
生物质炭与氮肥配施对春小麦产量及其C︰N︰P的影响*
2017-10-14 03:19南学军蔡立群刘小宁高志强StephenYeboah张仁陟
中国生态农业学报(中英文) 2017年8期
南学军, 蔡立群,2, 武 均, 刘小宁, 高志强, Stephen Yeboah,3, 张仁陟,2
生物质炭与氮肥配施对春小麦产量及其C︰N︰P的影响*
南学军1, 蔡立群1,2**, 武 均1, 刘小宁1, 高志强1, Stephen Yeboah1,3, 张仁陟1,2
(1. 甘肃农业大学资源与环境学院/甘肃省干旱生境作物学省部共建国家重点实验室 兰州 730070; 2. 甘肃省节水农业工程技术研究中心 兰州 730070; 3. CSIR-Crops Research Institute, P.O. BOX 3780-Kumasi, Ghana)
碳(C)、氮(N)、磷(P)生态计量化学为研究作物-土壤生态系统物质循环及其能量流动提供了崭新视角, 研究生物质炭配施不同用量氮肥下小麦C、N、P计量特征, 可为探明区域养分限制性以及进行合理施肥等提供理论依据。本文通过田间定位试验, 测定施50 kg(N)·hm-2氮肥、100 kg(N)·hm-2氮肥、施生物质炭、生物质炭与50 kg(N)·hm-2氮肥配施、生物质炭与100 kg(N)·hm-2氮肥配施等处理下小麦产量、CNP含量及其生态化学计量等指标。结果表明: 相比空白对照(不施氮肥和生物质炭)处理, 其他不同处理均显著提高了小麦秸秆和籽粒产量, 除了单施生物质炭处理, 其他处理均不同程度提高了小麦地上部各器官N含量, 生物质炭配施不同用量氮肥显著提高了茎秆和籽粒C和P含量。计量比结果表明, 相比对照处理, 生物质炭和50 kg(N)·hm-2氮肥配施显著降低了叶片C∶N和C∶P, 生物质炭和100 kg(N)·hm-2氮肥配施处理则显著降低了茎秆C∶N、C∶P、N∶P以及籽粒C∶N、C∶P。研究区小麦叶片N∶P大多为18~23, 因此小麦可能受到P元素的限制。生物质炭配施氮肥显著提高了作物产量, 增加了小麦CNP养分含量, 降低了植物C∶N、C∶P、N∶P。总体而言, 生物质炭配施100 kg(N)·hm-2氮肥施肥措施的综合表现最优。
碳氮磷; 生态计量比; 氮肥; 生物质炭; 春小麦
生态化学计量学(ecological stoichiometry)是研究生物系统各种化学元素(C, N, P, O, S)质量的多重平衡及对生态交互作用的科学[1], 其主要基于C∶N∶P的比率来研究有机体的特性/行为与生态系统过程间的相互关系[2], 因此这使得生物学科不同层次(分子、细胞、有机体、种群、生态系统和全球尺度)的研究理论能够有机地统一起来[3-4]。作为调控有机体的特性/行为的重要指标, C、N、P计量比不仅影响寄主病原、物种共生、群落结构与动态、营养级动态、生物的养分限制[5-11], 而且对生态系统养分循环与供求平衡和全球生物地球化学循环等关系也起到重要的作用。尤其是N∶P计量比, 植物N∶P计量比的变化一方面可以反映植物体内某些组织的特性, 另一方面也说明土壤有机质的分解和矿化作用效果, 从而进一步表征植物体的生长发育状况。如有机体的C∶N∶P化学计量比与生态系统结构和功能之间的关系、植物养分利用效率及受限情况、植物生长速率等因素有关, 是决定群落结构和功能的关键性指标。
生物质炭与氮肥配施作为影响农田植物有机体行为、生长以及生理特征的农艺措施, 作为补充氮磷营养的最有效手段, 生物质炭配施氮磷肥能够改变旱作农田对土壤养分的吸收和利用, 从而改变小麦的不良生长状况并提高其光合生产力。如杜衍红等[12]认为炭-肥互作可显著提高肥料表观利用率以及氮素利用率, 从而促进作物的生长以及作物的增产增收; Huong[13]研究认为高量生物质炭添加一定氮肥显著提高了西北干旱区小麦(L.)氮磷吸收; 冯爱青等[14]也认为生物质炭的添加可以促进氮磷养分的吸收, 同时提高了土壤速效养分含量。这些变化势必会影响到植物体C、N、P养分计量比的分配与变化特征。然而, 当前从机理上以生态化学计量学理论揭示生物质炭配施氮肥对植物养分计量影响的研究少见相关报道, 因此研究施肥方式对作物CNP计量比作为农田养分生态化学计量指标的影响和作物化学计量特征对农田养分限制性的指示作用, 具有重要的科学及现实意义。
陇中黄土高原雨养农业区降水多集中在7—9月, 该区干旱缺水, 传统施肥方式直接加剧了土壤水分以及养分的流失, 土壤微生物生态环境严重恶化[15-17]。如何提升区域农田地力进而促进耕地资源的可持续发挥, 是摆在区域农业研究工作者面前亟待探讨与解决的问题。而生物质炭具有高度的稳定性、巨大的比表面积以及对氮磷养分较强的吸附能力等优点[18-19], 在改良土壤理化性质、吸附土壤养分以及调节系统内部氮磷养分平衡等方面发挥着重大作用。基于以上生物质炭所发挥的优势, 本研究在定西市李家堡镇展开生物质炭配施氮肥措施下植物碳氮磷生态化学计量学特征的研究, 并探讨区域农田碳氮磷元素的养分限制性, 为探索区域生物地球化学循环过程以及促进区域耕地地力的逐步提升等方面提供一定的理论和实际指导。
1 材料与方法
1.1 试验区概况及试验设计
研究区位于陇中黄土高原半干旱丘陵沟壑区的甘肃省定西市李家堡镇。该区海拔2 000 m左右, 年均太阳辐射594.7 kJ·cm-2, 日照时数2 476.6 h, 年均气温6.4 ℃, ≥0 ℃积温2 933.5 ℃, ≥10 ℃积温2 239.1 ℃, 无霜期140 d, 多年平均降水390.9 mm, 年蒸发量1 531 mm, 干燥度2.53, 80%保证率的降水量为365 mm, 变异系数为24.3%, 是典型的旱作农业区。土壤为典型的黄绵土, 土质绵软, 土层深厚, 质地均匀, 储水性能良好。2014年布设了生物质炭配施氮肥长期定位试验, 并于2016年展开对土壤-植物生态系统养分生态化学计量方面的研究。试验前0~30 cm土壤理化性质为: pH 8.1, 有机质16.0 g·kg-1, 全氮1.55 g·kg-1, 全磷0.82 g·kg-1, 全钾14.4 g·kg-1, 碱解氮51.1 mg·kg-1, 速效磷21.2 mg·kg-1。
本试验共设6个处理, 分别为: CK(无氮肥和生物质炭添加, 对照)、N50[氮肥50 kg(N)·hm-2、无生物质炭添加]、N100[氮肥100 kg(N)·hm-2、无生物质炭添加]、B(生物质炭15 t·hm-2, 无氮肥)、BN50[氮肥50 kg(N)·hm-2、生物质炭15 t·hm-2]、BN100[氮肥100 kg(N)·hm-2、生物质炭15 t·hm-2], 每个处理重复3次, 所有处理采用随机区组排列, 小区面积为3 m×6 m。以上各处理的氮肥从2014年开始连续每年春播前施用, 生物质炭则于2014年春播前向土壤一次性施入15 t·hm-2, 所有处理均统一施入土壤磷肥100 kg·hm-2。本研究试验小麦于2016年3月下旬用免耕播种机进行播种, 供试品种为‘定西40号’, 播量187.5 kg·hm-2, 行距20 cm, 播深7 cm, 7月底收获, 之后测定指标。供试生物质炭来源于金和福农业科技股份有限公司生产的玉米秸秆生物质炭, C含量53.28%, N含量1.04%, P含量0.26%, Ca含量0.8%, K含量0.51%, Mg含量0.47%, 灰分含量35.64%。
1.2 样品测定与分析
1.2.1 样品采集与测定
于小麦成熟期(2016年7月26日)取样。取样方法按“S”法取样, 每小区设5个0.6 m×0.5 m样方, 齐地刈割后, 准确称取各样方植株鲜重, 按茎、叶、籽粒等分开, 分开样品用蒸馏水冲洗干净后晾干, 然后在105 ℃恒温条件下杀青30 min, 再在75 ℃恒温条件下烘至恒重, 并称重。从每个样品中随机选取50株, 粉碎, 过筛, 用于测定叶片、茎部和籽粒CNP含量。C含量采用C/N联合分析仪测定; 各器官样品H2SO4-H2O2消煮, 用凯氏定氮法测定全N含量, 分光光度计法测定P含量。先计算各器官的C∶N、C∶P和N∶P的质量比, 为方便与其他作者的研究结果进行对比, 最终转换为摩尔计量比。具体计算如下:

(2)
(3)
以上各式中,C:N、C:P、N:P分别表示C∶N、C∶P和N∶P,C、N、P分别表示C、N和P元素的相对原子质量分数。
1.2.2 数据处理分析
数据处理与作图采用Microsoft Ecel 2010软件, 方差分析与显著性检验分别采用SPSS 22.0软件Duncan法和Pearson法进行。
2 结果与分析
2.1 生物质炭与氮肥配施对春小麦产量的影响
从表1可知, 相比CK处理, 不同处理均显著提高了小麦生物产量, 其中小麦生物产量以BN100处理为最高, 其增幅达54.60%, BN50增幅次之, 为31.78%, N50增幅最低, 为17.35%。不同氮肥用量条件下各处理结果表明, 无生物质炭添加条件下N50和N100处理秸秆产量显著高于CK处理, 相比CK处理, 其增幅分别为23.08%和33.07%; 生物质炭添加条件下, 相比单施生物质炭B处理, BN100处理显著提高了秸秆产量(<0.05)。籽粒产量方面, 无生物质炭添加情况下, N100处理籽粒产量较N50和CK处理差异十分显著(<0.05), 生物质炭添加下各处理差异均十分显著(<0.05), 生物质炭和氮肥的交互作用对小麦籽粒产量影响十分显著(<0.05)。
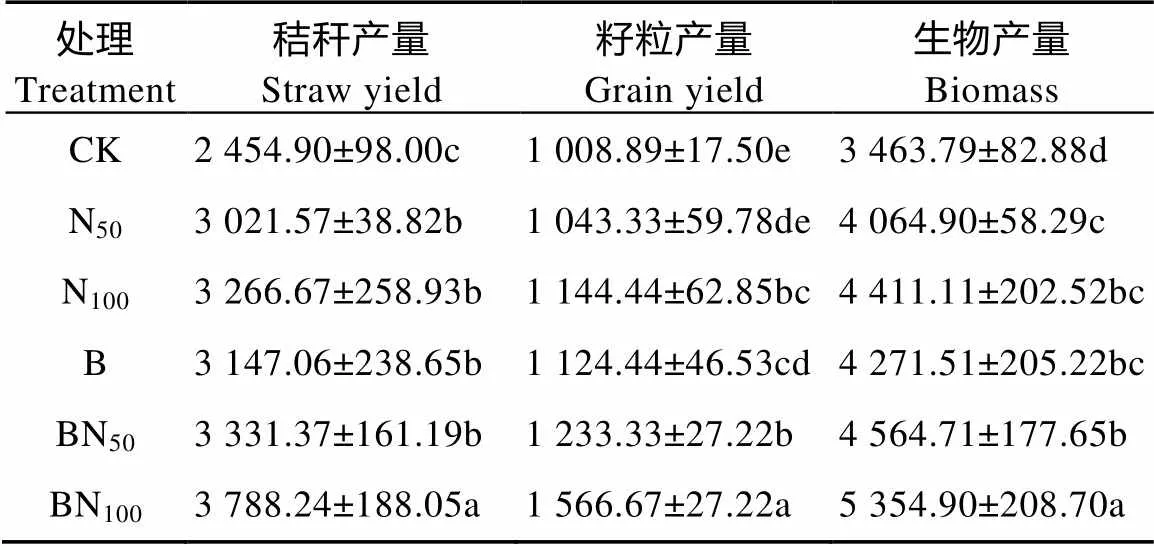
表1 生物质炭配施氮肥对春小麦产量的影响
CK: 无氮肥和生物质炭添加, 对照; N50: 氮肥50 kg(N)·hm-2、无生物质炭添加; N100: 氮肥100 kg(N)·hm-2、无生物质炭添加; B: 生物质炭15 t·hm-2, 无氮肥; BN50: 氮肥50 kg(N)·hm-2、生物质炭15 t·hm-2; BN100: 氮肥100 kg(N)·hm-2、生物质炭15 t·hm-2。数据为平均值±标准偏差(=6), 不同小写字母表示处理间在0.05水平上差异显著。CK: non N fertilizer and biocar; N50: 50 kg(N)·hm-2N fertilizer without biochar; N100: 100 kg(N)·hm-2N fertilizer without biochar; B: 15 t·hm-2biochar; BN50: 50 kg(N)·hm-2N fertilizer with 15 t·hm-2biochar; BN100: 100 kg(N)·hm-2N fertilizer with 15 t·hm-2biochar. Values are mean±S.D. (=6). Different lowercase letters indicate significant differences among treatments at 0.05 level.
2.2 生物质炭与氮肥配施对春小麦C、N、P含量的影响
由表2可知, 试验点茎秆中的C含量平均值为381.17 g·kg-1, 叶片和籽粒C含量平均分别为353.30 g·kg-1和363.83 g·kg-1。方差分析表明, 无生物质炭添加下不同处理小麦叶片C含量差异不显著, 生物质炭添加下BN50和BN100处理则不同程度提高了小麦叶片、茎秆和籽粒C含量, 其中BN50相比CK处理各器官分别提高3.5%、2.49%、1.9%, BN100则分别提高4.13%、2.23%、2.34%, 但二者之间差异不显著(>0.05)。单施生物质炭并未显著提高不同器官C含量(>0.05)。
如表2, 植株籽粒、叶片和茎秆N含量分别为22.59 g·kg-1、8.9 g·kg-1和6.05 g·kg-1。除B处理, 其他施肥措施均不同程度提高了小麦籽粒N含量, 尤其以BN100处理为最高, 其次为BN50处理, B处理N含量为最小; 叶片含量同样以BN100处理为最高, 其次为N100处理, CK对照处理叶片N含量为最小。茎秆N含量以BN100的为最高, 其次为BN50处理, CK对照处理为最小。整体来看, 除B处理, 其他不同处理均不同程度地提高了地上部器官N含量, 其籽粒N含量依次为BN100、BN50>N100>N50>CK、B; 叶片含量大小依次为BN100、N100>BN50>N50>CK、B; 茎秆N含量依次为BN100>BN50>N100>N50>CK、B。
如表2, 植株籽粒P、叶片和茎秆P含量分别为2.29 g·kg-1、1.04 g·kg-1和0.64 g·kg-1。方差分析表明, 无生物质炭添加下, 小麦不同地上器官P含量差异不显著; 生物质炭添加条件下, 随着氮肥用量的提高, 小麦地上各器官P含量有上升趋势, 且相比B处理, BN100处理显著提高了小麦籽粒P含量, 其增幅达8.33%, 茎秆和叶片P含量各处理之间无显著性差异; 同等氮肥用量条件下, 生物质炭添加显著提高了茎秆和籽粒P含量, 其中茎秆和籽粒P含量以BN100处理增幅最大, 其增幅为33.93%和9.78%, BN50处理茎秆和籽粒P含量增幅次之, 相比N50处理, 其增幅分别为32.73%和10.10%, 除茎秆P含量相对提高外, 单施生物质炭并未提高叶片和籽粒P含量。
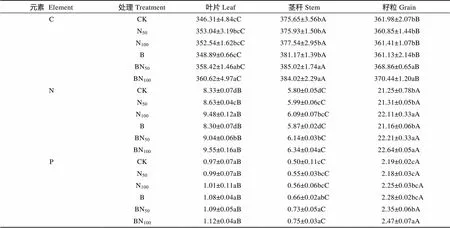
表2 生物质炭配施氮肥对春小麦C、N、P含量的影响
CK: 无氮肥和生物质炭添加, 对照; N50: 氮肥50 kg(N)·hm-2、无生物质炭添加; N100: 氮肥100 kg(N)·hm-2、无生物质炭添加; B: 生物质炭15 t·hm-2, 无氮肥; BN50: 氮肥50 kg(N)·hm-2、生物质炭15 t·hm-2; BN100: 氮肥100 kg(N)·hm-2、生物质炭15 t·hm-2。数据为平均值±标准偏差(=6), 不同小写和大写字母均表示处理间在0.05水平上差异显著, 其中小写字母表示处理之间的差异显著, 大写字母则表示地上部器官之间的差异显著。CK: non N fertilizer and biocar; N50: 50 kg(N)·hm-2N fertilizer without biochar; N100: 100 kg(N)·hm-2N fertilizer without biochar; B: 15 t·hm-2biochar; BN50: 50 kg(N)·hm-2N fertilizer with 15 t·hm-2biochar; BN100: 100 kg(N)·hm-2N fertilizer with 15 t·hm-2biochar. Values are mean±S.D. (=6). Different lowercase and capital letters indicate significant differences among treatments at 0.05 level, where the lowercase letters indicate the significance of the processing, and the capital letters indicate the difference of above organs.
2.3 生物质炭与氮肥配施对春小麦C、N、P计量比的影响
2.3.1 地上部器官C∶N
从C∶N生态化学计量特征来看(表3), 不同器官叶片、茎秆和籽粒C∶N的范围分别为44.07~49.06、71.50~75.72以及19.01~19.91, 变异系数分别为4.26%、2.12%以及1.91%。不同处理小麦茎秆C∶N显著高于叶片和籽粒C∶N, 且叶片和籽粒C∶N差异显著。从不同处理C∶N变化趋势来看, 无论是否有生物质炭添加, 随着氮肥用量的增加, 小麦地上器官碳氮比表现为不同幅度的下降趋势。同等氮肥用量条件下, 除BN50处理叶片C∶N相对下降以外, 其他处理不同器官C∶N均无显著差异(>0.05)。总体来看, 各器官C∶N均在BN100处理达到最小值, 其叶片、茎秆和籽粒较CK处理分别下降了4.43、4.06和0.89个单位。
2.3.2 地上部器官C∶P
如表3, 植物叶片、茎秆和籽粒C∶P分别为830.01~940.55、1 347.83~2 055.37以及385.51~428.36, 变异系数分别为5.39%、15.31%和3.47%。除N100处理小麦茎秆C∶P较高, 且叶片和籽粒之间无显著差异以外, 其他处理小麦C∶P茎秆显著高于叶片和籽粒, 且叶片和籽粒差异十分显著。从不同处理C∶P变化趋势来看, 无生物质炭添加下, 小麦叶片、茎秆和籽粒C∶P表现出增加的趋势。生物质炭添加下, 小麦地上器官C∶P则表现较为稳定。同等氮肥用量对比结果表明, 相比N100处理, BN100处理茎秆和籽粒C∶P显著降低(<0.05), BN50处理籽粒C∶P显著低于N50处理。总体来看, 叶片和茎秆C∶P在N100处理达到最大值, 分别较CK处理增加了42.95和301.77个单位, 籽粒C∶P在N50处理达到最大值, 且相比CK处理增加12.09个单位。
2.3.3 地上部器官N∶P
如表3, 小麦N∶P叶片、茎秆和籽粒分别为17.10~21.68、18.72~28.41以及20.28~22.33, 变异系数分别为7.19%、14.90%以及3.99%。不同器官N∶P分布特征结果表明, 无生物质炭添加下, CK和N50处理茎秆N∶P显著高于叶片N∶P, N100处理各器官N∶P则无显著性差异(>0.05)。生物质炭添加下, B和BN50处理籽粒N∶P显著高于叶片N∶P, BN100处理下各器官N∶P则无显著性差异(>0.05)。总体来看, 无生物质炭添加下各处理茎秆N∶P为最高, 其次为籽粒, 叶片N∶P为最低, 生物质炭添加下各处理籽粒N∶P为最高, 其次为茎秆, 叶片N∶P为最低, 因此表现出较大的波动和敏感性。
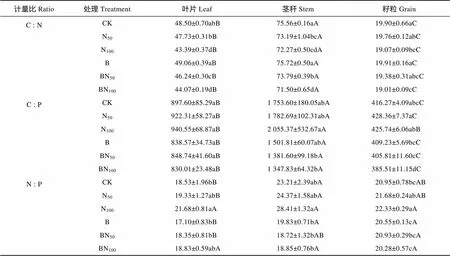
表3 生物质炭配施氮肥对春小麦C∶N、C∶P、N∶P的影响
CK: 无氮肥和生物质炭添加, 对照; N50: 氮肥50 kg(N)·hm-2、无生物质炭添加; N100: 氮肥100 kg(N)·hm-2、无生物质炭添加; B: 生物质炭15 t·hm-2, 无氮肥; BN50: 氮肥50 kg(N)·hm-2、生物质炭15 t·hm-2; BN100: 氮肥100 kg(N)·hm-2、生物质炭15 t·hm-2。数据为平均值±标准偏差(=6), 不同小写和大写字母均表示处理间在0.05水平上差异显著, 其中小写字母表示处理之间的差异显著, 大写字母则表示地上部器官之间的差异显著。CK: non N fertilizer and biocar; N50: 50 kg(N)·hm-2N fertilizer without biochar; N100: 100 kg(N)·hm-2N fertilizer without biochar; B: 15 t·hm-2biochar; BN50: 50 kg(N)·hm-2N fertilizer with 15 t·hm-2biochar; BN100: 100 kg(N)·hm-2N fertilizer with 15 t·hm-2biochar. Values are mean±S.D. (=6). Different lowercase and capital letters indicate significant differences among treatments at 0.05 level, where the lowercase letters indicate the significance of the processing, and the capital letters indicate the difference of above organs.
从不同处理N∶P变化趋势来看, 无生物质炭添加下, 小麦叶片、茎秆和籽粒N∶P呈现增高趋势; 生物质炭添加下, 小麦叶片N∶P呈现下降趋势, 茎秆和籽粒N∶P则具有一定波动性。同等氮肥用量条件下, 除BN100处理则显著降低了茎秆和籽粒N∶P外(<0.05), 其他处理均未显著改变N∶P。总体来看, 叶片、茎秆和籽粒N∶P均在N100处理达到最大值, 分别较CK处理增加3.15、5.2和1.38个单位。
3 讨论
3.1 小麦产量对生物质炭配施氮肥的响应
冯爱青等[14]的连续2年黑炭和氮肥配施试验表明, 小麦产量在第2年内显著增加, 施氮配施黑炭较施氮处理籽粒显著增产8.19%, 不施氮条件下, 黑炭施加较不施氮肥处理籽粒及秸秆均增产显著。Chan等[20]发现氮肥与高量生物质炭(100 t·hm-2)配施, 萝卜(Linn.)产量的增幅达95%~266%。本研究表明, 无论是否施氮肥, 添加生物质炭均可显著增加籽粒产量, 尤其是100 kg(N)·hm-2氮肥配施生物质炭处理小麦籽粒产量显著高于不施氮肥对照处理, 其增幅达32.99%, 这与上述学者的研究结果一致。究其原因, 主要在于生物炭的高度稳定性和较强的吸附能力赋予肥料养分缓释性能, 从而与肥料形成互补协同的关系; 生物质炭与氮肥配施对产量的影响比单施生物质炭效果更显著, 这除了与生物质炭自身含有较少的养分有关之外, 还可能与生物质炭配施氮肥可以显著改善土壤孔隙度和持水性能[21]、增加土壤pH以及减少活性铝[22]、增强K、P、Mg、Ca等营养元素的有效性[23]、提高阳离子交换量[24]以及与作物类型等密切相关[25], 其具体原因还有待进一步研究。
3.2 植物C、N、P含量对生物质炭配施氮肥的响应
本研究表明, 无论是否有生物质炭的添加, 随着施氮量增加, 小麦地上部分C含量呈逐渐上升趋势, 但不同处理小麦叶片碳含量均远低于Elser等[2]研究的全球492种陆生植物叶片碳平均含量464 g·kg-1,说明该研究区植物的有机质含量和碳储量较低。不同处理的结果表明, 相比生物质炭添加各处理, 无生物质炭处理小麦C含量有提高趋势, 但差异性不显著; 而在生物质炭添加100 kg(N)·hm-2氮肥的条件下, 籽粒C素含量达到370.44 g·kg-1, 相比CK处理其增幅达2.34%。这可能与生物质炭本身较高的C含量有关。生物质炭和氮肥的施用, 小麦秸秆干物质量的生产和枯落物的分解增加了植物和土壤有机质总量, 从而改善了小麦根系的生长, 促进了地上部光合功能的发挥, 并最终导致小麦C素含量的相应提高。单施生物质炭并未提高器官C含量, 这是由于植物体主要通过叶片的光合作用进行C素的吸收和固定, 因此其C含量相对变化较小。
研究区小麦地上部分N平均含量为10.51 g·kg-1, 远低于Han等[26]研究的全国753种陆地植物氮平均含量18.6 g·kg-1, 这可能与该地区土壤N含量较低有关。从不同器官来看, 随着氮肥和生物质炭的施入, 小麦籽粒产量显著提高, 小麦的生长促进了N素向籽粒器官的再分配, 受到稀释效应的影响, 从而导致植物茎秆和叶片氮含量相对较低。另一方面, 从不同处理氮磷含量来看, 由于本研究有一定的磷肥作为补充, 因此生物质炭添加下小麦不同器官氮磷含量显著提高, 这与杜衍红、陈心想等的研究结论十分相似[12,27]。生物质炭具有较高的N、P、K、Mg含量, 因此配施不同量氮肥可以弥补单施生物质炭供给N素不足的缺陷, 这就在一定程度上能够显著提高肥料的表观利用率以及氮素利用率, 从而造成土壤C∶N的下降, 进而进一步提高了N素的有效性, 最终使得小麦叶片、茎秆和籽粒N含量得以提高。和N素含量类似, 生物质炭配施氮肥利于植物对P素的吸收。这可能与生物质炭配施氮肥显著提高了土壤P含量和某些微量元素的有效性, 并提高了土壤速效磷等有效养分含量等因素有关。由于环境中养分在供应短缺的情况下以提高养分重吸收率为其养分利用的主导方式适应环境, 而在土壤养分供应相对富足情况下植物则主要以提高养分吸收能力的方式适应环境, 因此小麦在衰老过程中P含量可能存在较高的转移率, 养分重吸收明显[28]。
3.3 生态化学计量比对生物质炭配施氮肥的响应
N∶P能较好地反映N、P养分的限制作用, 是土壤向植物提供养分状况的重要指标, 同时对植物生产力限制性起重要的指示作用[29]。C∶N和C∶P则能够反映植物养分利用效率的高低状况[9]。不同人为措施下植物可以通过不同的生理生化反应调节C、N、P的代谢和循环特征, 从而使植物体最终表现出特定的元素生态化学计量特征。从小麦C∶N和C∶P角度来看, 随着氮肥用量的增加以及生物质炭的补充, 小麦C∶N显著降低, 而C∶P则相对升高, 这说明植物在N素营养元素供应缺乏的情况下往往具有较高的N素利用效率, 同时表明植物体N同化C的能力较强, 这可能是植物适应养分贫乏状态的生存策略。而从N∶P比角度来看, Yan等[30]对黄土高原施肥措施下小麦计量比研究结果表明, 随着氮肥用量的升高, 小麦茎秆和籽粒N含量显著增加, P含量则呈现下降趋势, N∶P显著增加, 本研究结果则不尽一致, 可能是由于试验处理有一定的磷肥补充, 氮磷养分的交互作用使得植物体P素含量有所上升, 因此N∶P相对有所增加。而生物质炭配施不同用量氮肥下植物N∶P则相对恒定, 且相比无生物质炭添加各处理, 生物质炭与氮肥配施处理显著降低了小麦N∶P, 这可能是由于生物质炭本身含有丰富的有机大分子和发达的孔隙结构, 施入土壤后又较易形成大团聚体[31], 因此对土壤中的NH4+-N、NO3--N以及有效磷等不同形态的营养元素有很强的吸附作用, 因此可以进一步影响到作物N∶P比在地上部各器官之间分配与代谢过程。
3.4 植物N∶P比及其养分的限制性
从植物N∶P计量比的养分限制性来看, 一般认为, 低N∶P意味着群落缺N, 而对于高N∶P的生态学机制却还存在争议。由于土壤养分有效性、物种以及植物年龄在不同研究区域、生态系统或植被类型变异很大, 因此N∶P表现出较大的变异性, 从而使得判断N、P限制作用的N∶P值不同, N∶P临界指标变化很大。如有些研究者认为高N∶P说明植被生产力受P限制, 而有些学者观点则认为高N∶P比还有可能是N、P共同限制的结果。如Güsewell[32]认为植物N∶P比<13和>16分别表示植被受N限制和P限制, 而N∶P在两者之间则为N、P共同限制。Tessier和Koerselman等[9-10]对湿地生态系统进行施肥试验结果显示, 当植物体N∶P>16为P限制, N∶P<14则受N限制。Braakhekke和Hooftman[33]则认为当N∶P大于14而植物叶片P含量低于1.0 g·kg-1时, 这个系统受P限制; 而当N∶P小于10, 而植物叶片N含量低于20.0 g·kg-1时, 则该生态系统受N限制; N∶P比在10~14范围内被认为是受到2种元素共同限制(N<20.0 g·kg-1、P<1.0 g·kg-1)或两种元素都不缺少(N>20.0 g·kg-1、P>1.0 g·kg-1)。而Zhang等[34]对我国内蒙古地区草原的施肥试验表明, 当N∶P>23时表现为P限制, N∶P<21则表现为N限制。研究区小麦叶片N素在10 g·kg-1以下, P素含量则在1.0 g·kg-1左右, N∶P为18~23, 安卓等[28]氮素添加试验认为氮肥加强了P素对植物生长的限制, 因此旱作农田小麦可能受到P的限制。
从全球N、P限制类型的全球分布模式来看, 全球气候变化加剧热带地区P的限制, 而高海拔或高纬度地区, 限制类型有从N限制转向P限制的趋势。根据汪涛等[35]对中国第2次土壤普查数据分析结果显示, 黄土高原黄绵土土壤的有效磷含量较全国平均值要低(黄绵土有效磷密度为1.9 g·m-2, 全国土壤有效磷平均密度为3.4 g·m-2)。Zheng等[36]通过对黄土高原7个不同地区的126种植物叶片化学计量特征进行研究的结果认为, 黄土高原地区不同生活型植物叶片N∶P计量比平均值为14.9, 且各地区N∶P比总体上随着纬度和年太阳辐射量的升高而升高, 但随年平均降雨量和年平均温度的下降, 叶片N∶P比值则升高。本研究N∶P比显著高于Zheng等[36]测定的禾本科叶片计量比, 由于研究区地处黄土高原西部, 因此太阳辐射较强, 而降水则相比较少, 受到太阳辐射和降水的影响, 从而使得CNP计量比相对较高, 再次证明了试验区受到P素的限制观点, 符合Güsewell[32]提出的N∶P比值的临界值假说。因此在本研究中相较于N限制而言, 小麦种群可能更多地受P素的限制。
4 结论
研究区小麦地上器官C、N、P含量的影响随着生物质炭和氮肥等处理方式的差异呈现出较大的差异性, 其中100 kg(N)·hm-2氮肥配施生物质炭的小麦各器官的碳含量大于对照处理以及单施生物质炭的C含量, 且小麦N、P含量随着氮肥用量的提高呈上升趋势。不同处理小麦地上器官C∶N、C∶P、N∶P因作物C、N、P含量的不同也发生较大变异, 尤其是生物质炭配施100 kg(N)·hm-2氮肥措施下小麦光合产量最高, 此时其茎秆和籽粒C∶N最小, 而C∶P比则最低, 50 kg(N)·hm-2氮肥配施生物炭处理下小麦N∶P为最低。但与此同时, 生物质炭配施氮肥措施进一步加强了P素对植物生长的限制, 从而促使研究区小麦由N限制向P素限制的转变。由于本研究在一定程度上探讨了生物质炭添加氮肥对农田作物化学计量学特征的影响, 但生物质炭配施氮肥对于作物的影响是一个长期的过程, 受温度、降水等气候条件以及土壤理化性质等因素的多重影响, 只有通过多年的连续观测才能完整地揭示旱作农田生态系统对生物质炭配施氮肥方式的响应。此外,考虑到旱作农田在恢复建设过程中, 为防止养分的消耗和流失以及进一步促进旱农区耕地资源的可持续发展, 研究建议增加不同用量磷肥, 并配合一定的生物质炭进行组合施用。
[1] Tang K W, Dam H G. Limitation of zooplankton production: Beyond stoichiometry[J]. Oikos, 1999, 84(3): 537–542
[2] Elser J J, Sterner R W, Gorokhova E, et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000, 3(6): 540–550
[3] 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索[J]. 植物生态学报, 2005, 29(6): 1007–1019 Zeng D H, Chen G S. Ecological stoichiometry: A science to explore the complexity of living systems[J]. Acta Phytoecologica Sinica, 2005, 29(6): 1007–1019
[4] Zhang L X, Bai Y F, Han X G. Application of N∶P stoichiometry to ecology studies (English)[J]. Acta Botanica Sinica, 2003, 45(9): 1009–1018
[5] Smith V H. Applicability of resource-ratio theory to microbial ecology[J]. Limnology and Oceanography, 1993, 38(1): 239–249
[6] Smith V H. Implications of resource-ratio theory for microbial ecology[M]//Jones J G. Advances in Microbial Ecology. New York: Springer, 1993: 1–37
[7] Urabe J, Kyle M, Makino W, et al. Reduced light increases herbivore production due to stoichiometric effects of light/nutrient balance[J]. Ecology, 2002, 83(3): 619–627
[8] Sterner R W, Hessen D O. Algal nutrient limitation and the nutrition of aquatic herbivores[J]. Annual Review of Ecology and Systematics, 1994, 25(1): 1–29
[9] Tessier J T, Raynal D J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation[J]. Journal of Applied Ecology, 2003, 40(3): 523–534
[10] Koerselman W, Meuleman A F M. The vegetation N∶P ratio: A new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 1996, 33(6): 1441–1450
[11] Aerts R, Chapin Ⅲ F S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns[J]. Advances in Ecological Research, 1999, 30: 1–67
[12] 杜衍红, 蒋恩臣, 王明峰, 等. 炭-肥互作对芥菜产量和肥料利用率的影响[J]. 农业机械学报, 2016, 47(4): 59–64 Du Y H, Jiang E C, Wang M F, et al. Effect of interaction of biochar and fertilizer on mustard yield and fertilizer utilization rate[J]. Transactions of the CSAM, 2016, 47(4): 59–64
[13] Huong N T. 生物质炭对西北地区土壤质量及作物产量的影响[D]. 杨凌: 西北农林科技大学, 2016 Huong N T. Effects of biochar amendment on soil quality, crop yield in Northwest China[D]. Yangling: Northwest A&F University, 2016
[14] 冯爱青, 张民, 李成亮, 等. 秸秆及秸秆黑炭对小麦养分吸收及棕壤酶活性的影响[J]. 生态学报, 2015, 35(15): 5269–5277 Feng A Q, Zhang M, Li C L, et al. Effects of straw and straw biochar on wheat nutrient uptake and enzyme activity in brown soil[J]. Acta Ecologica Sinica, 2015, 35(15): 5269–5277
[15] 蔡立群, 齐鹏, 张仁陟, 等. 不同保护性耕作措施对麦——豆轮作土壤有机碳库的影响[J]. 中国生态农业学报, 2009, 17(1): 1–6 Cai L Q, Qi P, Zhang R Z, et al. Effects of different conservation tillage measures on soil organic carbon pool in two sequence rotation systems of spring wheat and pease[J]. Chinese Journal of Eco-Agriculture, 2009, 17(1): 1–6
[16] 罗珠珠, 蔡立群, 李玲玲, 等. 长期保护性耕作对黄土高原旱地土壤养分和作物产量的影响[J]. 干旱地区农业研究, 2015, 33(3): 171–176 Luo Z Z, Cai L Q, Li L L, et al. Long-term effects of tillage system on soil nutrients and grain yields in rainfed area of Loess Plateau[J]. Agricultural Research in the Arid Areas, 2015, 33(3): 171–176
[17] 蔡艳, 郝明德, 臧逸飞, 等. 不同轮作制下长期施肥旱地土壤微生物多样性特征[J]. 核农学报, 2015, 29(2): 344–350 Cai Y, Hao M D, Zang Y F, et al. Effect of long-term fertilization on microbial diversity of black loessial soil based on 454 sequencing technology[J]. Journal of Nuclear Agricultural Sciences, 2015, 29(2): 344–350
[18] 陈温福, 张伟明, 孟军. 农用生物炭研究进展与前景[J]. 中国农业科学, 2013, 46(16): 3324–3333 Chen W F, Zhang W M, Meng J. Advances and prospects in research of biochar utilization in agriculture[J]. Scientia Agricultura Sinica, 2013, 46(16): 3324–3333
[19] Lehmann J, Gaunt J, Rondon M. Bio-char sequestration in terrestrial ecosystems: A review[J]. Mitigation and Adaptation Strategies for Global Change, 2006, 11(2): 395–419
[20] Chan K Y, Van Zwieten L, Meszaros I, et al. Agronomic values of green waste biochar as a soil amendment[J]. Australian Journal of Soil Research, 2007, 45(8): 629–634
[21] Busscher W J, Novak J M, Evans D E, et al. Influence of pecan biochar on physical properties of a Norfolk loamy sand[J]. Soil Science, 2010, 175(1): 10–14
[22] 崔立强. 生物黑炭抑制稻麦对污染土壤中Cd/Pb吸收的试验研究[D]. 南京: 南京农业大学, 2011 Cui L Q. An experiment study on reducing cadmium and lead uptake from polluted soil by rice and wheat with biochar amendment[D]. Nanjing: Nanjing Agricultural University, 2011
[23] Laird D, Fleming P, Wang B Q, et al. Biochar impact on nutrient leaching from a Midwestern agricultural soil[J]. Geoderma, 2010, 158(3/4): 436–442
[24] 陈红霞, 杜章留, 郭伟, 等. 施用生物炭对华北平原农田土壤容重、阳离子交换量和颗粒有机质含量的影响[J]. 应用生态学报, 2011, 22(11): 2930–2934 Chen H X, Du Z L, Guo W, et al. Effects of biochar amendment on cropland soil bulk density, cation exchange capacity, and particulate organic matter content in the North China Plain[J]. Chinese Journal of Applied Ecology, 2011, 22(11): 2930–2934
[25] 周加顺, 郑金伟, 池忠志, 等. 施用生物质炭对作物产量和氮、磷、钾养分吸收的影响[J]. 南京农业大学学报, 2016, 39(5): 791–799 Zhou J S, Zheng J W, Chi Z Z, et al. Effects of biochar amendment on crop yield and the uptake of nitrogen, phosphorus and potassium[J]. Journal of Nanjing Agricultural University, 2016, 39(5): 791–799
[26] Han W X, Fang J Y, Guo D L, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377–385
[27] 陈心想, 何绪生, 耿增超, 等. 生物炭对不同土壤化学性质、小麦和糜子产量的影响[J]. 生态学报, 2013, 33(20): 6534–6542 Chen X X, He X S, Geng Z C, et al. Effects of biochar on selected soil chemical properties and on wheat and millet yield[J]. Acta Ecologica Sinica, 2013, 33(20): 6534–6542
[28] 安卓, 牛得草, 文海燕, 等. 氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C∶N∶P化学计量特征的影响[J]. 植物生态学报, 2011, 35(8): 801–807 An Z, Niu D C, Wen H Y, et al. Effects of N addition on nutrient resorption efficiency and C∶N∶P stoichiometric characteristics inof steppe grasslands in the Loess Plateau, China[J]. Chinese Journal of Plant Ecology, 2011, 35(8): 801–807
[29] Güsewell S. N∶P ratios in terrestrial plants: Variation and functional significance[J]. New Phytologist, 2004, 164(2): 243–266
[30] Yan W, Zhong Y, Shangguan Z. The relationships and sensibility of wheat C∶N∶P stoichiometry and water use efficiency under nitrogen fertilization[J]. Plant Soil Environment, 2015, 61(5): 201–207
[31] Brodowski S, John B, Flessa H, et al. Aggregate-occluded black carbon in soil[J]. European Journal of Soil Science, 2006, 57(4): 539–546
[32] Güsewell S. Nutrient resorption of wetland graminoids is related to the type of nutrient limitation[J]. Functional Ecology, 2005, 19(2): 344–354
[33] Braakhekke W G, Hooftman D A P. The resource balance hypothesis of plant species diversity in grassland[J]. Journal of Vegetation Science, 1999, 10(2): 187–200
[34] Zhang L X, Bai Y F, Han X G. Differential responses of N∶P stoichiometry ofandto N additions in a steppe ecosystem in Nei Mongol[J]. Acta Botanica Sinica, 2004, 46(3): 259–270
[35] 汪涛, 杨元合, 马文红. 中国土壤磷库的大小、分布及其影响因素[J]. 北京大学学报: 自然科学版, 2008, 44(6): 945–952 Wang T, Yang Y H, Ma W H. Storage, patterns and environmental controls of soil phosphorus in China[J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 2008, 44(6): 945–952
[36] Zheng S X, Shangguan Z P. Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China[J]. Trees, 2007, 21(3): 357–370
Effect of combined application of biochar and N-fertilizer on yield and C∶N∶P ratio of spring wheat*
NAN Xuejun1, CAI Liqun1,2**, WU Jun1, LIU Xiaoning1, GAO Zhiqiang1, Stephen Yeboah1,3, ZHANG Renzhi1,2
(1. College of Resources and Environmental Sciences, Gansu Agricultural University / Gansu Provincial Key Laboratory of Arid Land Crop Science, Gansu Agricultural University, Lanzhou 730070, China; 2. Gansu Province Research Center for Water-saving Agriculture Engineering Technology, Lanzhou 730070, China; 3. CSIR-Crops Research Institute, P.O. BOX 3780-Kumasi, Ghana)
Carbon (C), nitrogen (N) and phosphorus (P) are the 3 main elements existing in living organisms. It has long been known that C∶N∶P stoichiometry of consumers and their resources affect both the structure and function of the food web that eventually influence broad-scale processes such as global carbon cycles. The balance of these nutrients can influence plant growth and its stoichiometry in ecosystems. Wheat is one of the most important food crops and wheat production affects global food security. The combined application of biochar and N fertilizer has been reported to be a beneficial agronomic measure, and noted to affect grain yield and C∶N∶P allocation patterns in wheat. In addition, the study of different N fertilizer and biochar on wheat yield, C, N and P contents and their ratio have been closely linked with nutrient limitation and rational fertilization. In this study, we explored the yield and C, N and P contents and their ratios of wheat under different N-fertilizer and biochar treatments (50 kg·hm-2N-fertilizer, 100 kg·hm-2N-fertilizer, biochar, biochar combined with 50 kg·hm-2N-fertilizer, biochar combined with 100 kg·hm-2N-fertilizer and no N-fertilizer or biochar as control). The results showed that different treatments significantly increased wheat straw and grain yield, compared with that of control treatment. Except for the single biochar application, other treatments increased N contents of wheat organs. Biochar combined with different doses of N-fertilizer significantly increased C and P contents of both wheat stem and grain. Biochar combined with 50 kg·hm-2N-fertilizer significantly decreased leaf C∶N and C∶P ratios. Then biochar combined with 100 kg·hm-2N-fertilizer significantly reduced stem C∶N, C∶P and N∶P ratios and grain C∶N and C∶P ratios. In terms of nutrient limitation, N∶P ratio in wheat leaf in the study area was 18‒23, wheat may be limited by P element. Biochar combined with N-fertilizer significantly increased wheat yield and CNP nutrient contents. At the same time, it decreased wheat C∶N, C∶P and N∶P ratios. From the above, the comprehensive performance of biochar plus 100 kg·hm-2N-fertilizer was the best pattern to enhance wheat productivity.
CNP; Ecological stoichiometry; Nitrogen fertilizer; Biochar; Spring wheat
10.13930/j.cnki.cjea.161184
S532.01
A
1671-3990(2017)08-1154-09
* 国家自然科学基金面上项目(3157101843)、“十二五”《循环农业科技工程》项目(2012BAD14B03)和甘肃省自然科学基金项目(145RJZA204, 145RJZA106)资助
**通讯作者:蔡立群, 主要从事土壤生态学和保护性耕作研究等工作。E-mail: cailq@gsau.edu.cn
南学军, 研究方向为恢复生态学。E-mail: 1530679768@qq.com
2016-12-28 接受日期: 2017-02-21
* This study was supported by the National Natural Science Foundation of China (3157101843), the Project of the Twelfth Five-Year Plan for Circular Agriculture Science and Technology of China (2012BAD14B03) and the Natural Science Foundation of Gansu Province (145RJZA204, 145RJZA106).
, E-mail: cailq@gsau.edu.cn
Dec. 28, 2016; accepted Feb. 21, 2017
南学军, 蔡立群, 武均, 刘小宁, 高志强, Stephen Yeboah, 张仁陟. 生物质炭与氮肥配施对春小麦产量及其C∶N∶P的影响[J]. 中国生态农业学报, 2017, 25(8): 1154-1162
Nan X J, Cai L Q, Wu J, Liu X N, Gao Z Q, Stephen Yeboah, Zhang R Z. Effect of combined application of biochar and N-fertilizer on yield and C∶N∶P ratio of spring wheat[J]. Chinese Journal of Eco-Agriculture, 2017, 25(8): 1154-1162
猜你喜欢
中国化肥信息(2022年9期)2022-11-23
江西农业大学学报(2022年3期)2022-07-07
今日农业(2021年21期)2022-01-12
能源工程(2021年5期)2021-11-20
生物质化学工程(2021年1期)2021-01-26
农业机械学报(2020年11期)2020-11-25
中国造纸(2020年9期)2020-10-20
中国化肥信息(2019年4期)2019-05-31
农业机械学报(2019年4期)2019-04-29
中国化肥信息(2019年3期)2019-04-25
