
福建地区马尾松生物量转换和扩展因子的影响因素
2017-10-13 12:32:58欧强新李海奎
生态学报 2017年17期
欧强新,李海奎,*,杨 英
1 中国林业科学研究院资源信息研究所, 北京 100091 2 国家林业局调查规划设计院, 北京 100714
福建地区马尾松生物量转换和扩展因子的影响因素
欧强新1,李海奎1,*,杨 英2
1 中国林业科学研究院资源信息研究所, 北京 100091 2 国家林业局调查规划设计院, 北京 100714
基于第8次国家森林资源清查福建省331块马尾松的固定样地调查数据,利用增强回归树法(BRT)研究地上生物量转换和扩展因子(BCEF)和地下BCEF的影响因素。研究结果表明: 林分特征因子和地形因子是影响地上BCEF以及地下BCEF的主导因素,二者对地上BCEF影响的相对贡献率之和为87.20%、地下BCEF为86.59%。其中,龄组和坡向分别是林分特征因子和地形因子中影响地上BCEF的最大因素(41.13%和14.52%)。地上BCEF随龄组的增大而逐渐减小;在东南坡最大、西坡最小。此外,龄组和坡向分别是林分特征因子和地形因子中影响地下BCEF的最大因素(41.54%和15.16%)。地下BCEF随龄组的增大而逐渐减增大;在东南坡最小、西坡最大。土壤因子对地上BCEF以及地下BCEF的影响都较小(12.80%和13.41%),腐殖层厚度是土壤因子中影响地上BCEF以及地下BCEF的最大因素(9.02%和9.13%)。在所有的影响因素中,龄组对地上BCEF以及地下BCEF的影响均最大,依据龄组计算相应的BCEF或者建立林龄普适的BCEF模型,可以有效地提高生物量的估算精度。
马尾松;生物量转换和扩展因子(BCEF);国家森林资源清查;增强回归树(BRT)
Abstract: In this study, the boosted regression trees (BRT) method was used to analyze the main factors that influenced the above- and underground biomass conversion and expansion factor (BCEF) of Masson pine. Data from the permanent sample plot of the 8thChinese National Forest Inventory for Fujian Province was used. The results showed that the cumulative relative influences of stand characteristic and topographic factors on above- and underground BCEF were 87.20 and 86.59%, respectively. Stand characteristic and topographic factors were the most important factors for both above- and underground BCEF. Among the stand characteristic factors, the age group was dominant, contributing to 41.13% of the aboveground BCEF differences that decreased with an increase in the age group, and 41.54% of the underground BCEF differences that increased with an increase in the age group. Among the topographic factors, slope was the key variable, contributing to 14.52% of aboveground BCEF differences, which were the highest at the southeast slope and the lowest at the west slope, and 15.16% of the underground BCEF differences, which were the lowest at the southeast slope and the highest at the west slope. In addition, soil factors had a lower effect on both above and underground BCEF. The relative influences of soil factors on above- and underground BCEF were 12.80 and 13.41%, respectively. Among the soil factors, the thickness of the humus layer was the key variable, contributing to 9.02% of the aboveground BCEF differences and 9.13% of the underground BCEF differences. Among all influencing factors, age group had the highest effect on both above- and underground BCEF. Consequently, the ways, such as calculating BCEF based on age group or building universal BCEF models of all age, could achieve a better precision of biomass.
KeyWords: Masson pine; biomass conversion and expansion factor (BCEF); national forest inventory; boosted regression trees (BRT)
森林生物量是评估森林碳收支的重要参数[1- 2],准确的生物量估算对量化碳固定率、评估气候变化的潜在影响等内容至关重要[3- 4]。生物量转换和扩展因子(BCEF)是估算森林生物量普遍使用的估算参数[5],BCEF可以直接将蓄积转换并扩展为生物量[6-13]。对林分BCEF的影响因素进行研究,确定影响BCEF的主要因素,可提高林分生物量估算的准确性[14]。已有的研究表明,BCEF并非是一种固定不变的常数[7,15-17],而与立地、林龄等因子密切相关[15,18-21]。如Lehtonen等[18]发现欧洲赤松(Pinussylvestris)和欧洲云杉(Piceaabies)总BCEF均随着林分年龄的增加而变小,而地下BCEF均随着林分年龄的增加而变大;Soares and Tomé[15]发现蓝桉树(Eucalyptusglobulus)总BCEF随立地指数的增加而减小;González-García等[20]发现亮果桉(Eucalyptusnitens)地上BCEF与林分年龄呈现出十分显著的负相关关系。现有学者主要利用回归模型、方差分析以及相关性分析等统计学方法,对BCEF的影响因素进行了研究[5, 18- 20,22]。如左舒翟等[5]利用回归模型方法分析了林分特征因子对杉木(Cunninghamialanceolata)地上BCEF的影响;罗云建等[19]利用回归模型、相关性分析方法研究了林分特征因子对落叶松(Larixgmelinii)地上BCEF的影响;吴小山[22]利用方差分析方法研究了地形、土壤和人为因子对杨树(Populus)地上BCEF的影响。然而,关于林分特征因子与环境因子如何协同影响BCEF的以及对BCEF影响的相对贡献率是怎样的,目前尚未见报道。
马尾松(Pinusmassoniana)在造林、用材等方面都有着较为广泛的应用[23]。由第八次国家森林资源清查可知,马尾松总面积和总蓄积高达1.0×107hm2和5.9×108m3,分别占全国乔木林总面积和总蓄积的6.1%和4.0%。福建省森林覆盖率为65.95%,其中福建省马尾松面积占福建省乔木林总面积的比例为13.79%,福建省马尾松蓄积占福建省乔木林总蓄积的比例为11.88%。在我国,森林资源调查的主要内容由立地与土壤、森林特征、森林功能、土地利用与覆盖以及其他因素等5大方面,其中,林分特征因子、土壤因子以及地形因子这3类因子不仅与林木的生长息息相关,而且相对容易获取。增强回归树(BRT)是一种利用随机抽样和自学习方法而建立的多重回归树[24],BRT方法提高了计算结果的稳定性和精度,可获得自变量对因变量影响的相对贡献率,已经在分析土地利用的影响因素[25]、区域生物量的影响因素[26]以及火灾的影响因素[27]等研究中得到成功应用。本文以第8次国家森林资源清查福建省优势树种为马尾松的固定样地数据作为研究材料,将BRT方法和森林资源调查数据相结合,分别确定地上BCEF以及地下BCEF的影响因素及其相对贡献率,以便提高林分生物量估算的准确性。
1 研究区概况与数据
福建省位于23°33′—28°20′N,115°50′—120°40′E之间,属亚热带海洋性季风气候,以丘陵和山地为主,广泛分布着黄壤、红壤以及砖红壤,最高海拔达2158米。 森林资源主要分布于龙岩、三明以及南平3个区市。马尾松、杉木以及桉树(Eucalyptus)是福建省分布最为广泛的3个乔木树种,其面积占福建乔木林总面积的比例分别为13.79%、22.36%以及4.47%;天然林中,马尾松、针叶混交林以及阔叶混交林占天然林面积比重相对较大,分别为9.44%、12.81%和57.27%;人工林中,桉树、马尾松林以及杉木占人工林面积比重相对较大,分别为9.77%、18.95%和40.35%。
福建省采用系统抽样方法、按4km×6km公里网格布设方形固定样地,共5059块。样地面积为0.0667hm2。从5059块样地中筛选出优势树种为马尾松的样地331块,然后对筛选的样地做进一步的处理:去除其它伴生树种以及检尺类型分别为多测木、枯倒木、采伐木以及枯立木的样木,只保留马尾松活立木。样地的调查因子包括:起源、龄组、地貌、海拔、坡向、坡位、坡度、土壤种类、土壤厚度和腐殖层厚度等。上述各因子的确定与划分标准参照国家森林资源清查技术规定。
2 方法
2.1 单木生物量和材积计算
本研究采用生物量模型和材积模型计算相应的生物量和材积,模型形式以及参数选用中华人民共和国林业行业标准《LY/T 2263—2014:立木生物量模型及碳计量参数——马尾松》中总体一的相关模型公式,该标准中依据总体一所建立的相关模型适用于福建省。上述标准的建模数据在选取过程中,充分考虑了在立地条件、起源以及龄组等方面具有广泛代表性的样本,建模数据调查取样的方法详见曾伟生[28]一文。由于国家森林资源清查只对胸径大于等于5cm的样木进行了每木检尺,而未进行每木测高[29- 31],故本研究选用以胸径(胸径≥5cm)为唯一自变量的一元模型计算各单株木的生物量和材积。表1为具体的模型形式。

表1 生物量模型和材积模型的模型形式以及评价指标
D:胸径(cm且≥5cm)
2.2 样地水平BCEF计算
BCEF可以将蓄积转换并扩展为生物量,单位为t/m3。在本研究中,依据下述公式计算样地水平各项BCEF[18]:

(1)
式中,i表示林木第i个分项(如:地上部分、地下部分);k表示第k个样地;j表示第j株树;Nk表示第k个样地中树木的总株树;BCEFik表示第k个样地、分项i的样地水平生物量转换和扩展因子;Bij表示第k个样地、第j株树、分项i的生物量;Vj表示第k个样地、第j株树的材积。
各项BCEF的计算结果以及部分土壤、地形因子的统计见表2;部分土壤、地形和林分特征因子各级(类)别样地数的统计见表3。
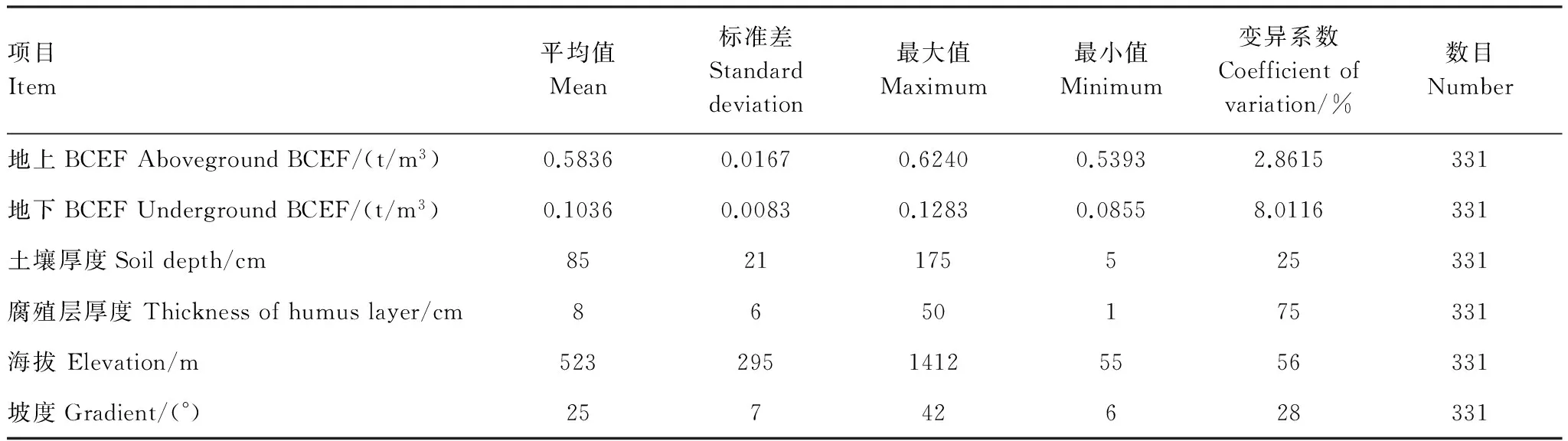
表2 各项BCEF以及部分土壤、地形因子的统计
表3部分土壤、地形、林分特征因子样地数的统计
Table3Statisticsofplotnumberforsomesoilfactors,topographyfactorsandstandcharacteristicfactors
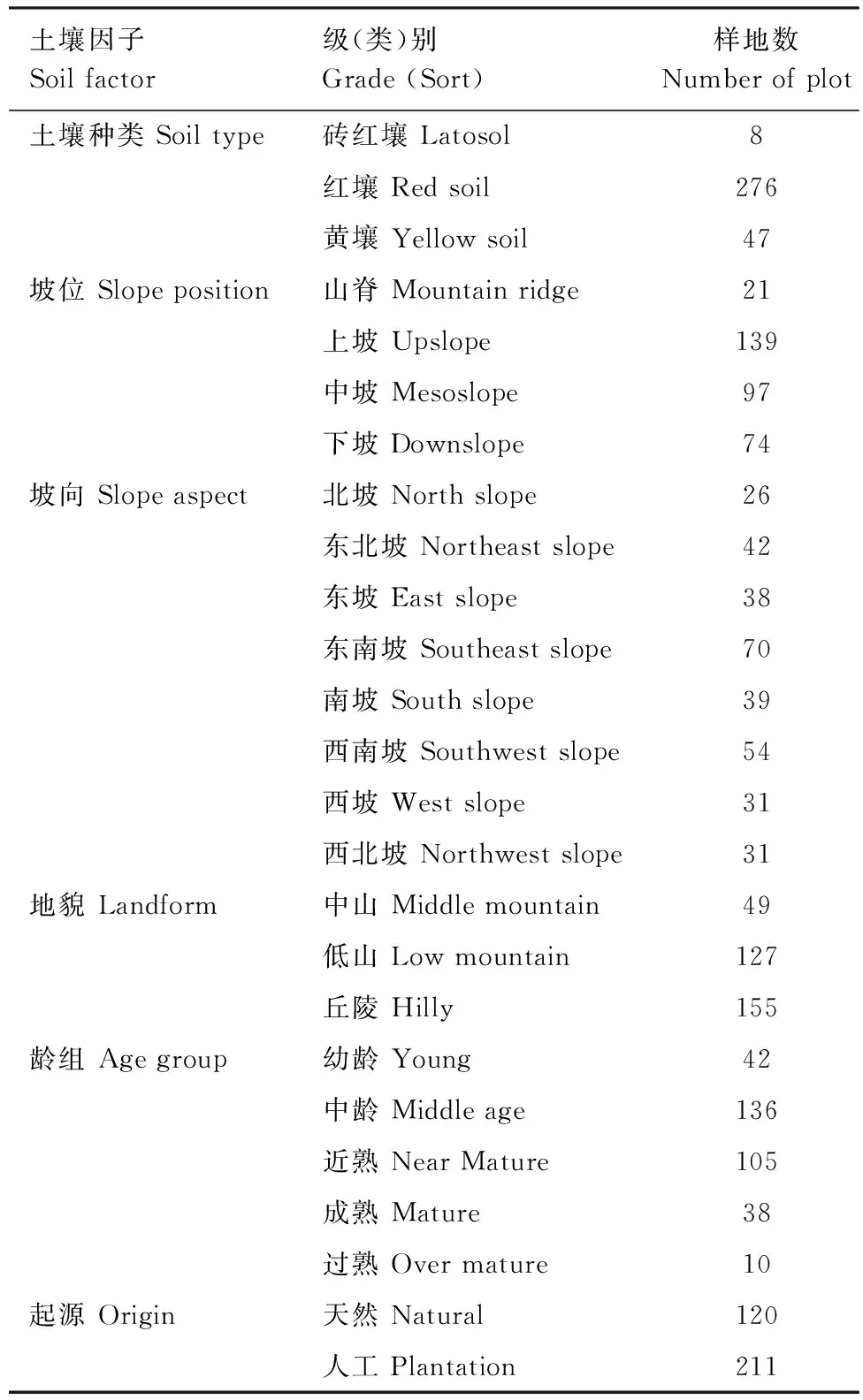
土壤因子Soilfactor级(类)别Grade(Sort)样地数Numberofplot土壤种类Soiltype砖红壤Latosol8红壤Redsoil276黄壤Yellowsoil47坡位Slopeposition山脊Mountainridge21上坡Upslope139中坡Mesoslope97下坡Downslope74坡向Slopeaspect北坡Northslope26东北坡Northeastslope42东坡Eastslope38东南坡Southeastslope70南坡Southslope39西南坡Southwestslope54西坡Westslope31西北坡Northwestslope31地貌Landform中山Middlemountain49低山Lowmountain127丘陵Hilly155龄组Agegroup幼龄Young42中龄Middleage136近熟NearMature105成熟Mature38过熟Overmature10起源Origin天然Natural120人工Plantation211
2.3 BRT法分析主要调查因子对BCEF的影响
BRT法是一种基于分类和回归树的、高级形式的机器学习方法[32]。BRT法利用训练数据分析各个自变量对因变量的影响、利用测试数据检验拟合结果,最终获得多重回归结果的平均值[27,33]。BRT法不仅可获得自变量对因变量影响的相对贡献率,而且还可以获得在其它自变量不变或取均值的前提下,因变量随某一自变量变化而变化的边际效应[27]。
BRT模型有4个重要的输入参数需要设置:损失函数的形式(distribution),学习速率(learning.rate),再抽样比率(bag.fraction),交叉验证折数(cv.folds)[24,32,34]。损失函数的形式容易设定,分类问题选择bernoulli分布,回归问题选择gaussian分布[24]。学习速率决定模型达到最优时所需的训练时间,学习速率值过小,则收敛速度慢、 训练时间长;学习速率值过大,则容易在抽样时产生过多噪音以及使得响应函数平滑性降低、稳定性差[34]。交叉验证折数是指将全部数据随机k等分,k-1部分作为训练数据,剩下的部分作为测试数据。再抽样比率是实际用于BRT模型建模的数据占交叉验证中训练数据的比例,从训练数据中再抽样不仅减少了运算量,而且在一定程度上避免了过度学习。依据De′Ath[32]和Elith等[24]推荐的经验法则,并结合本研究的情况,BRT模型参数设置为:①损失函数的形式为“gaussian”;②学习速率为0.005;③交叉验证折数为8;④再抽样比率为0.5。本文在统计软件R 3.3.0下调用“gbm”包进行BRT统计分析,此外,使用到的软件还有Excel 2013。
通过使用BRT法,可以量化林分特征因子(起源和龄组)、地形因子(地貌、海拔、坡向、坡位和坡度)以及土壤因子(土壤种类、土壤厚度和腐殖层厚度)分别对地上BCEF以及地下BCEF影响的相对贡献率,并获取相应的边际效应。
3 结果分析
3.1 多重因素分别对地上BCEF和地下BCEF影响的相对贡献率
由图1A可以看出,龄组对地上BCEF的影响最大,相对贡献率为41.13%,其次为坡向(14.52%)和海拔(10.57%),土类最小(0.18%);由图1B可以看出,龄组对地下BCEF的影响最大(41.54%),坡向(15.16%)和腐殖层厚度(9.13%)次之,土类最小(0.37%)。值得一提的是,海拔对地上BCEF的影响力(10.57%)明显大于地下BCEF(8.87%),这可能是由于马尾松地上部分和地下部分适应生境变化的能力不同引起的。本文中马尾松海拔差异较大(55—1412 m,表2),海拔不同,导致热、水、光和气等的综合效应产生差异[35- 36],植物的生境发生变化。
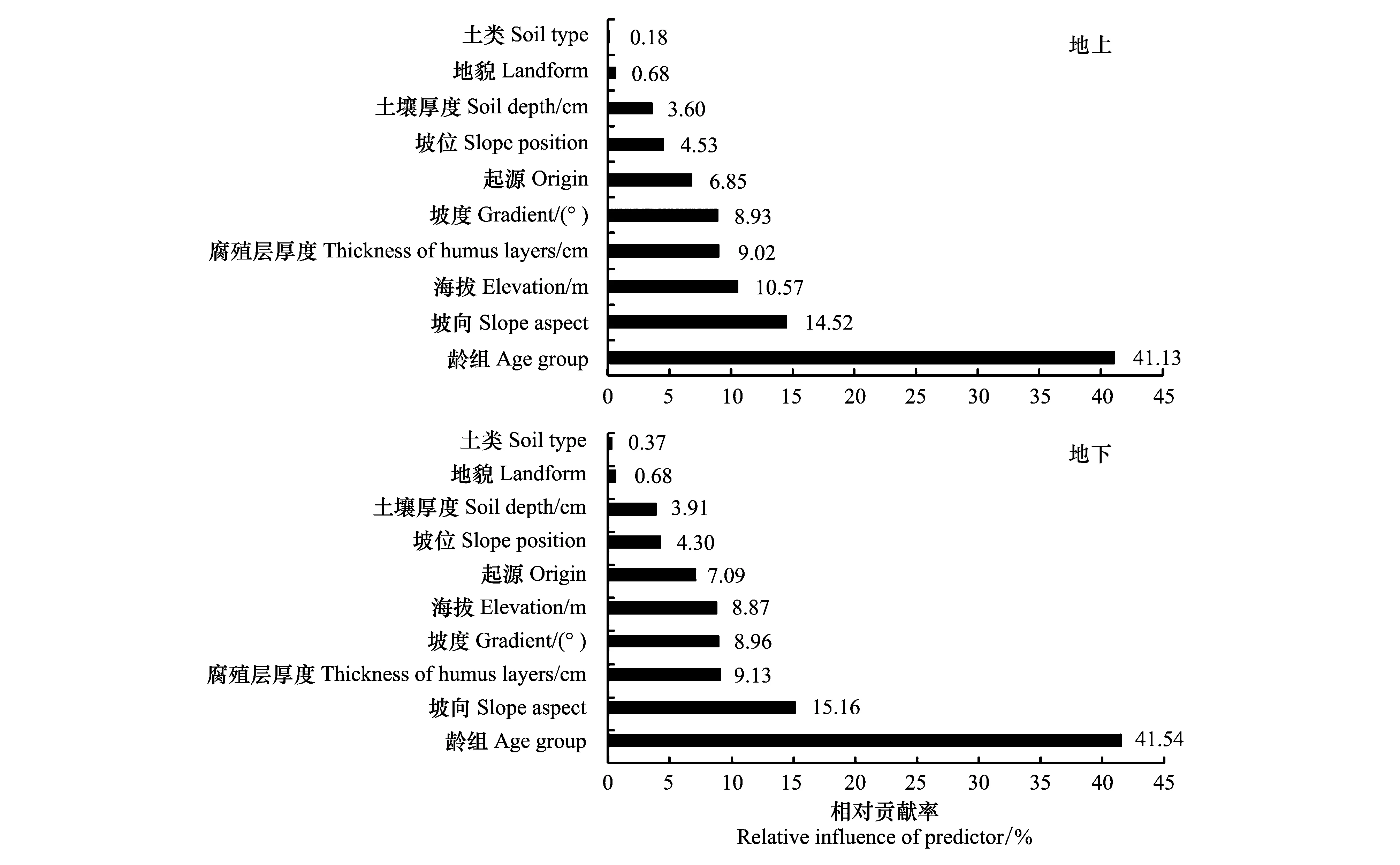
图1 各自变量分别对地上BCEF和地下BCEF影响的相对贡献率Fig.1 Relative influences of predictor variables affecting aboveground BCEF and underground BCEF
3.2 多重因素对地上BCEF影响的边际效应
林分特征因子对地上BCEF影响的相对贡献率为47.98%,其中,龄组41.13%,起源6.85%(图1)。地上BCEF随龄组由幼龄向过熟变化而减小,幼龄林最大、过熟林最小(图2)。此外,从图2可以看出,人工林地上BCEF显著大于天然林,与之相似的结果也出现在其它森林类型的研究中,罗云建等[37]发现落叶松人工林地上BCEF显著大于天然林,罗云建等[37]认为光照是引起差异的主要原因,一般而言,天然林通常为异林龄,人工林通常为同龄林[38],两者的林分结构存在差异,受光不同,光照调控植物不同器官的生长,进而影响着树木的材积和生物量。
地形因子对地上BCEF影响的相对贡献率为39.23%,其中,坡向最大(14.52%),其次为海拔(10.57%)、坡度(8.93%)、坡位(4.53%),地貌最小(0.68%)(图1)。对于坡向,地上BCEF在东南坡最大,在西坡最小(图2)。对于海拔,地上BCEF随海拔的升高而升高,并在海拔大于1000m后趋向平稳(图2)。一般海拔每升高100m气温下降0.5—0.6 ℃[39],随海拔上升,环境条件逐渐趋于恶劣[35],植物的地上部分通过加强同化能力、积累有机物来适应生境变化,故地上BCEF随着海拔的升高而升高;一般而言,海拔1000—1200m是马尾松分布的低温极限,然而,少数福建省马尾松林可以分布在海拔1100m以上[39],极端的生境条件下植物生长受限可能是地上BCEF在海拔大于1000m后趋向平稳的原因。此外,对于坡度,当坡度小于30°时,地上BCEF相对稳定,当处于30°—35°之间时,地上BCEF先升高后下降,大于35°后,地上BCEF不再变化;对于坡位,地上BCEF在上坡处最大、中坡处最小;对于地貌,丘陵处的地上BCEF较中山和低山的大(图2)。
土壤因子对地上BCEF影响的相对贡献率为12.80%,其中,腐殖层厚度最大(9.02%),土壤厚度次之(3.60%),而土壤种类最小,仅为0.68%(图1)。从图2可知,当腐殖层厚度小于10cm时,地上BCEF随腐殖层厚度的增加而减小,超过10cm后,地上BCEF不再变化并趋于平稳;当土壤厚度小于50cm时,地上BCEF相对稳定,当介于50—110cm之间时,地上BCEF先升高后下降,大于110cm后,地上BCEF不再变化;土壤种类对地上BCEF无影响。
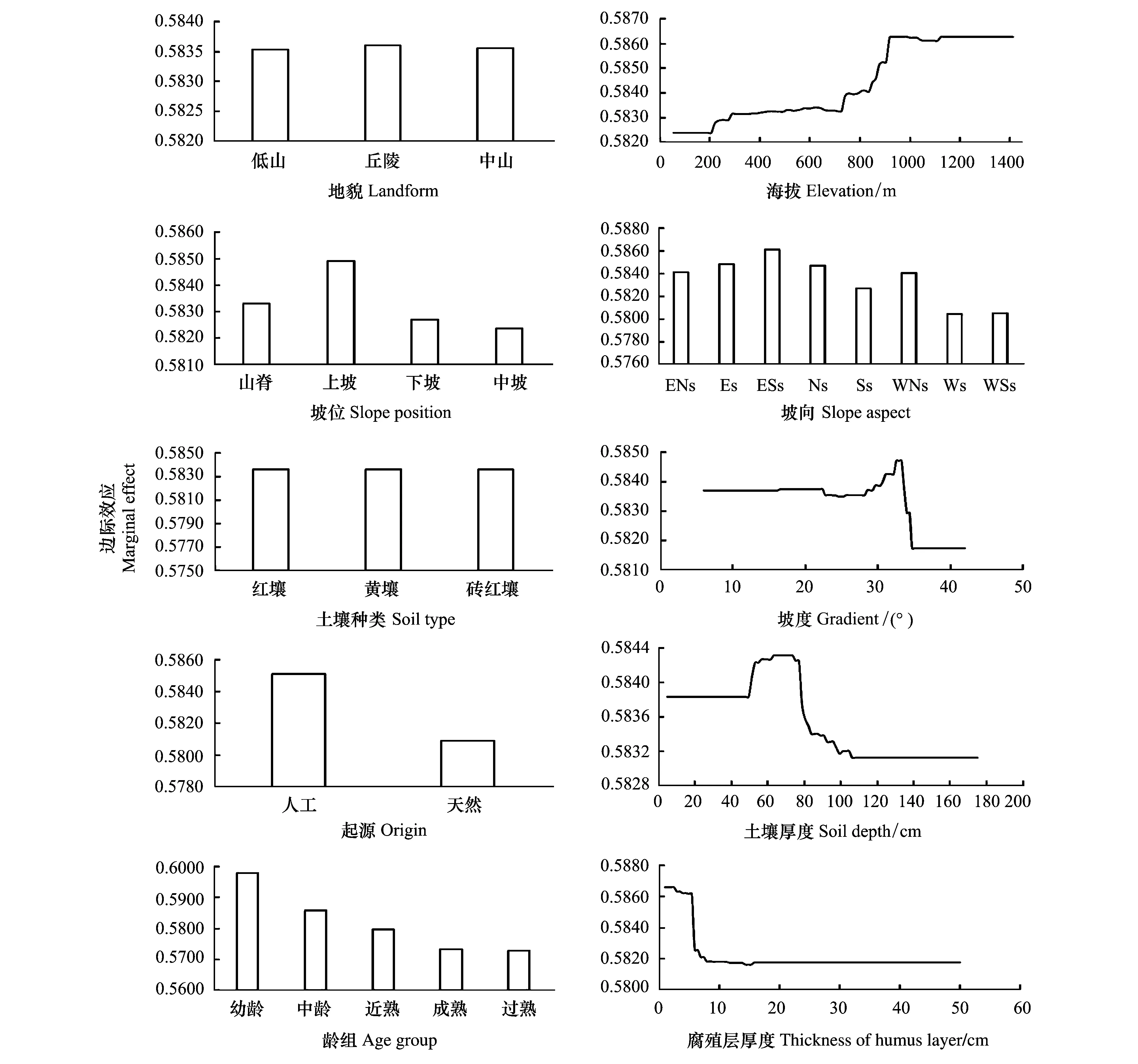
图2 各自变量对地上BCEF影响的边际效应Fig.2 Marginal effect of predictor variables on aboveground BCEF坡向:ENs(东北坡),Es(东坡),ESs(东南坡),Ns(北坡),Ss(南坡),WNs(西北坡),Ws(西坡),WSs(西南坡)
3.3 多重因素对地下BCEF影响的边际效应
林分特征因子对地下BCEF影响的相对贡献率为48.63%,其中,龄组和起源分别为41.54%和7.09%(图1)。地下BCEF随龄组由幼龄向过熟变化而增大,幼龄林最小、过熟林最大(图3)。对于起源,人工林地下BCEF明显小于天然林地下BCEF(图3),地下BCEF随起源而变化的趋势与地上BCEF相反,这可能是因为不同起源的植物受光不同,进而调控有机物向地上部分和地下部分分配的策略不同。
地形因子对地下BCEF影响的相对贡献率为37.97%,其中,坡向最大(15.16%),坡度(8.96%)、海拔(8.87%)、坡位(4.30%)次之,地貌最小(0.68%)(图1)。从图3可知,对于坡向,地下BCEF在东南坡最小,在西坡最大;对于海拔,地下BCEF随着海拔的升高而减小,在海拔大于1000m后逐渐趋向平稳;对于坡度,当坡度小于30°时,地下BCEF相对稳定,处于30°—35°之间时,地下BCEF先下降后升高,大于35°后,地下BCEF不再变化;对于坡位,地下BCEF在上坡处最小、中坡处最大;对于地貌,丘陵处的地下BCEF较中山和地山的小。
土壤因子对地下BCEF影响的相对贡献率为13.41%,其中,腐殖层厚度最大(9.13%),土壤厚度次之(3.91%),土壤种类最小,仅为0.37%,(图1)。从图3可知,对于腐殖层,当腐殖层厚度小于10cm时,地下BCEF随腐殖层厚度的增加而增大,超过10cm后,地下BCEF不再变化并趋于平稳;对于土壤厚度,当土壤厚度小于50cm时,地下BCEF一直处于稳定状态,介于50—100cm之间时,地下BCEF先下降后升高,大于100cm后,地下BCEF不再变化;对于土壤种类,红壤处的地下BCEF最大,黄壤处的地下BCEF最小。
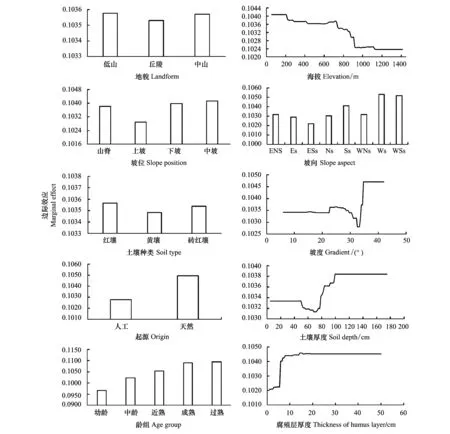
图3 各自变量对地下BCEF影响的边际效应Fig.3 Marginal effect of predictor variables on underground BCEF
4 结论与讨论
利用森林资源调查数据,探究林分特征因子、地形因子以及土壤因子如何协同影响林分BCEF以及多重因素对林分BCEF影响的相对贡献率,可为森林生物量和碳储量的准确估算提供科学根据[14,26]。本文研究结果表明,林分特征因子和地形因子是影响地上BCEF以及地下BCEF的主导因素,二者对地上BCEF影响的相对贡献率之和为87.20%、地下BCEF为86.59%。其中,龄组是林分特征因子中影响地上BCEF以及地下BCEF的最大因素,也是所有影响因素中对地上BCEF以及地下BCEF影响相对贡献率最大的因素(41.13%和41.54%)(图1);坡向是地形因子中影响地上BCEF以及地下BCEF的最大因素(14.52%和15.16%)(图1)。土壤因子对地上BCEF以及地下BCEF的影响都较小(12.80%和13.41%),腐殖层厚度是土壤因子中影响地上BCEF以及地下BCEF的最大因素(9.02%和9.13%)。
植物的生长与环境密切相关,面对复杂、多变的环境,植物必须不断地调节自身内部机能,以实现对生境的适应[40]。地下部分为地上部分提供水、矿质元素,而地上部分为地下部分提供有机物[41],当植物所处的生境发生变化时,调控有机物向地上部分和地下部分分配的策略也会不同。本文发现,当林分的龄组依次从幼龄向过熟变化时,地上BCEF逐渐减小,而地下BCEF的变化趋势恰好相反。地上BCEF随着林龄的增加而减小,这与罗云建等[19]、Pajtík等[42]、González-García等[20]的研究结果一致;地下BCEF随着林龄的增加而增大,这与Lehtonen等[18]的研究结果一致。林木各器官生物量的累积随着林龄的变化而发生变化[26],当林分处于中幼龄阶段时,林木间的相互竞争以及林分郁闭,需要更多的光合作用产生的有机物以促进林分的快速生长,此时林分地上部分生长迅速,地下部分的生长受到一定的限制[43]。当林分处于成过熟阶段时,林分地上部分的细胞分化成熟、不再具有分裂能力[41],地上部分的生长就生长放缓或停止;而植物的地下部分——根端具有特殊的顶端分生组织[41],可以无限分裂,故地下部分可以继续生长。地上BCEF在东南坡最大、西坡最小,这不同于罗云建[19]、吴小山[22]的研究结果,罗云建[19]、吴小山[22]发现不同坡向间的地上BCEF无显著差异;地下BCEF在东南坡最小、西坡最大。坡向的差异会导致光照强度、水分、温度、土壤湿度等生态环境的变化,不同坡向间日照时长、生长季长度等因素也存在差异,从而对植物个体生长产生影响[44- 45]。
龄组对地上BCEF和地下BCEF的影响均极大,且当林分分别处于幼龄和成过熟阶段时,幼龄林的地上BCEF会远远大于成过熟林的地上BCEF,幼龄林的地下BCEF会小于成过熟林的地下BCEF。本文建议,在进行外业调查估算森林生物量和碳储量时,应按照各龄组的林分比例进行采样,依据龄组计算相应的BCEF或者建立林龄普适的BCEF模型。此外,在采样时,应综合考虑生物因素和非生物因素对林分的影响,尽可能选择林分条件、地形、土壤等方面具有广泛代表性以及多样性的样地,尽量避免只选生长良好、环境条件单一的林分[26]。本文仅研究了土壤因子、地形因子以及林分特征因子对各项BCEF的影响,然而林木生长还会受到诸如气候因子、火因子、生物因子以及人为因子的影响[26],有待深入研究。
[1] Keith H, Mackey B, Berry S, Lindenmayer D, Gibbons P. Estimating carbon carrying capacity in natural forest ecosystems across heterogeneous landscapes: addressing sources of error. Global Change Biology, 2010, 16(11): 2971- 2989.
[2] 雷相东, 张会儒, 牟惠生. 东北过伐林区蒙古栎林分相容性生物量模型研究. 第四纪研究, 2010, 30(3): 559- 565.
[3] Temesgen H, Affleck D, Poudel K, Gray A, Sessions J. A review of the challenges and opportunities in estimating above ground forest biomass using tree-level models. Scandinavian Journal of Forest Research, 2015, 30(4): 326- 335.
[4] Timmons D S, Buchholz T, Veeneman C H. Forest biomass energy: Assessing atmospheric carbon impacts by discounting future carbon flows. GCB Bioenergy, 2016, 8(3): 631- 643.
[5] 左舒翟, 任引, 王效科, 张小全, 罗云建. 中国杉木林生物量估算参数及其影响因素. 林业科学, 2014, 50(11): 1- 12.
[6] 罗云建, 张小全, 王效科, 朱建华, 侯振宏, 张治军. 森林生物量的估算方法及其研究进展. 林业科学, 2009, 45(8): 129- 134.
[7] Schroeder P, Brown S, Mo J M, Birdsey R, Cieszewski C. Biomass estimation for temperate broadleaf forests of the united states using inventory data. Forest Science, 1997, 43(3): 424- 434.
[8] Somogyi Z, Cienciala E, Mäkipää R, Muukkonen P, Lehtonen A, Weiss P. Indirect methods of large-scale forest biomass estimation. European Journal of Forest Research, 2007, 126(2): 197- 207.
[9] Tobin B, Nieuwenhuis M. Biomass expansion factors for Sitka spruce (Piceasitchensis(Bong.) Carr.) in Ireland. European Journal of Forest Research, 2007, 126(2): 189- 196.

[11] Magalhães T M, Seifert T. Tree component biomass expansion factors and root-to-shoot ratio of Lebombo ironwood: measurement uncertainty. Carbon Balance and Management, 2015, 10(1): 1- 14.
[12] Tolunay D. Coefficients that can be used to calculate biomass carbon storage of Turkish forests. Journal of the Faculty of Forestry Istanbul University, 2017, 67(2), doi: 10.17099/jffiu.xxxxx.
[13] Di Cosmo L, Gasparini P, Tabacchi G. A national-scale, stand-level model to predict total above-ground tree biomass from growing stock volume. Forest Ecology and Management, 2016, 361: 269- 276.
[14] Lam T Y, Li X D, Kim R H, Lee K H, Son Y M. Bayesian meta-analysis of regional biomass factors forQuercusmongolicaforests in South Korea. Journal of Forestry Research, 2015, 26(4): 875- 885.
[15] Soares P, Tomé M. Analysis of the effectiveness of biomass expansion factors to estimate stand biomass//Proceedings of the International Conference on Modeling Forest Production. Austria, 2004.
[16] 方精云, 刘国华, 徐嵩龄. 我国森林植被的生物量和净生产量. 生态学报, 1996, 16(5): 497- 508.
[17] 刘国华, 傅伯杰, 方精云. 中国森林碳动态及其对全球碳平衡的贡献. 生态学报, 2000, 20(5): 733- 740.
[18] Lehtonen A, Mäkipää R, Heikkinen J, Sievänen R, Liski J. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests. Forest Ecology and Management, 2004, 188(1/3): 211- 224.
[19] 罗云建. 华北落叶松人工林生物量碳计量参数研究[D]. 北京: 中国林业科学研究院, 2007.
[20] González-García M, Hevia A, Majada J, Barrio-Anta M. Above-ground biomass estimation at tree and stand level for short rotation plantations ofEucalyptusnitens(Deane & Maiden) Maiden in Northwest Spain. Biomass and Bioenergy, 2013, 54: 147- 157.
[21] Teobaldelli M, Somogyi Z, Migliavacca M, Usoltsev V A. Generalized functions of biomass expansion factors for conifers and broadleaved by stand age, growing stock and site index. Forest Ecology and Management, 2009, 257(3): 1004- 1013.
[22] 吴小山. 杨树人工林生物量碳计量参数研究[D]. 雅安: 四川农业大学, 2008.
[23] 黄云奉, 刘屹, 黄世友, 何邦亮, 陈丽洁, 李燕琼. 不同林龄马尾松生长及生物量分配研究. 四川林业科技, 2015, 36(4): 72- 75.
[24] Elith J, Leathwick J R, Hastie T. A working guide to boosted regression trees. Journal of Animal Ecology, 2008, 77(4): 802- 813.
[25] Weisberg P J, Shandra O, Becker M E. Landscape influences on recent timberline shifts in the Carpathian Mountains: Abiotic influences modulate effects of land-use change. Arctic, Antarctic, and Alpine Research, 2013, 45(3): 404- 414.
[26] 闫晶, 罗云建, 郑德福, 王水城. 桉树生物量估算差异的源解析. 林业科学, 2014, 50(2): 92- 98.
[27] 焦琳琳, 常禹, 申丹, 胡远满, 李春林, 马俊. 利用增强回归树分析中国野火空间分布格局的影响因素. 生态学杂志, 2015, 34(8): 2288- 2296.
[28] 曾伟生. 全国立木生物量方程建模方法研究[D]. 北京: 中国林业科学研究院, 2011.
[29] Li H K, Zhao P X. Improving the accuracy of tree-level aboveground biomass equations with height classification at a large regional scale. Forest Ecology and Management, 2013, 289: 153- 163.
[30] 傅煜, 雷渊才, 曾伟生. 区域尺度杉木生物量估计的不确定性度量. 林业科学, 2014, 50(12): 79- 86.
[31] 傅煜, 雷渊才, 曾伟生. 单木生物量模型估计区域尺度生物量的不确定性. 生态学报, 2015, 35(23): 7738- 7747.
[32] De′Ath G. Boosted trees for ecological modeling and prediction. Ecology, 2007, 88(1): 243- 251.
[33] 李春林, 刘淼, 胡远满, 徐岩岩, 孙凤云. 基于增强回归树和Logistic回归的城市扩展驱动力分析. 生态学报, 2014, 34(3): 727- 737.
[34] Zhang Y, Chen H Y H, Reich P B. Forest productivity increases with evenness, species richness and trait variation: a global meta-analysis. Journal of Ecology, 2012, 100(3): 742- 749.
[35] 刘彦春, 张远东, 刘世荣, 张笑鹤. 川西亚高山针阔混交林乔木层生物量、生产力随海拔梯度的变化. 生态学报, 2010, 30(21): 5810- 5820.
[36] 吴栋栋, 周永斌, 于大炮, 戴冠华. 不同海拔长白山岳桦的生理变化. 生态学报, 2009, 29(5): 2279- 2285.
[37] 罗云建, 张小全, 侯振宏, 于彭涛, 朱建华. 我国落叶松林生物量碳计量参数的初步研究. 植物生态学报, 2007, 31(6): 1111- 1118.
[38] 罗云建, 王效科, 张小全, 朱建华, 张治军, 侯振宏. 华北落叶松人工林的生物量估算参数. 林业科学, 2010, 46(2): 6- 11.
[39] 张大鹏. 不同海拔高度对马尾松茎木质部结构的影响. 生态学杂志, 1994, 13(2): 12- 18.
[40] 姜汉侨, 段昌群, 杨树华, 王崇云, 苏文华. 植物生态学. 北京: 高等教育出版社, 2010.
[41] 胡金良. 植物学. 北京: 中国农业大学出版社, 2012.
[42] Pajtík J, Konpka B, Lukac M. Individual biomass factors for beech, oak and pine in Slovakia: a comparative study in young naturally regenerated stands. Trees, 2011, 25(2): 277- 288.
[43] Vanninen P, Ylitalo H, Sievänen R, Mäkelä A. Effects of age and site quality on the distribution of biomass in Scots pine (PinussylvestrisL.). Trees, 1996, 10(4): 231- 238.
[44] 郭滨德, 张远东, 王晓春. 川西高原不同坡向云、 冷杉树轮对快速升温的响应差异. 应用生态学报, 2016, 27(2): 354- 364.
[45] 王富刚, 徐伟洲, 亢福仁, 尚爱军, 张静. 不同坡向条件下小叶杨光合光响应曲线研究. 水土保持研究, 2015, 22(6): 177- 182.
[46] 李俊清. 森林生态学(第二版). 北京: 高等教育出版社, 2010.
FactorsaffectingthebiomassconversionandexpansionfactorofmassonpineinFujianProvince
OU Qiangxin1, LI Haikui1, *, YANG Ying2
1ResearchInstituteofForestResourceInformationTechniques,ChineseAcademyofForestry,Beijing100091,China2AcademyofForestryInventoryandPlanning,StateForestryAdministration,Beijing100714,China
国家自然科学基金 (31370634);中国清洁发展机制基金赠款项目:2020年后林业增汇减排的行动目标研究(2013014)
2016- 06- 13; < class="emphasis_bold">网络出版日期
日期:2017- 04- 24
10.5846/stxb201606131140
*通讯作者Corresponding author.E-mail: lihk@ifrit.ac.cn
欧强新,李海奎,杨英.福建地区马尾松生物量转换和扩展因子的影响因素.生态学报,2017,37(17):5756- 5764.
Ou Q X, Li H K, Yang Y.Factors affecting the biomass conversion and expansion factor of masson pine in Fujian Province.Acta Ecologica Sinica,2017,37(17):5756- 5764.
猜你喜欢
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年5期)2019-11-29 13:17:37
测绘学报(2019年11期)2019-11-20 01:31:42
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
广西林业科学(2016年1期)2016-03-20 05:33:00
广西林业科学(2016年1期)2016-03-20 05:32:59
广西林业科学(2016年1期)2016-03-20 05:32:58
土壤与作物(2015年3期)2015-12-08 00:46:55
亚热带资源与环境学报(2015年3期)2015-01-22 07:05:36
