
AHLs对小球藻PSⅡ光化学活性与生化指标的影响研究*
2017-10-17 08:08唐学玺张文慧姜智飞高金伟周文礼
中国海洋大学学报(自然科学版) 2017年11期
窦 勇, 甘 雨, 唐学玺, 张文慧, 姜智飞, 高金伟, 周文礼**
(1.天津农学院水产学院,天津市水产生态及养殖重点实验室,天津 300384; 2.中国海洋大学海洋生命学院,山东 青岛 266003)
AHLs对小球藻PSⅡ光化学活性与生化指标的影响研究*
窦 勇1, 甘 雨1, 唐学玺2, 张文慧1, 姜智飞1, 高金伟1, 周文礼1**
(1.天津农学院水产学院,天津市水产生态及养殖重点实验室,天津 300384; 2.中国海洋大学海洋生命学院,山东 青岛 266003)
小球藻是一种重要的资源微藻,在水产养殖、生物能源和功能型保健食品等领域都有广泛的应用。小球藻藻际共栖细菌产生的群感信号分子N-酰基-高丝氨酸内酯类化合物(N-acyl-homoserine lactones,AHLs)不仅可以调控细菌的生物学功能,还能影响藻类的生命活动,常会引起小球藻高密度培养系统崩溃,因此研究AHLs对小球藻生理代谢的影响和机理具有重要的现实意义。本研究采用批次培养的方法,研究了100、200、400 mmol/L C10-HSL(N-癸酰-L-高丝氨酸内酯)对普通海水小球藻(Chlorellavulgaris)PSⅡ光化学活性与生化指标(SOD、CAT、GPx和Mg2+-ATPase)的影响。结果显示:在C10-HSL作用下,小球藻PSⅡ光化学活性指标——最大光能转化效率Fv/Fm、实际光能转化效率Yeild及表观光合电子传递效率ETR均明显下降,而且C10-HSL对Yeild的影响强于对Fv/Fm的作用;小球藻SOD、CAT、GPx和Mg2+-ATPase活性均大致呈现先升高后降低的变化趋势,但峰值出现时间不同,C10-HSL对生化指标的影响均未表现随时间规律变化的剂量-效应关系。本研究阐明了AHLs对小球藻光合生理、抗氧化酶系统和能量代谢的影响规律,暗示AHLs引发的氧化损伤以及能量代谢紊乱可能是造成小球藻培养体系崩溃的重要原因,实验结果为小球藻无菌化健康培养与养殖体系优化提供了一定的依据。
AHLs;海水普通小球藻;光化学活性;生化指标
小球藻(Chlorella)是绿藻门的单细胞微藻,细胞内氨基酸、维生素、多糖等含量丰富,它在水产养殖、生物能源生产和功能型保健食品开发领域有广泛的应用。在自然水体和室内高密度培养体系中,藻际环境常存在着大量共栖细菌,这些细菌会产生一系列次级代谢产物,这些物质可以启动细菌密度依赖的基因表达,使细菌产生独特而多样的生物学功能,这一现象称为细菌的群感效应(Quorum Sensing, QS),而介导群感效应的细菌代谢产物称为群感信号物质[3],研究证实微生物的生物发光、聚集生长、胞外多糖的合成与分泌、生物膜的形成都受到群感信号分子的调控[4-7]。群感信号分子主要分为3类,其中N-酰基-高丝氨酸内酯类化合物(N-acyl-homoserine lactones,AHLs)是革兰氏阴性菌群感系统中最常见的信号物质,AHLs家族成员众多,其共同点是分子上都含有1个高丝氨酸内酯环,不同的AHLs分子含有不同的酰化支链且具有不同的生物活性[8],而目前的研究多集中于N-己酰-L-高丝氨酸内酯(C6-HSL)、N-酮己酰-L-高丝氨酸内酯(OHHL)、N-辛酰-L-高丝氨酸内酯(C8-HSL)、N-癸酰-L-高丝氨酸内酯(C10-HSL)这几种分子上[9-14]。
近年来的研究表明群感信号分子不仅能对细菌自身的生物学功能进行反馈调节,而且可以介导藻类-微生物的种间相互作用,徐鲁燕等[15]发现某些海洋细菌产生的C6-HSL和C8-HSL显著抑制了东海原甲藻、赤潮异弯藻生长;毕相东等[16]指出藻际异养菌产生的C6-HSL会诱导小球藻抗氧化酶系统应激表达,而另有学者[17-18]证实微藻能通过降解AHLs分子或合成AHLs类似物来干扰共栖细菌的胞间通讯。以往的工作多侧重于AHLs某一方面的作用而缺乏系统性,本研究综合考察了C10-HSL对C.vulgarisPSⅡ光化学活性、氧化应激和能量代谢的影响,探讨了C.vulgaris对AHLs胁迫的响应机制,旨在为小球藻无菌化健康培养与养殖体系优化提供依据。
1 材料与方法
1.1 藻种来源与培养条件
实验用小球藻由中国海洋大学藻种室提供,采用f/2培养液培养,培养温度(23±1)℃,光暗比12 h∶12 h,光强为60 μmol·m-2·s-1。培养期间每天摇瓶6次,防止微藻细胞附壁下沉。选取处于对数生长期的小球藻细胞进行实验。
1.2 药品与试剂
N-癸酰-L-高丝氨酸内酯(C10-HSL)标准品购自sigma公司,原药溶于无水乙醇配制成1 mol/L的母液,于4℃保存,使用时根据实际情况加入无水乙醇稀释配成具有浓度梯度的工作液,然后加入到不同处理组的小球藻培养液中,对照组(CK)藻液中仅加入无水乙醇。
1.3 实验体系构建和分析指标系统
根据前期预实验结果,将配制好的C10-HSL母液加入到处于对数生长期的100 mL小球藻培养液中,使其在培养液中的最终浓度为100、200、400 mmol/L,对照组按照1.2设置。每个实验组设置3个平行,实验重复3次。在实验开始的0、1、2、3、4、5、6、7 d测定小球藻PSⅡ光化学活性——PSⅡ最大光能转化效率(Fv/Fm)、PSⅡ的实际光能转化效率(Yeild)、表观光合电子传递效率(ETR);生化指标——超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)、Mg2+-ATP酶(Mg2+-ATPase)。
1.4 测定方法
1.4.1 小球藻PSⅡ光化学活性测定 使用德国Walz公司生产的IMAGING-PAM调制脉冲荧光仪测定PSⅡ光化学活性。向比色杯中依次加入3 mL蒸馏水和15 μL藻液,混匀,将样品暗适应15 min,读取Fv/Fm、Yeild和ETR数值。
1.4.2 小球藻生化指标测定 每次取样时量取各处理组藻液40 mL在4℃下以6 000 r/min离心10 min,取沉淀藻泥加入3 mL生理盐水,冰浴研磨,4 ℃下3 000 r/min离心10 min,上清液用于生化指标测定。将SOD对超氧化物阴离子反应抑制率达到50%时定义为一个SOD酶活力单位,将每秒钟每毫克组织蛋白分解1 μmol H2O2的量定义为一个CAT酶活力单位,将每毫克组织蛋白每分钟催化1 μmol NADPH转变为NADP定义为一个GPx单位,将1 h内每毫克组织蛋白中ATP酶通过分解作用而产生1 μmol无机磷的量定义为一个Mg2+-ATP酶活力单位,以上四种酶活单位均为U/mgprot。所有酶活分析均采用比色法,吸光度值使用UV-1240紫外-可见分光光度计测定。
1.5 数据处理与分析
所有实验数据均以平均值±标准误(mean±SE,n=3)表示。使用SPSS 17.0软件进行单因素方差one-way ANOVA分析,并采用LSD方法进行多重比较,显著性水平α=0.05,标记*的处理组表示与对照组差异显著(P<0.05),反之差异不显著(P>0.05)。
2 结果与分析
2.1 C10-HSL对小球藻PSⅡ光化学活性的影响
2.1.1 C10-HSL对小球藻Fv/Fm的影响Fv/Fm为最大量子产量,反映了植物PSII潜在最大光合能力,而环境条件改变时微藻Fv/Fm会发生显著变化[19-20]。如图1所示,随着C10-HSL处理时间的延长小球藻Fv/Fm总体呈现下降趋势,实验前5天变化并不明显,而从第6天开始显著降低,至实验结束时Fv/Fm达到最小值,此时100、200和400 mmol/L处理组Fv/Fm分别为0.41、0.49、0.52,较初始状态分别减小36.92%、27.94%和21.21%。C10-HSL对小球藻Fv/Fm的影响随时间变化表现不同的剂量-效应关系,实验前2天小球藻Fv/Fm随C10-HSL浓度的增加而降低,从第3天开始Fv/Fm随C10-HSL浓度的增加而升高。

图1 C10-HSL对小球藻Fv/Fm的影响
2.1.2 C10-HSL对小球藻Yeild的影响 Yeild为实际光化学量子产量,反映植物PSII在部分关闭情况下的实际原初光能捕获效率。实验期间小球藻Yeild波动较大无明显规律(见图2),前2天Yeild缓慢降低,第3~5天维持相对稳定,第6天显著下降,而在第7天有所回升。C10-HSL对小球藻Yeild的影响随时间变化表现不同的剂量-效应关系,实验前2天微藻Yeild随C10-HSL浓度的增加而降低,第3~5天 Yeild随C10-HSL浓度的增加而升高,尤其是200和400 mmol/L处理组小球藻Yeild显著高于对照(P<0.05),而在实验最后2天100和400 mmol/L处理组Yeild高于200 mmol/L处理组。
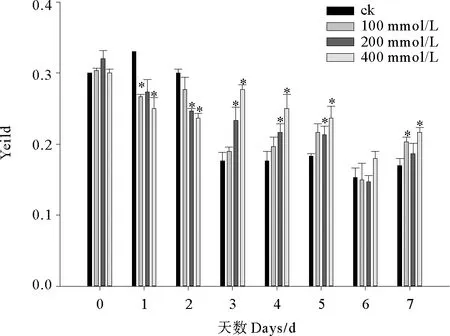
图2 C10-HSL对小球藻Yeild的影响
2.1.3 C10-HSL对小球藻ETR的影响 ETR为表观光合电子传递速率,是反映植物PSII反应中心活性与光合电子传递效率强弱的一个重要指标,其数值与光强、植物吸收入射光和能量分布比例以及光子通量密度有关[21]。小球藻ETR变化规律与Yeild大体一致,也随时间呈“下降—稳定—下降—回升”的波动趋势(见图3)。C10-HSL对小球藻ETR的影响随时间表现出不同的剂量-效应关系,实验初期C10-HSL处理组小球藻ETR均显著低于对照组(P<0.05),从第3天开始AHL对微藻ETR的刺激作用显现,C10-HSL处理组的光合电子传递速率逐渐高于对照。实验结束时100、200和400 mmol/L处理组小球藻ETR较初始状态分别下降了 33.33%、40.91%和28.57%。
2.2 C10-HSL对小球藻生化指标的影响
2.2.1 C10-HSL对小球藻SOD活性的影响 实验期间小球藻SOD活性呈先升高后降低的变化趋势(见图4)。实验刚开始SOD就被大量诱导,从第3天起C10-HSL处理组微藻SOD活性均显著高于对照组(P<0.05),至第5天 100、200和400 mmol/L处理组酶活平均水平达到峰值49.37、55.03和61.67 U/mgprot,较初始阶段分别上升了56.39%、76.95%和99.78%,但从第6天开始酶活力迅速回落,至实验结束时各处理组小球藻SOD活性接近第2~3天的水平。C10-HSL对小球藻SOD的影响未表现出随时间规律变化的剂量-效应关系,实验第1、3、5天微藻SOD活力随C10-HSL浓度增加而上升,实验第7天酶活随C10-HSL浓度增加而降低。

图3 C10-HSL对小球藻ETR的影响
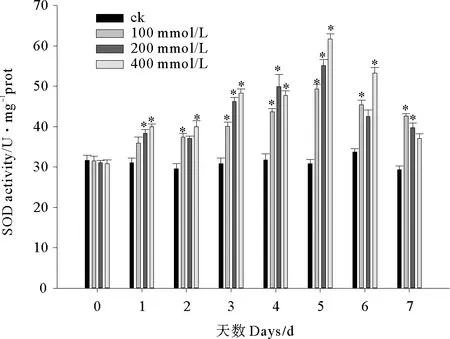
图4 C10-HSL对小球藻SOD活力的影响
2.2.2 C10-HSL对小球藻CAT活性的影响 小球藻CAT活性的变化与SOD大体一致,也表现出先升高后降低的趋势,但在数值上略低于SOD(见图5)。实验开始微藻CAT活性便逐渐上升,从第3天起C10-HSL处理组酶活力均显著高于对照组(P<0.05),至第5天 100、200和400 mmol/L处理组酶活平均水平达到峰值41.27、49.70和53.27 U/mg prot,较初始阶段分别上升了45.48%、72.98%和78.95%,但从第6天起酶活力迅速下降,至实验结束时各C10-HSL处理组CAT活性平均值仅为峰值水平的86.83%、64.99%和61.95%。实验第1、3、5天微藻CAT活性随C10-HSL浓度增加而上升,其他时间C10-HSL对CAT的影响未表现出浓度依赖的剂量-效应关系。
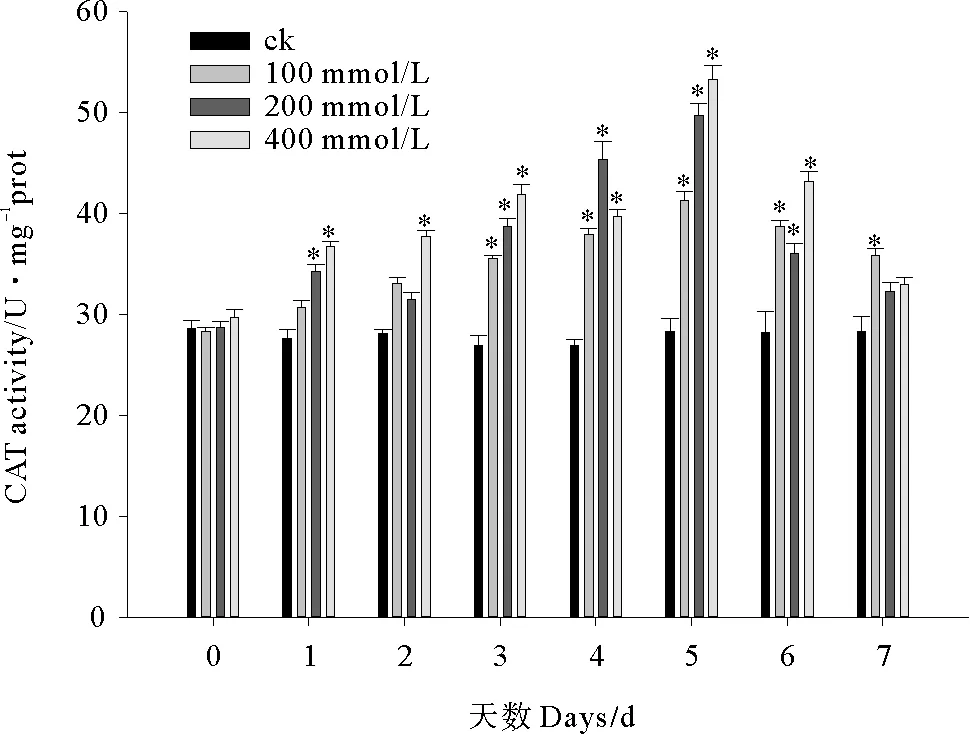
图5 C10-HSL对小球藻CAT活力的影响
2.2.3 C10-HSL对小球藻GPx活性的影响 实验期间小球藻GPx活性呈先升高后降低的变化趋势(见图6)。实验第1~4天,各C10-HSL处理组微藻GPx活性逐渐上升,至第4d时达到峰值,100、200和400 mmol/L处理组酶活平均水平分别为38.50、35.93和45.17 U/mgprot,从第5天起各处理组酶活力逐渐下降,至实验结束时微藻GPx活性接近初期水平。实验第1、3天小球藻GPx活性随C10-HSL浓度增加而上升,实验第6、7天酶活随C10-HSL浓度增加而降低,其他时间C10-HSL对GPx的影响未表现出浓度依赖的剂量-效应关系。
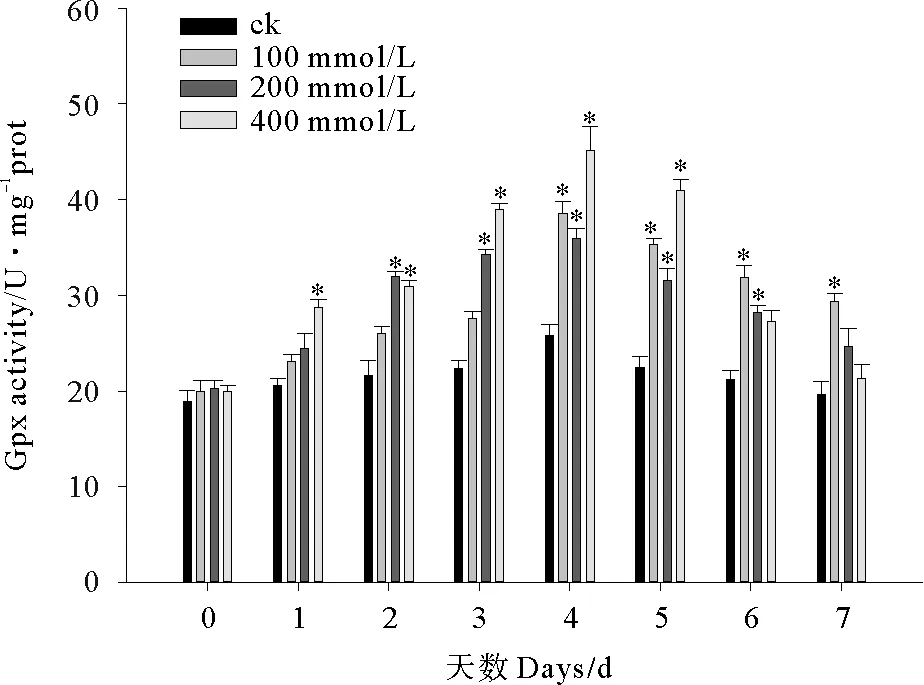
图6 C10-HSL对小球藻GPx活力的影响
2.2.4 C10-HSL对小球藻Mg2+-ATPase活性的影响 在C10-HSL作用下,小球藻Mg2+-ATPase活性表现出先升高后下降的变化规律(见图7)。实验初期Mg2+-ATPase活力便迅速上升,至第3天达到峰值,此时100、200和400 mmol/L处理组酶活平均水平分别为23.67、21.33和24.60 U/mgprot,从第4天起各处理组酶活力逐渐下降,至实验结束时微藻GPx活性接近初始水平。C10-HSL对小球藻Mg2+-ATPase活性的影响未表现出随时间变化的浓度依赖型特征,在第3天酶活力随C10-HSL浓度增加而上升,实验第6天酶活随C10-HSL浓度增加而降低。

图7 C10-HSL对小球藻Mg2+-ATPase活力的影响
3 讨论
有文献[22]指出,在正常状态下藻类的最大量子产量Fv/Fm值稳定在0.65左右,当受到环境胁迫时该值会显著下降,而在本研究中小球藻的Fv/Fm值处于0.41~0.63之间,表明C10-HSL对小球藻产生了较强的胁迫作用,在一定程度上抑制了微藻PSII的光化学活性。另外本实验结果证实在C10-HSL胁迫下,小球藻Yeild的下降幅度超过Fv/Fm,暗示C10-HSL对微藻实际光合能力的影响强于对其最大光合潜力的作用。
SOD、CAT、GPx均为抗氧化酶,是生物体清除活性氧保护细胞免受氧化损伤的重要屏障[23]。在本研究中小球藻细胞内3种抗氧化酶均被C10-HSL诱导表达,说明C10-HSL对微藻细胞施加了一定程度的氧化胁迫。Mg2+-ATPase是调节细胞能量代谢和离子转运的关键酶,是维持生物体正常生理功能的重要物质[24]。本实验中小球藻Mg2+-ATPase活性波动剧烈,表明C10-HSL干扰了微藻细胞固有的ATP合成和离子运输过程。由此可以推测,AHLs引发的氧化损伤以及能量代谢紊乱可能是共栖细菌造成小球藻高密度培养体系崩溃的重要原因。本研究中C10-HSL对SOD、CAT、GPx和Mg2+-ATPase的作用均为先诱导后抑制,这与其他学者[16]的结论有所不同,造成此种差异的原因可能与实验中不同的AHLs分子结构以及胁迫浓度有关。
值得注意的是,AHLs家族的不同分子及处理浓度对其生物活性的发挥存在很大差异,毕相东[25]发现在400 nmol/L的C6-HSL胁迫下小球藻PSII的D1蛋白表达量显著下调,而烯醇酶的表达量显著上调,表明C6-HSL在损伤微藻细胞光合系统的同时加强了其厌氧呼吸速率,周丽娟等[26]证实10~80 μmol/L的α-氨基-γ-丁内酯氢溴酸(AHLs的一种)可以明显抑制铜绿微囊藻(Microcystisaeruginosa)和集胞藻(Synechocystisp.)的生物量及蛋白表达,以上结果说明AHLs的生物活性取决于其分子结构、作用浓度及作用对象,因此摸索不同AHLs对资源微藻的生物活性差异有助于优化微藻无菌化培养系统,同时也可为微藻的集约化健康培养提供参考。
有研究[8]表明AHLs介导的LuxI/LuxR双元件系统是革兰氏阴性菌群感信号途径的物质基础,该系统调控着微生物的种群动态和代谢生理,其中AHLs发挥了类似转录因子的作用,但目前AHLs影响藻类生理的机制尚不明晰,AHLs分子是否同样作为转录因子类似物影响藻类相关基因的表达进而影响其生理代谢的强度和途径的疑问至今未得到圆满回答,所以在基因转录和表达水平上探讨AHLs对藻类生理过程的作用机制应该作为未来研究的重要方向。
4 结论
(1)在100、200和400 mmol/L C10-HSL作用下,小球藻PSⅡ光化学活性指标——最大光能转化效率Fv/Fm、实际光能转化效率Yeild及表观光合电子传递效率ETR均明显下降,而且C10-HSL对Yeild的影响强于对Fv/Fm的作用;
(2)在C10-HSL作用下,小球藻SOD、CAT、GPx和Mg2+-ATPase活性均大致呈现先升高后降低的变化趋势,但峰值出现时间不同,C10-HSL对生化指标的影响均未表现随时间规律变化的剂量-效应关系,浓度依赖型特征不突出。
[1] 刘加慧, 杨洪帅, 王辉. 温度、盐度和pH对小球藻生长率的联合效应[J]. 水生生物学报, 2014, 38(3): 446-453. Liu J H, Yang H S, Wang H. Combined effects of temperature, salinity and pH on the specific growth rate ofChlorella[J]. Acta Hydrobiologica Sinica, 2014, 38(3): 446-453.
[2] 梁英, 黄徐林, 田传远, 等. 海洋药源微藻研究进展[J]. 中国海洋大学学报(自然科学版), 2016, 46(11): 32-43. Liang Y, Huang X L, Tian C Y, et al. Advances in studies on marine medical microalgae[J]. Periodical of Ocean University of China, 2016, 46(11): 32-43.
[3] Fuqua W C, Winans S C, Greenberg E P. Quorum sensing in bacteria: the LuxR-LuxI family of cell density responsive transcriptional regulators[J]. Journal of Bacteriology, 1994, 176(2): 269-275.
[4] Paerl H W, Pinckney J L. A mini-review of microbial consortia: Their roles in aquatic production and biogeochemical cycling[J]. Microbiology Ecology, 1996, 31(3): 225-247.
[5] Ianora A, Boersma M, Casotti R, et al. New trends in marine chemical ecology[J]. Estuaries and Coasts, 2006, 29(4): 531-551.
[6] Qian Y P, Lau C S, Dahms U H, et al. Marine biofilms as mediators of colonization by marine macroorganisms: Implications for antifouling and aquaculture[J]. Marine Biotechnology, 2007, 9(4): 399-410.
[7] Merchant S S, Prochnik E S, Vallon O, et al. TheChlamydomonasgenome reveals the evolution of key animal and plant functions[J]. Science, 2007, 318(5848): 245-251.
[8] Miller M, Bassler B. Quorum sensing in bacteria[J]. Annual Review of Microbiology, 2001, 55(3): 165-199.
[9] Liu B H, Koh P K, Kim S J, et al. The effects of betonicine, floridoside, and isethionic acid from the red algaAhnfeltiopsisflabelliformison quorum-sensing activity[J]. Biotechnology and Bioprocess Engineering, 2008, 13(4): 458-463.
[10] 徐鲁燕. 海洋微藻共栖细菌抑藻活性及群体感应相关基因的初步研究[D]. 青岛: 国家海洋局第一海洋研究所, 2013. Xu L Y. Preliminary Study on the Algicidal Activity of Commensalism Bacteria Associated with Marine Microalgae and Genes Related with Quorum-sensing Effects[D]. Qingdao: The First Institute of Oceanography, SOA, 2013.
[11] Priha O, Virkajarvi V, Juvonen R, et al. Quorum sensing signalling and biofilm formation of brewery-derived bacteria, and inhibition of signaling by natural compounds[J]. Current Microbiology, 2014, 69(5): 617-627.
[12] 侯保连, 李安婕, 孙趣. AHLs群体感应信号分子对硝化污泥附着生长及硝化效果的影响[J]. 环境科学学报, 2015, 35(9): 2773-2779. Hou B L, Li A J, Sun Q. The effect of N-acy-homoserine lactones mediated quorum-sensing on the adhesion growth and nitrification of nitrifying sludge[J]. Acta Scientiae Circumstantiae, 2015, 35(9): 2773-2779.
[13] Pande S G, Natrah M F, Flandez V A, et al. Isolation of AHL-degrading bacteria from micro-algal cultures and their impact on algal growth and on virulence ofVibriocampbelliito prawn larvae[J]. Applied Microbiology and Biotechnology, 2015, 99(24): 10805-10813.
[14] Teplitski M, Chen H C, Rajamani S, et al.Chlamydomonasreinhardtiisecretes compounds that mimic bacterial signals and interfere with quorum sensing regulation in bacteria[J]. Plant Physiology, 2004, 134(1): 137-146.
[15] 徐鲁燕, 郑立, 韩笑天, 等. 具有群体感应系统微藻共栖细菌的筛选及抑藻活性的研究[J]. 海洋与湖沼, 2012, 43(6): 1149-1155. Xu L Y, Zheng L, Han X T, et al. Screening of microalgae associated bacteria with quorum sensing system and their algicidal activity[J]. Oceanologia et Limnologia Sinica, 2012, 43(6): 1149-1155.
[16] 毕相东, 周文礼, 邢克智, 等. AHLs信号分子对小球藻生长及抗氧化酶系统的影响[J]. 海洋环境科学, 2012, 31(6): 897-900. Bi X D, Zhou W L, Xing K Z, et al. Effects of AHLs (N-hexanoyl-DL-homoserine lactone, C6-HSL) on growth and antioxidant defense system ofChlorellavulgaris[J]. Marine Environmental Science, 2012, 31(6): 897-900.
[17] Natrah M F, Defoirdt T, Sorgeloos P, et al. Disruption of bacterial cell-to-cell communication by marine organisms and its relevance to aquaculture[J]. Marine Biotechnology, 2011, 13(2): 109-126.
[18] Wietz M, Duncan K, Patin V N, et al. Antagonistic interactions mediated by marine bacteria: The role of small molecules[J]. Journal of Chemical Ecology, 2013, 39(7): 879-891.
[19] 张虎, 张桂艳, 温小斌, 等. pH对小球藻Chlorellasp. XQ-200419光合作用、生长和产油的影响[J]. 水生生物学报, 2014, 38(6): 1084-1091. Zhang H, Zhang G Y, Wen X B, et al. Effect of pH on the photosynthesis, growth and lipid production ofChlorellasp. XQ-200419[J]. Acta Hydrobiologica Sinica, 2014, 38(6): 1084-1091.
[20] 文欣, 刘宝宁, 周成旭, 等. 颗石藻Pleurochrysiscarterae抗捕食特征[J]. 生态学报, 2016, 36(2): 525-534. Wen X, Liu B N, Zhou C X, et al. Characterization of anti-predator abilities ofPleurochrysiscarterae[J]. Acta Ecologica Sinica, 2016, 36(2): 525-534.
[21] 徐兴莲, 宋熙坤, 岳瑞, 等. 氮限制对硅藻三角褐指藻光系统Ⅱ光化学反应的影响[J]. 生态学杂志, 2016, 35(1): 183-188. Xu X L, Song X K, Yue R, et al. Effects of nitrogen limitation on photosystem Ⅱ photochemistry reaction in diatomPhaeodactylumtricornutum[J]. Chinese Journal of Ecology, 2016, 35(1): 183-188.
[22] 刘凯. 固定化小球藻的原位光合作用及其对废水中无机氮磷的影响[D]. 天津: 天津科技大学, 2012. Liu K. In Situ Photosynthesis of ImmobilizedChlorellasorokinianaGxnn01 and Removal of Nitrogen and Phosphorus from Waste Water[D]. Tianjin: Tianjin University of Science & Technology, 2012.
[23] 窦勇, 乔秀亭, 陈丽梅, 等. 萘暴露对环文蛤的氧化胁迫与损伤研究[J]. 南方水产科学, 2014, 10(4): 39-44. Dou Y, Qiao X T, Chen L M, et al. Study on oxidative stress and damage ofCyclinasinensisexposed to naphthalene[J]. South China Fisheries Science, 2014, 10(4): 39-44.
[24] Wang G X, Roux B, Feng F, et al. The decoy substrate of a pathogen effect and a pseudokinase specify pathogen-induced modified-self recognition and immunity in plants[J]. Cell Host & Microbe, 2015, 18(3): 285-295.
[25] 毕相东. 小球藻与优势共栖异养菌间的相互作用及其对细菌群体感应信号分子的响应[D]. 青岛: 中国海洋大学, 2013. Bi X D. Ecological Effects BetweenChlorellavulgarisand Dominant Associated Heterotrophic Bacteria and Response to Quorum Sensing Signal Molecules[D]. Qingdao: Ocean University of China, 2013.
[26] 周丽娟, 陈小兰, 王波, 等. 外源AHLs信号物质对蓝藻生长代谢的影响[J]. 云南大学学报(自然科学版), 2007, 29(3): 303-307. Zhou L J, Chen X L, Wang B, et al. The effects of exogenous AHLs on the growth and metabolism of blue-green algae[J]. Journal of Yunnan University, 2007, 29(3): 303-307.
Abstract: As an excellent resource microalgae,Chlorellahas wide applications for aquaculture, production of biofuel and development of functional food. AHLs(N-acyl-homoserine lactones)are known as the quorum sensing signal molecule produced by commensalism bacteria in phycosphere. AHLs are playing important roles in not only regulating the biological function of bacteria, but also affecting the life activities of algae. Therefore, AHLs could result in the high-density culture system ofChlorellacollapse. A laboratory study was conducted to evaluate the effects of 100, 200, 400 mmol/LC10-HSL (N-decanoyl-L-homoserine lactones) on PSⅡ photochemistry activity, antioxidase(SOD, CAT, GPx) activities and energy metabolism enzyme Mg2+-ATPase activity of the marine microalgaeChlorellavulgaris, in the photoautotrophic culture process. The results were as follows: PSⅡ photochemistry activity indicators—Fv/Fm(maximal photochemical efficiency), Yeild (actual photochemical efficiency) and ETR (apparent photosynthetic electron transport ratio) decreased obviously under the effect of C10-HSL. C10-HSL exerted higher effect on Yeild thanFv/Fm. There was a same fluctuation trend of the enzymes activities of SOD, CAT,GPx and Mg2+-ATPase which increased first and then decreased, but the time of reaching peak was different. There was no regular dose-effect relationship of effect that exerted onC.vulgarisby C10-HSL over time. The concentration-dependent effect of C10-HSL onC.vulgariswas not obvious. The antioxidase activities ofC.vulgariswere induced expression in different extent, which meant the microalgae was subjected to oxidative stress exerted by C10-HSL. Mg2+-ATPase activity ofC.vulgarisfluctuated fiercely,which meant the process of ATP synthesis and ion transportation were disturbed by C10-HSL. The current research not only illuminated the changing discipline of PSⅡ photochemistry activity, antioxidase activities and energy metabolism enzyme activity ofChlorellavulgaris, but also implied that the oxidative damage and turbulence of energy metabolism were supposed to the important reasons for the collapse ofC.vulgarisdensity-culture system. The results provided a basis for the axenic culture and cultivation system optimization ofC.vulgaris.
Key words: AHLs; marineChlorellavulgaris; photochemistry activity; biochemical indexes
责任编辑 高 蓓
Effects of AHLs on PSⅡ Photochemistry Activity and Biochemical Indexes ofChlorellavulgaris
DOU Yong1, GAN Yu1, TANG Xue-Xi2, ZHANG Wen-Hui1, JIANG Zhi-Fei1, GAO Jin-Wei1,ZHOU Wen-Li1
(1.Tianjin Key Lab for Aquaculture Ecology and Cultivation, Fisheries College of Tianjin Agriculture University, Tianjin 300384, China; 2. College of Marine Life Science, Ocean University of China, Qingdao 266003, China)
X173
A
1672-5174(2017)11-040-07
10.16441/j.cnki.hdxb.20170096
窦勇, 甘雨, 唐学玺, 等. AHLs对小球藻PSⅡ光化学活性与生化指标的影响研究[J]. 中国海洋大学学报(自然科学版), 2017, 47(11): 40-46.
DOU Yong, GAN Yu, TANG Xue-Xi, et al. Effects of AHLs on PSⅡ photochemistry activity and biochemical indexes ofChlorellavulgaris[J]. Periodical of Ocean University of China, 2017, 47(11): 40-46.
天津市水产产业技术体系创新团队项目(ITTFRS2017005);天津市科技重大专项与工程项目(15ZXBFNC00120);卫星海洋环境动力学国家重点实验室开放基金项目(SOED1419);农业部南海渔业资源开发利用重点实验室开放基金项目(FREU2015-04)资助 Supported by Innovation Team Program of Tianjin Fisheries Research System (ITTFRS2017005); Maior Science and Technology Project of Tianjin(15ZXBFNC00120); Fund of State Key Laboratory of Satellite Ocean Environment Dynamics(SOED1419); Fund of Key Laboratory of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture, P. R. China(FREU2015-04)
2017-04-18;
2017-06-13
窦 勇(1985-),男,博士,讲师,主要从事微藻生理生态学研究。E-mail: douyonghero@163.com
** 通讯作者:E-mail: saz0908@126.com
猜你喜欢
中国岩溶(2022年3期)2022-11-30
南方农业(2022年12期)2022-11-23
西南农业学报(2022年8期)2022-10-28
科学技术创新(2022年28期)2022-10-21
海洋湖沼通报(2022年2期)2022-05-08
古今农业(2022年1期)2022-05-05
现代盐化工(2021年3期)2021-08-27
知识就是力量(2021年6期)2021-07-09
食品与生物技术学报(2020年4期)2020-01-06
新课程·下旬(2015年10期)2015-10-21
