然温度的变化而变化,均呈先上升后下降的趋势,其中莎车 11 号、莎车 1 号、莎车 13 号在从 11 月 15 日到1月24日呈上升趋势,并在 1月 24 日 POD 活性最高,较休眠初期分别增加了21.10%、 49.63%、30.05%,而晚丰和小软壳在2月10日POD酶活性最高,较休眠初期分别增加了47.28%和72.28%,但是浓帕烈枝条在越冬期间与温度变化不一致,从11月15日到12月21日随着温度的降低,POD活性增加,并出现一个峰值,较休眠初期增加了64.15%,从11月21日到1月7日其活性迅速下降,从1月7日到2月10日其活性又呈上升趋势,并在2月10日出现第二个高峰值,较温度最低时增加了36.52%。之后随温度升高呈下降趋势。图5
2.4自然越冬过程中不同扁桃品种渗透调节物质变化
2.4.1蛋白质含量变化
研究表明,6个扁桃品种的枝条在自然越冬过程中蛋白质的含量变化与自然气温变化一致,均呈先上升后下降的趋势。晚丰、矮丰、浓帕烈均在11月15日至1月7日随着低温的不断加剧,可溶性蛋白含量在逐渐积累,并且在1月7日含量最高,较休眠初期分别增加了42%、45%、77%。之后,随着温度的回升,可溶性蛋白含量明显下降。矮丰峰值出现在12月21日,较休眠初期增加了11%,苦巴旦和莎车14蛋白质含量在1月24日最高,较休眠初期分别增加了50%和48%,之后均呈缓慢下降趋势。桃巴旦与苦巴旦差异不显著,小软壳可溶性蛋白含量最低,且与其他个品种间差异显著。图6
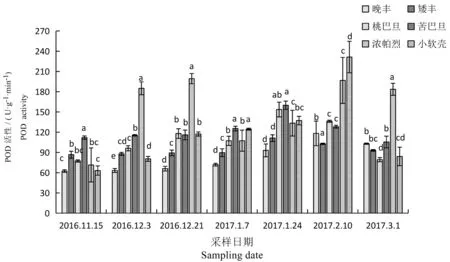
图5 越冬过程中6个扁桃品种POD活性变化
Fig.5 Changes of the POD activity of six almond cultivars during natural overwintering

图6 越冬过程中6个扁桃品种蛋白质含量变化
Fig.6 Changes of the Soluble protein content of six almond cultivars during natural overwintering
2.4.2 可溶性糖含量变化
研究表明,自然越冬条件下,6个扁桃品种的枝条在的可溶性糖含量随温度变化呈先上升后下降的趋势,不同品种变化幅度不同,晚丰、矮丰、桃巴旦、浓帕烈从11月15日到翌年2月10日呈上升趋势,并在2月10日出现最大值,其可溶性糖的含量分别增加了68.3%、55.3%、44.8%、56.7%;而苦巴旦和小软壳的最大值出现在翌年1月26日,其含量分别增加了44.8%和60.2%。在冬季温度最低时不同品种枝条可溶性糖含量不同,浓帕烈枝条可溶性糖含量最高(11.71%),与晚丰、矮丰和小软壳枝条可溶性糖含量差异显著。与桃巴旦、苦巴旦枝条可溶性糖含量差异不显著;小软壳与其他品种间均存在有显著性差异,其可溶性糖含量最低为9.17%。在各个不同时期,桃巴旦枝条的可溶性糖含量较其它品种而言相对较高。图7

图7 越冬过程中6个扁桃品种可溶性糖含量变化
Fig.7 Changes of the soluble sucrose content of six almond cultivars during natural overwintering
2.4.3 脯氨酸含量变化
研究表明,6个扁桃品种的枝条在越冬期间脯氨酸的变化随自然温度变化而变化,大致呈先上升后下降的趋势,但不同品种间的变化趋势及幅度不同。晚丰、桃巴旦、浓帕烈、和小软壳均为先上后下降的趋势。晚丰变化趋势不明显,在12月26号出现峰值,较休眠初期增加了56%;桃巴旦和浓帕烈在2月10日出现峰值,较休眠初期分别增加了77.90%和79.44%;小软壳在1月26日脯氨酸含量出现最大值为0.27%,比刚入休眠期时增加了80.35%。矮丰和苦巴旦均呈上升趋势,并在3月1日出现峰值,较休眠初期分别增加了85.04%和76.24%。在温度最低时可看出,浓帕烈与其它品种差异显著,晚丰和矮丰差异不显著,矮丰与小软壳差异不显著,与其它品种差异显著。浓帕烈在任何一个时期其脯氨酸含量明显高于其它品种。图8
2.5 生理指标与抗寒性关系
研究表明,自然越冬过程中最低温度与其生理特性指标变化均呈负相关关系,与相对电导率呈极显著负相关,与SOD活性、可溶性蛋白质含量、可溶性糖含量呈显著性负相关关系,说明在自然越冬过程中,扁桃枝条的生理特性随着自然温度下降其值相应上升,温度上升其值相应减小。相对电导率与MDA、可溶性蛋白、可溶性糖呈极显著正相关,MDA含量与可溶性蛋白、可溶性糖、脯氨酸呈显著性正相关,POD活性与脯氨酸含量呈显著性正相关关系,可溶性蛋白与可溶性糖呈极显著正相关关系。表1

图8 越冬过程中6个扁桃品种脯氨酸含量变化
Fig.8 Changes of the proline content of six almond cultivars during natural overwintering表1 不同抗寒性指标相关性
Table 1 The analysis correlation of different cold resistance index
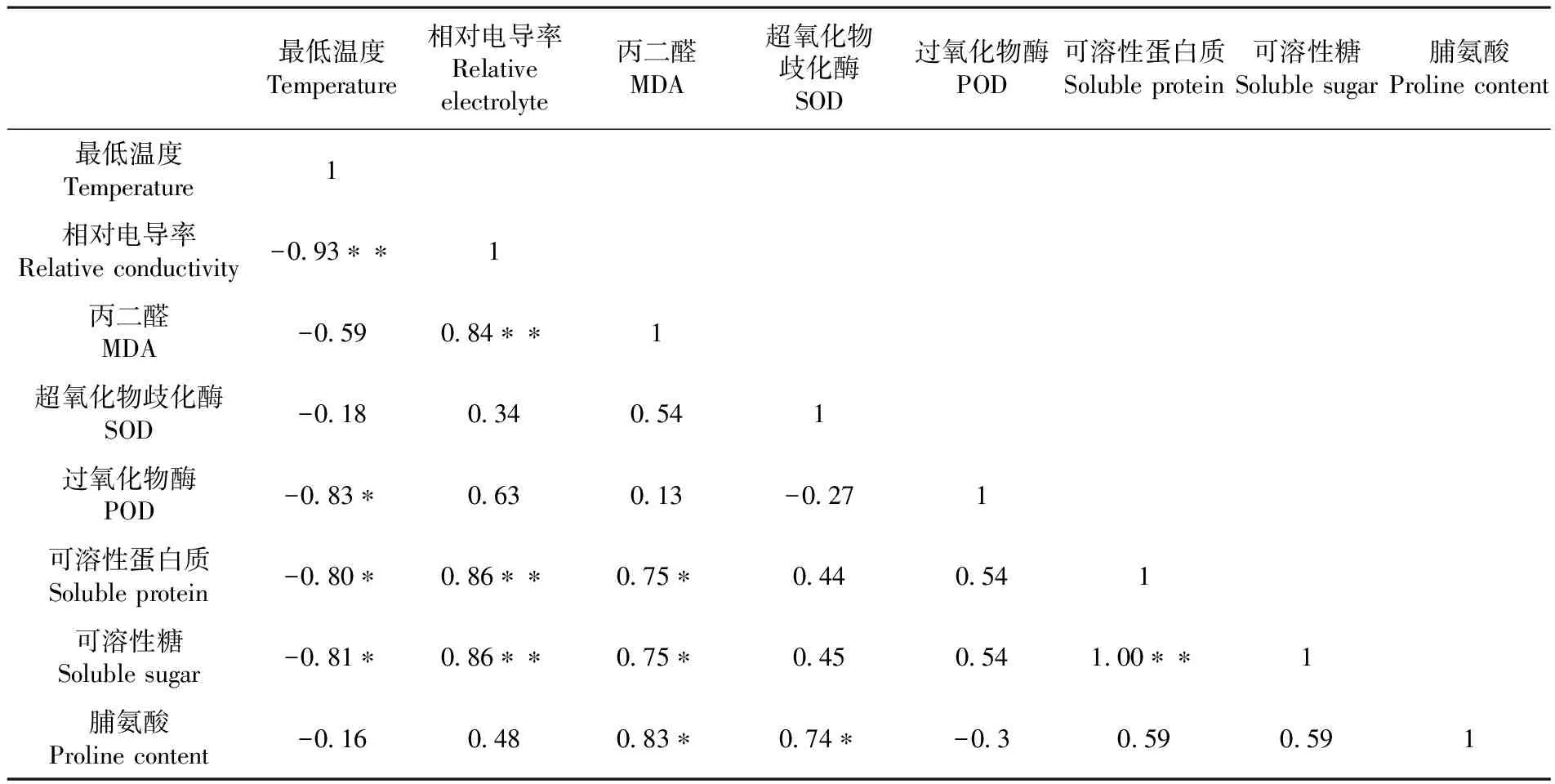
最低温度Temperature相对电导率Relativeelectrolyte丙二醛MDA超氧化物歧化酶SOD过氧化物酶POD可溶性蛋白质Solubleprotein可溶性糖Solublesugar脯氨酸Prolinecontent最低温度Temperature1相对电导率Relativeconductivity-093∗∗1丙二醛MDA-059084∗∗1超氧化物歧化酶SOD-0180340541过氧化物酶POD-083∗063013-0271可溶性蛋白质Solubleprotein-080∗086∗∗075∗0440541可溶性糖Solublesugar-081∗086∗∗075∗045054100∗∗1脯氨酸Prolinecontent-016048083∗074∗-030590591
注:*P<0.05显著相关,**P<0.01极显著相关
Note:Single asterisk meant significant correlation at 0.05 level,double asterisks meant extremely significant correlation at level
2.6 越冬过程中不同扁桃品种抗寒性评价
采用模糊数学隶属函数的方法,对与扁桃枝条自然越冬期间细胞膜透性、保护酶活性、渗透调节物质的含量作为越冬期间抗寒适应性综合能力指数进行评价。晚丰、矮丰、桃巴旦、苦巴旦、小软壳、浓帕烈的隶属度平均值分别为0.27、0.50、0.74、0.40、0.75、0.14;综合评价结果最高的是浓帕烈,其次是桃巴旦,两者相差不大,综合评价结果最低的是小软壳。因此可根据扁桃枝条在自然越冬期间细胞膜透性、保护酶活性、渗透调节物质的含量的综合评价结果来看,可综合评定这6个扁桃品种越冬抗寒性的大小,为浓帕烈>桃巴旦>矮丰>苦巴旦>晚丰>小软壳。表2
表2 6个扁桃品种越冬抗寒性评价
Table 2 Synthetic evaluation of the cold resistance character of almond six species during overwintering
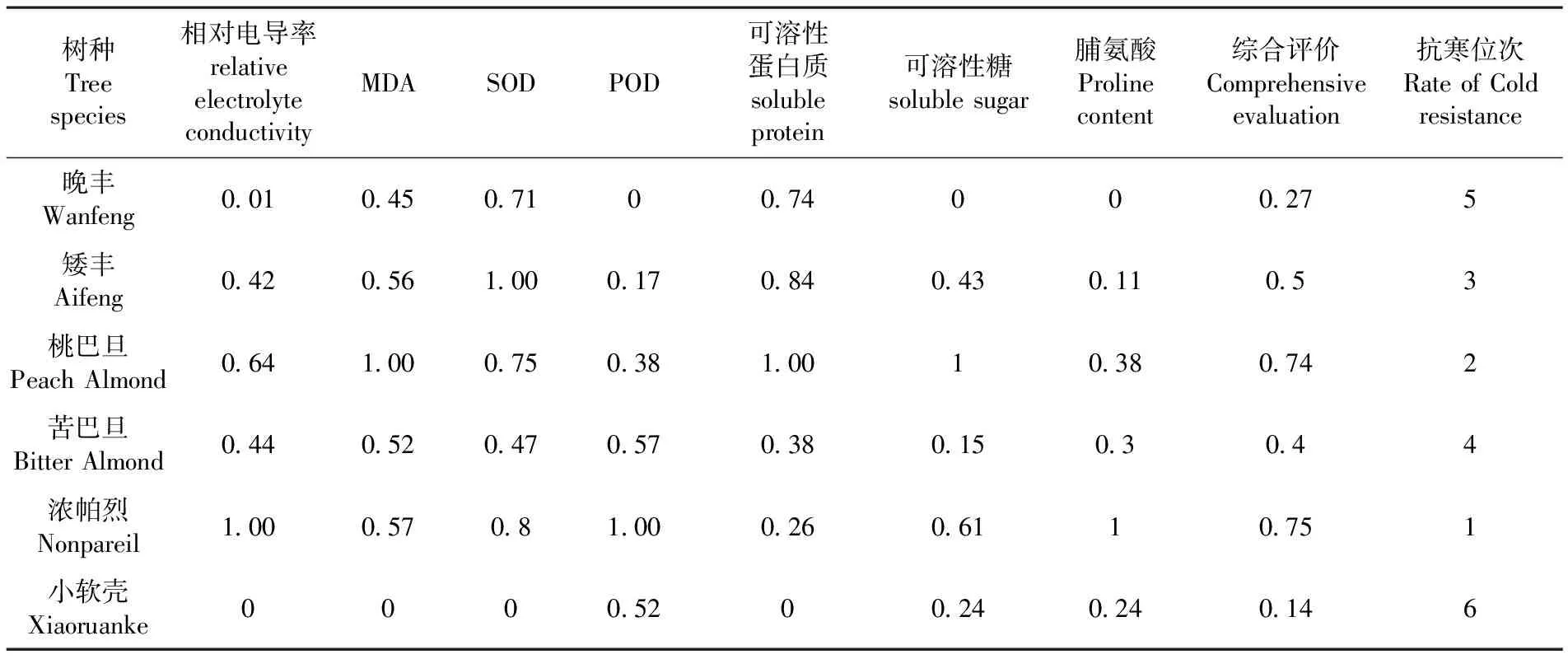
树种Treespecies相对电导率relativeelectrolyteconductivityMDASODPOD可溶性蛋白质solubleprotein可溶性糖solublesugar脯氨酸Prolinecontent综合评价Comprehensiveevaluation抗寒位次RateofColdresistance晚丰Wanfeng0010450710074000275矮丰Aifeng042056100017084043011053桃巴旦PeachAlmond06410007503810010380742苦巴旦BitterAlmond04405204705703801503044浓帕烈Nonpareil1000570810002606110751小软壳Xiaoruanke00005200240240146
3 讨 论
3.1 膜透性与越冬抗寒性关系
植物在低温条件下,细胞膜会发生膜脂的物相变化,膜脂从液品相变为凝胶相,导致膜结构被破坏,膜透性增大,膜内可溶性物质、电解质大量向膜外渗漏,破坏了细胞内外的离子平衡,因此电解质渗出率能直观的反应植物在低温胁迫下细胞膜透性变化。研究表明,扁桃枝条在越冬期间电导率与冬季气温变化呈显著性负相关关系,扁桃枝条在越冬期间随着温度的降低,对细胞伤害越大,导致膜透性增强渗透率升高,后期随温度的升高电导率降低,植物在越冬期间低温对扁桃枝条伤害性小,植物本身产生渗透调节物质和保护酶物质对细胞膜进行修护,电解质渗透率降低。这一结果与撒俊逸等[14],田景花等[15]研究结果一致。在低温胁迫下会加剧膜脂过氧化的作用,将会产生大量 MDA,它对质膜有毒害作用,它是细胞膜被破坏的标志性物质。试验表明,在越冬期间扁桃枝条中MAD含量变化与电导率变化一致,并与电导率呈显著性正相关关系,说明随着自然气温的降低加剧了膜脂过氧化作用,MDA含量增加,温度最低时,MDA含量最大。这一结果与王宁等[16],徐龙[17]研究结果一致。不同扁桃品种呈现不同程度的变化,其中矮丰,浓帕烈变化较为明显。植物在低温胁迫过程中会产生自由基,自由基的积累会导致细胞内活性氧的产生和清除平衡遭到破坏,从而引发或加剧膜脂过氧化作用,引起膜上蛋白质聚合和变性,降低了膜的流动性,膜的通透性增强,细胞膜受到了伤害。
3.2 保护酶与越冬抗寒性关系
植物在低温胁迫过程中会产生自由基,自由基的积累会导致细胞内活性氧的产生和清除平衡遭到破坏,从而引发或加剧膜脂过氧化作用,引起膜上蛋白质聚合和变性,降低了膜的流动性,膜的通透性增强,细胞膜受到了伤害。在植物细胞体内,植物自身产生了一套自我保护的抗氧化酶机制,SOD、POD活性是酶系统中重要的保护酶,SOD能将低温胁迫下产生的氧化物质H2O2还原成H2O和OH-,POD能够清除氧化还原产生OH-。试验研究表明,SOD活性、POD活性自然气温变化呈负相关关系,在越冬期间呈现先上升后下降的趋势。说明扁桃枝条在越冬期间温度的降低,导致于细胞内膜脂氧化作用,产生和大量的H2O2、OH-,细胞内产生了SOD,POD,将H2O2、OH-分解,减轻过氧化物质对细胞的伤害,之后随着温度的升高,过氧化物质减少,SOD、POD酶活性降低,这一结果与代汉萍等[18]、王倩等[19]的研究结果一致。但是浓帕烈枝条在越冬期间出现两个高峰值分别在12月21日和翌年的2月10日并在越冬最寒冷时期其含量降迅速下降,出现第一个高峰值可能是随着温度降低细胞内产生应激反应;随着温度降低POD活性下降但仍比入冬时期的活性高,随着环境温度的降低,植物体内活性氧增加, POD要分解活性氧生成OH-,持续增加的OH-而抑制POD酶活性,因此所出现的负反馈抑制[20],故在1月7日其活性下降;之后继续增加在2月10日出现第二个高峰值,可能是清除在低温环境中所产生的活性氧,因此POD活性增加,这一结果与何西凤等[21]的研究结果一致。
3.3 渗透调节物质与越冬抗寒性的关系
渗透调节是植物对逆境条件下的适应性,植物通过自身的防御系统来控制代谢,通过渗透调节物质来适应逆境带来的伤害。大量研究已证明,植物在低温条件下,植物细胞会失去大量的水分,植物的生长将会受到影响,但是植物在生长过程中会自身诱导产生一些渗透调节物质,以提高植物细胞液浓度,降低细胞的渗透势,植物就可以从外界吸收水分,维持正常的代谢生长。试验表明:自然越冬期间扁桃枝条中渗透调节物质可溶性蛋白质、可溶性糖、游离脯氨酸的含量与自然气温呈显著性负相关关系,其含量均随温度的变化呈先上升后下降的趋势。前期随着温度的降低,渗透调节物质的含量增加,可能是扁桃枝在越冬期间低温的胁迫,从而保证其安全越冬,后期其随温度升高含量降低。这一结果与李叶云等[22]、李鹏程等[23]、姚方等[24]研究结果一致。且可溶性蛋白质和可溶性糖在越冬期间的变化一致,且极显著相关。
4 结 论
植物越冬抗寒性生理的变化是一个复杂的过程,影响抗性大小的因素很多。主要以6个扁桃品种为试材,分别对6个扁桃品种在冬季越冬期间生理特性的研究,在冬季越冬电解质渗透率、MDA含量、SOD活性、POD活性、蛋白质含量、可溶性糖和脯氨酸与自然越冬温度均呈负相关关系,这些生理指标与植物冬季越冬抗寒性相关,并发现在1月温度最低的,对扁桃枝条伤害最大。采用这些生理指标可以对扁桃树抗寒性进行综合评价,也可用于扁桃树抗寒性的鉴定。不同品种间在冬季越冬期间生理变化存在显著性差异,不同扁桃品种抗寒性不同,利用采用模糊数学隶属函数的方法,综合评价6个扁桃品种冬季越冬抗寒性的大小,顺序为为浓帕烈>桃巴旦>矮丰>苦巴旦>晚丰>小软壳。
References)
[1] 兰彦平,吐拉克孜,郭文英,等. 巴旦杏的研究现状及开发利用前景[J]. 林业科学研究,2004,(5):674-679.
LAN Yan-ping, Tulakezi, GUO Wen-ying, et al. (2004). Research Progress and Utilization on Amydalus communis [J].ForestResearch, (5):674-679. (in Chinese)
[2]梅立新,刘文倩,魏钰,等. 中国扁桃资源与利用价值分析[J]. 西北林学院学报,2014,(1):69-72.
MEI Li-xin, LIU Wen-qian, WEI Yu, et al. (2014). Evaluation of the Resources and Development Potential of AmygdaLus spp. in China [J].JournalofNorthwestForestryUnrocrsity,(1):69-72. (in Chinese)
[3]武彦霞,王占和,何勇,等,田建保. 我国扁桃的生产现状及发展前景[J]. 山西农业科学,2005,(2):20-22.
WU Yan-xia, HE Yong, WANG Juan, et al. (2005). Present Status and Prospect of Almond Production in China [J].JournalofShanxiAgriculturalSciences, (2): 20-22. (in Chinese)
[4]张倩茹,尹蓉,王红宁,等. 扁桃种质资源及其开发利用研究进展[J]. 农产品加工,2016,(16):49-51.
ZHANG Qian-ru, YIN Rong, WANG Hong-ning, et al. (2016). Evaluation of the Resources and Development Potential ofAmygdaLus.spp. in China [J].AgriculturalProductsProcessing, (16): 49-51. (in Chinese)
[5]李疆,李文胜,成建红. 新疆扁桃生产的现状及发展对策[J]. 经济林研究,1998,(3):58-59.
LI Jiang, LI Wen-sheng, CHENG Jian-hong. (1998). The Present Situation and Development Countermeasures of Xinjiang Almond Production [J].EconomicForestResearches, (3):58-59. (in Chinese)
[6] 廖运建. 2014年新疆特色林果业统计表[J]. 新疆年鉴. 2015.
Liao Yunjian.In 2014 Statistics of Xinjiang Characteristic Forestry and Fruit Industry[J]. Xinjiang Yearbook, 2015. (in Chinese)
[7]李鹏,罗淑萍,田嘉,等. 低温冻害对扁桃花蕾抗寒机制的影响[J]. 经济林研究,2015,(2):20-25.
LI Peng, LUO Shu-ping, TIAN Jia, et al. (2015). Effects of frozen injury on freezing-tolerance mechanism of flower buds in almond [J].NonwoodForestResearch,(2):20-25. (in Chinese)
[8]孙士伟,张李华. 扁桃的开发价值及栽培技术[J]. 中国新技术新产品,2010,(10):228.
SUN Shi-wei, ZHANG Li-hua. (2015). The Development Value and Cultivation Technology of Almond [J].ChinaNewTechnology,NewProducts, (2):20-25. (in Chinese)
[9]王琳,王梦雪,李志军. 新疆莎车县5个扁桃品种抗寒性比较研究[J]. 塔里木大学学报,2014,(1):11-16.
WANG Lin, WANG Meng-xue, LI Zhi-jun. (2014). Comparative Study on Cold Resistance of Five Almond Cultivars of Xinjiang Shache County [J].JournalofTarimUniversity, (1):11-16. (in Chinese)
[10]杨晓静,徐崇志. 新疆扁桃F1群体抗寒性研究[J]. 塔里木大学学报,2012,(3):15-23.
YANG Xiao-jing, XU Chong-zhi. (2012). Study on Cold Resistance of Xinjiang Almond F1 Groups [J].JournalofTarimUniversity, (3):15-23. (in Chinese)
[11] 王江波,王合理,吴翠云,等. 十个引种红枣品种的抗寒性比较研究[J]. 北方园艺,2014,(12):18-20.
WANG Jiang-bo, WANG, WU Cui-yun, et al. (2014). Comparative Study on Cold Resistance of Ten Varieties of Introduced Ziziphus jujuba Mill. [J].NorthernHorticulture, (12):18-20. (in Chinese)
[12] 李合生.植物生理生化试验原理和技术[M].北京: 高等教育出版社,2004.
LI He-sheng. (2004).PrincipleandTechnologyofPlantPhysiologyandBiochemistryExperiment[M]. Beijing: Higher Education Press. (in Chinese)
[13] 陈新华. 甜樱桃不同品种抗寒性评价[D].保定:河北农业大学硕士学位论文,2009.
CHEN Xin-hua. (2009).TheEvaluationofColdResistanceinDifferentSweetCherryVarieties[D]. Master Thesis. Agricultural University of Hebei, Baoding. (in Chinese)
[14] 撒俊逸,白茹,孙志红,等. 自然越冬过程中库尔勒香梨抗寒性生理指标的变化[J]. 石河子大学学报(自然科学版),2016,(2):170-175.
SA Jun-yi, BAI Ru, SUN Zhi-hong, et al. (2016). Changes During Natural Overwintering in Korla Pear Cold Resistance Physiological Indexes [J].JournalofShiheziUniversity(NaturalScience) , (2): 170-175.(in Chinese)
[15] 田景花,王红霞,高仪,等. 核桃属植物休眠期的抗寒性鉴定[J]. 园艺学报,2013,(6):1 051-1 060.
TIAN Jing-hua, WANG Hong-xia, GAO Yi, et al. (2013). Walnut Plant Dormancy Period of Cold Resistance [J].ActaHorticulturaeSinica, (6):1,051-1,060. (in Chinese)
[16] 王宁,姚方,袁美丽,等. 自然降温过程中5种樟树的抗寒性分析[J]. 河南农业大学学报,2014,(3):302-309.
WANG Ning, YAO Fang, YUAN Mei-li, et al. (2014). Analysis on cold tolerance of five camphor tree species during the natural drop in temperature process [J].JournalofHenanAgriculturalUniversity,(3):302-309. (in Chinese)
[17] 徐龙. 不同种源酸枣实生苗抗寒性比较[D].阿拉尔:塔里木大学硕士学位论文,2013.
XU Long. (2013).ComparisononColdResistanceofDifferentProvenancesZizibusjujubavar.spinosa(Bunge)HuSeedlings[D]. Master Thesis. Tarim University, Alar. (in Chinese)
[18] 代汉萍,薛志杰,郭修武. 树莓自然越冬枝条生理生化特性研究[J]. 吉林农业大学学报,2009,(5):573-577.
DAI Han-ping, XUE Zhi-jie, GUO Xiu-wu. (2009). Studies on Physiological and Biochemical Characteristics of Raspberry Canes under Natural Overwintering Condition [J].JournalofJilinAgriculturalUniversity, (5): 573-577. (in Chinese)
[19] 王倩,冷平生,关雪莲,等. 九种景天植物在越冬期间生理生化指标的变化[J]. 北方园艺,2010,(19):114-117.
WANG Qian, LENG Ping-sheng, GUAN Xue-lian, et al. (2010). Changes of Physiological and Biochemistry Characteristics 9 Varieties ofSeduamduring Autumn and Winter [J].Northernhorticulture, (19): 114-117. (in Chinese)
[20] Wang, S. Y., Faust, M., & Jiao, H. J. (1991). Changes in superoxide dismutase activity during thidiazuron-induced lateral budbreak of apple.HortscienceAPublicationoftheAmericanSocietyforHorticulturalScience, 26(9): 1,202-1,204.
[21]何西凤,杨途熙,魏安智,等. 自然越冬过程中花椒抗寒性生理指标的变化[J]. 东北林业大学学报,2009,(5):67-69.
HE Xi-feng, YANG Tu-xi, WEI An-zhi, et al. (2009).Changes of Physiological Indexes ofZanthoxylumbungeanumRelated to Cold Resistance during Natural Overwintering [J].JournalofNortheastForestryUniversity, (5): 67-69. (in Chinese)
[22]李叶云,舒锡婷,周月琴,等. 自然越冬过程中3个茶树品种的生理特性变化及抗寒性评价[J]. 植物资源与环境学报,2014,(3):52-58.
LI Ye-yun, SHU Xi-ting, ZHOU Yue-qin, et al. (2014).Change and Evaluation of Cold Resistance Physiological Characteristics of 3 Tea Cultivars During Natural Overwintering [J].JournalofPlantResourcesandEnvironment, (3):52-58. (in Chinese)
[23]李鹏程,郭绍杰,李铭,等. 自然越冬过程中山葡萄抗寒生理指标的变化[J]. 贵州农业科学,2013,(1):69-71.
LI Peng-cheng, GUO Shao-jie, LI Ming, et al. (2013). Change of Cold-resistant Physiological Indicators ofVitisamurensisduring Natural Overwintering [J].GuizouAgriculturalSciences,(1):69-71. (in Chinese)
[24]姚方,吴国新,梅海军. 自然降温过程中3种樟树渗透调节物质的动态变化[J]. 华南农业大学学报,2012,(3):378-383.
YAO Fang, WU Guo-xin, MEI Hai-jun. (2012). Dynamic Changes in Osmotic Adjustment Substance of Three Camphor Species Under Natural Cooling [J].JournalofSouthChinaAgriculturalUniversity,(3):378-383. (in Chinese)
ChangesofPhysiologicalIndexesandComprehensiveEvaluationofColdResistanceofAlmondduringNaturalOverwintering
OU Huan1,2, WANG Xu-chun1,2, WANG Zhen-lei1,2,3, XU Chong-zhi1,2,3,WANG Xin-jian1,2,3, LIN Min-juan1,2,3
(1.CollegeofPlantScience,TarimUniversity/EngineeringLaboratoryofCharacteristicFruitTreeProductioninSouthernXinjiang,ProductionandConstructionCorps,AlarXinjiang843300,China;2.TheNationalandLocalJointEngineeringLaboratoryofHighEfficiencyandSuperior-QualityCultivationandFruitDeepProcessingTechnologyofCharacteristicFruitTreesinSouthXinjiang,AlarXinjiang843300,China; 3.KeyLaboratoryofBiologicalResourcesProtectionandUtilizationinTarimBasin,XinjiangProductionandConstructionCorps,Tarim,AlarXinjiang843300,China)
ObjectiveIn this paper, the physiological characteristics of 6 almond varieties during their natural wintering will be evaluated, and the cold resistance of different almond branches will be comprehensively evaluated, which might provide a theoretical basis for introduction, cultivation and promotion of almond.MethodThe membrane permeability, protective enzyme activity and osmotic adjustment substance content of each almond were determined during the overwintering process. The changes and correlations of physiological indexes during winter were analyzed, and the differences among different cultivars were compared.ResultIn the process of natural overwintering, the temperature of natural winter was negatively correlated with electrolyte permeability, MDA content, SOD activity, POD activity, soluble protein content, soluble sugar content and proline content, and all increased first and then decreased. The change of temperature had a great effect on the change of electrolyte permeability, MDA content, SOD activity, soluble protein content and soluble sugar content, but had little effect on POD activity and proline content. And The varieties with strong cold resistance were more abundant than those with weak cold resistance, among which the changes of physiological indexes were more obvious during the overwintering period.ConclusionThe results showed that through the physiological indexes of almond branches during overwintering that were determined, the conductivity, SOD, soluble sugar and protein could be used as indexes to evaluate the cold resistance, and the relationship between the cold resistance of the six almond varieties was evaluated by combining the membership function. The cold resistance of 6 almond cultivars was in turn: Nonpareil > Peach Almond > Aifeng > Bitter Almond > Wanfeng > Xiaoruanke.
almond; natural overwintering; cold resistance; physiological index; comprehensive evaluation
Lin Min-juan (1979 -),Female, Native place: Hebei .Research direction: fruit germplasm resources, fruit quality and efficient cultivation physiology.(E-mail)lmjzky@163.com
S662.9
A
1001-4330(2017)10-1785-11
10.6048/j.issn.1001-4330.2017.10.003
2017-07-06
南疆特色林果科技成果转化示范基地建设(TDZX201614);巴旦杏高效生产关键技术集成与规模化示范(子课题5:2013BAD14B02)
欧欢(1992-),女,四川人,硕士研究生,研究方向为果树栽培生理与生态,(E-mail)547939605@qq.com
林敏娟(1979-),女,河北人,副教授,研究方向为果树种质资源、果树优质高效栽培生理,(E-mail)lmjzky@163.com
Supported by: Construction of demonstration base for scientific and technological achievements transformation of featured trees in southern Xinjiang(TDZX201614)and Almond efficient production key technology integration and large-scale demonstration (Sub-project 5: 2013BAD14B02)
猜你喜欢
当代水产(2022年7期)2022-09-20
热带作物学报(2022年5期)2022-06-01
黑龙江粮食(2021年10期)2021-12-13
中国果业信息(2021年7期)2021-12-01
今日农业(2021年21期)2021-11-26
中成药(2018年6期)2018-07-11
现代园艺(2017年11期)2017-06-28
中国农业文摘-农业工程(2016年5期)2016-04-12
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
