
长白山垂直带森林叶片-凋落物-土壤连续体有机碳动态
——基于稳定性碳同位素分析
2017-10-13 11:29:07司高月李晓玉程淑兰方华军于贵瑞于光夏
生态学报 2017年16期
司高月,李晓玉,程淑兰,*,方华军,,于贵瑞,耿 静,何 舜,于光夏
1 中国科学院大学资源与环境学院,北京 100049 2 中国科学院地理科学与资源研究所/生态系统观测与模拟重点实验室,北京 100101
长白山垂直带森林叶片-凋落物-土壤连续体有机碳动态
——基于稳定性碳同位素分析
司高月1,李晓玉2,程淑兰1,*,方华军1,2,于贵瑞2,耿 静2,何 舜1,于光夏1
1 中国科学院大学资源与环境学院,北京 100049 2 中国科学院地理科学与资源研究所/生态系统观测与模拟重点实验室,北京 100101
稳定性碳同位素自然丰度(δ13C)记录着生态系统碳循环过程的关键信息,常被用于评价全球变化情景下陆地生态系统碳的动态。以长白山北坡垂直带4种典型森林生态系统为研究对象,测定乔木建群种叶片、凋落物以及不同深度土壤有机碳(SOC)含量和δ13C值,探讨植物叶片-凋落物-土壤连续体碳含量、δ13C丰度的分布格局及其生态学暗示。研究结果表明:植物叶片碳含量随海拔高度的增加呈现抛物线型变化,且阔叶树叶片碳含量显著低于针叶树,体现气候要素和植被功能型的支配作用,并且暗示针叶树种潜在的碳蓄积能力更强。此外,植物叶片δ13C随海拔高度升高而降低,表明高海拔植物叶片水分利用效率较低,即固碳耗水成本更高。凋落物碳含量随海拔增加逐渐下降,而矿质表层土壤则表现为阔叶红松林、岳桦林显著高于暗针叶林,体现了植被类型和土壤质地的共同支配作用。总体上,岳桦林SOC周转最快,其次是暗针叶林,位于基带的阔叶红松林最慢。可见,小尺度上气候因子并不是温带森林地下碳循环的主导因素,植被功能型和土壤属性对SOC周转与稳定的影响更大。在探讨环境因子对陆地生态系统碳循环和碳平衡的影响时需要考虑研究尺度,不同的研究尺度影响SOC周转的驱动因子并不相同。研究方法方面,基于logSOC和δ13C的SOM周转模型能够很好地概括不同生态系统类型下SOM周转的相对快慢,可用来评价SOC动态对全球变化的响应。
13C自然丰度;叶片-凋落物-土壤连续体;水分利用效率;土壤碳周转;山地垂直带
Abstract: The natural abundance of the stable carbon isotope (δ13C) records key information regarding the ecosystem carbon (C) cycle and is commonly used to assess the C dynamics in terrestrial ecosystems under global change. In this study, we selected four typical forest ecosystems along the vertical transect distributed in Changbai Mountain and measured the C concentrations andδ13C values of leaves of constructive tree species, litter, and soils at different soil layers. The aim of this study was to explore the patterns of C content andδ13C values in the leaf-litter-soil continuum, as well as their ecological indications. The results showed that foliar C content first increased and then decreased with the increasing altitude, and the parabolic peak appeared at the Ermans birch-spruce-fir forest stand; moreover, the C content of broadleaved tree species was significantly lower than that of coniferous species, reflecting that coniferous species had a higher C sequestration capacity relative to that of broadleaved species. Climatic factors and vegetation types dominated the pattern of foliar C content. In addition, foliarδ13C decreased with increasing altitude, indicating that vegetation at high-altitude sites had lower water use efficiency (WUE) and higher water consumption by C sequestration relative to that at low altitude sites. Litter C content gradually decreased with increase in altitude, whereas topsoil C content at the 0—20 cm depth at the broad-leaved Korean pine forest (BLKP) and Ermans birch forest (EB) was higher than that of the Korean pine-spruce-fir forest (KPSF) and Ermans birch-spruce-fir forest (EBSF), reflecting the predominance of vegetation type and soil texture together. Overall, the birch forest had the highest SOC turnover rate, followed by that of the two dark coniferous forests, and that of the broad-leaved Korean pine forest was the lowest. Our results suggest that climatic factors are not the predominant factors in the belowground C cycle of temperate forests at a small scale, and vegetation functional types and soil properties could have greater effects on the turnover and stability of SOC. Because the factors driving the turnover of SOC are not the same at different study scales, we should more intensively consider the research scale when we explore the effects of environmental factors on C cycle and C budget in terrestrial ecosystems. The SOM turnover model, based on the regression of logSOC andδ13C, is a good method to characterize the rate of SOM turnover in various ecosystems, which can be used to evaluate the response of SOC dynamics to global change.
KeyWords:13C natural abundance; leaf-litter-soil continuum; water use efficiency; soil C turnover; mountain transect
森林作为陆地生态系统的重要组成部分,是大气温室气体的源和汇,也是陆地生态系统最大的碳库[1]。全球约有1400—1500 Pg (1Pg=1015g) 碳以有机态形式储存于土壤中,是陆地植被碳库的2—3倍,而森林土壤约占全球土壤有机碳(SOC)库的73%[2]。其中,温带森林面积为10.38亿hm2,植被和土壤碳储量分别为59Pg和100Pg,占全球森林总碳储量的10.9%,在森林生态系统碳循环和碳平衡中具有举足轻重的地位[3]。SOC储量受多种因素影响,包括气温、降水[3]、森林类型[4-5]、土壤理化性质[6]、干扰及人为管理等[7]。准确评价环境因子对森林生态系统碳储量和碳周转的影响,是构建碳素生物地球化学过程模型的前提和基础,也是有效降低区域和全球陆地生态系统碳平衡评估不确定性的关键。
植物-凋落物-土壤连续体稳定性碳同位素自然丰度(δ13C)记录了生态系统碳循环过程的关键信息[8]。植物通过光合作用固定大气CO2(δ13C=-8‰),一部分用于构建自身的生物量(δ13C=-25—-30‰),很大一部分通过植物自养呼吸返回大气,还有一部分碳通过植物根系、凋落物(δ13C=-26‰)的形式进入土壤[9]。凋落物和其它动植物残体在微生物作用下逐步降解成小分子化合物,然后再缩合成腐殖质(δ13C=-25‰),在此过程中释放CO2(δ13C=-27‰)到大气中。由于微生物选择性地利用轻的12C,导致残留反应物13C相对富集,而反应产物13C相对贫化[9]。因此,生态系统各组分的δ13C值差异明显,常被用于示踪研究气候变化和人为干扰情景下植被和土壤长期的碳动态。例如,植物叶片δ13C丰度与植物的水分利用效率(WUE)正相关,能够反映单位质量水分所能固定的碳量[10]。在土壤碳周转方面,粗颗粒、低密度SOM组分主要来源于最新进入、13C 贫化的凋落物残体,而细颗粒、高密度SOM组分则分解时间较长,13C相对富集[11]。因此,稳定性碳同位素丰度能够有效地表征凋落物和土壤碳储量的微小迁移与转化,以及定量评价新老SOC组分变化对碳储量的相对贡献[6,12]。然而,过去有关地下有机碳周转的研究多基于放射性的14C技术或13C标记方法[13-15],鲜有利用自然13C丰度来分析土壤碳含量动态和碳周转特征。
气候变化(如温度升高、降水格局改变等)会显著改变植被和土壤碳循环过程,进而影响生态系统的碳平衡[16]。野外控制实验、陆地生态系统样带和模型模拟等方法常被用于研究多个环境因子的交互作用对森林土壤碳循环的影响,3种方法各有所长,相辅相成互为补充[16]。山地垂直带谱具有天然的环境变化梯度,一般随着海拔高度的增加气温逐渐下降,而降水量会显著增加,为研究气候变化对生态系统碳氮循环过程的影响提供了理想的研究平台[17]。假设:随着海拔高度的增加,辐射增强,光合作用增加,植物碳含量增加;同时由于温度降低,降水增加,SOC周转会变慢,进而导致SOC积累。为了验证上述假设,本文以长白山北坡垂直带典型森林为研究对象,利用稳定性碳同位素技术,研究植物叶片-凋落物-土壤连续体碳含量和δ13C丰度的变化规律,分析SOC动态和δ13C丰度变化之间的关系,试图阐明不同气候带下碳的循环和周转特征。研究结果一定程度上可以加深森林生态系统碳循环过程及其驱动机制的理解。
1 材料与方法
1.1 研究区概况
研究区位于中国东北长白山自然保护区(127°38′E, 41°42′N),最大海拔高度2691 m。该区属于典型的温带大陆性山地气候,具有漫长寒冷的冬季和温暖的夏季。年均温为2.8—-7.3℃,年均降水量为679—1339 mm[18]。长白山北坡具有典型的垂直带谱,海拔500—1000 m为针阔混交林,1100—1700 m为暗针叶林,1700—2000 m为岳桦林,2000 m以上为高山苔原[18]。沿着长白山北坡山地垂直样带,我们选择了阔叶红松林(BLKP)、红松云冷杉林(KPSF)、岳桦云冷杉林(EFSF)和岳桦林(EB)等4个典型森林生态系统为研究对象。其中,阔叶红松林处于演替的顶级阶段,林龄约为200a;其他3个森林属于亚高山北方森林,均为老龄林。土壤类型分别是山地暗棕壤、棕色针叶林土和山地生草森林土。每个森林立地以及表层土壤属性如表1所示。

表1 长白山自然保护区垂直带4种典型森林立地和0—20 cm表层土壤属性
数据来源于中国生态系统研究网络(CERN)
1.2 植物与土壤采样
2015年7月中旬,在每个森林立地随机设置了3个独立的样方,面积10 m×10 m。在每个样方中每个建群种选择3株标准木,利用高枝剪采集冠层上方阳叶。阔叶红松林采集了红松、紫椴、水曲柳和蒙古栎4个乔木物种,红松云冷杉林采集了红松、鱼鳞云杉、冷杉3个物种,岳桦云冷杉林采集了鱼鳞云杉、冷杉、岳桦3个物种,岳桦林只采集了岳桦一个物种。除了水曲柳外,其他乔木树种都是外生菌根类型。样方内所选物种的总生物量占地上乔木生物量的80%以上[17],因此所选物种能够整体上代表该森林的植被。对于每个森林样方,用采样铲采集未分解的凋落物层(Oi)、半分解及完全分解的凋落物层(Oe+Oa)样品。通过挖掘土壤剖面来采集不同深度的矿质土壤样品,采样深度至母质层,每个样地挖掘一个土壤剖面,共12个;采样深度为40—50 cm,采样间隔为10 cm。采集的植物叶片先用水洗净表面浮尘,再用去离子冲洗干净,65℃下烘干。土壤样品自然风干,过2 mm筛。烘干的植物和风干的土壤样品利用球磨机粉碎过100目筛待测。
1.3 碳浓度与δ13C丰度的测定
利用元素分析仪(Flash EA1112, ThermoFinnigan, Milan, Italy)和稳定性同位素质谱仪(Finnigan MAT 253, Thermo Electron,Bremen, Germany)联机系统测定植物和土壤样品的碳浓度和δ13C丰度[10]。稳定性碳同位素丰度的计算公式为:

式中,δ13C为样品的13C丰度(‰),Rsample为样品的13C/12C比率,Rstandard为标准样品PDB的13C/12C比率。10份重复样品测定的标准差小于<0.4‰。
研究表明,影响土壤剖面δ13C分布格局的主要过程是碳的矿化,非生物扰动和溶解性有机碳(DOC)的淋溶影响较小[19]。因此,土壤δ13C值涵盖了新鲜凋落物向SOM转变过程时间信息[20]。利用δ13C与SOC含量对数的线性拟合方程(δ13C=a+K×logSOC)斜率(K)来描述凋落物向土壤有机质(SOM)转变引起的δ13C 变化,其内在的涵义表示碳含量每增加10倍引起的同位素分差[21]。K值的负值越大,表明微生物矿化引起的土壤碳周转越快[10,20-21]。
1.4 数据统计分析
利用双因素方差分析来检验森林立地和树种对叶片碳浓度和δ13C值的影响,利用单因素方差分析检验不同森林立地之间凋落物、矿质土壤有机碳含量及其δ13C值的差异,利用Tukey′s HSD进行均值比较。采用一元回归分析方法研究土壤δ13C和SOC含量自然对数值(logSOC)之间的关系,根据回归方程斜率值K来评价SOC周转随海拔高度的变化。利用SPSS 16.0软件进行数据统计分析,利用Sigmaplot 12.5软件进行统计绘图。
2 结果与分析
2.1 叶片碳含量和δ13C丰度
不考虑乔木类型,不同森林立地乔木叶片碳含量随着海拔高度的增加先增加后降低,呈现抛物线或二次方程变化,平均值变化范围在51.53%—56.70%,不同森林类型间差异显著,红松云冷杉林最高(图1,表2,P<0.001)。针叶树叶片碳含量均显著高于阔叶树,红松、云杉、鱼鳞冷杉最高,其次是紫椴和岳桦,蒙古栎和水曲柳叶碳含量最低(表2,P<0.001)。在同一森林立地中,阔叶红松林中红松叶片碳含量较其他阔叶树种高4.4%—10.6%,岳桦云冷杉林中针叶树种比岳桦叶片碳含量高1.7%—4.1%(图1)。
不考虑乔木类型,植物叶片δ13C值基本上随着海拔高度的增加逐渐下降(图1),但不同森林立地间差异不显著(表2,P=0.28)。就同一森林立地而言,针叶树叶片δ13C值显著低于阔叶树,阔叶红松林中红松以及岳桦云冷杉林中的云杉、冷杉叶片δ13C较阔叶树种低2.2%—5.4%。另外,乔木类型与森林立地之间具有明显的交互作用,随着海拔高度的增加,同一树种叶片δ13C有明显下降趋势(图1,表2,P=0.046)。
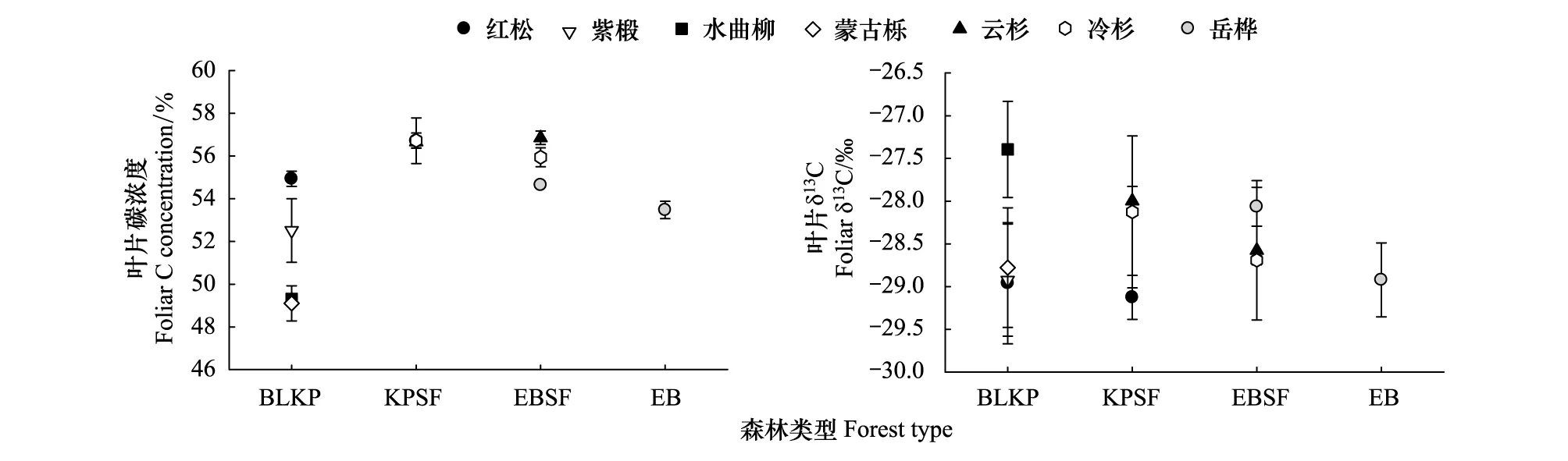
图1 不同森林不同乔木树种叶片碳含量和δ13C丰度的变化Fig.1 Variation of foliar C and δ13C among species and forests BLKP:阔叶红松林broad-leaved Korean pine forest,KPSF:红松云冷杉林Korean pine-spruce-fir forest,EBSF:岳桦云冷杉林Ermans birch-spruce-fir forest,EB:岳桦林Ermans birch forest

变异来源Sourceofvariation名称Name总碳含量TotalCcontent/%δ13C值δ13Cvalue/‰均值MeanP均值MeanP森林立地ForeststandsBLKP51.53c<0.001-28.34a0.28KPSF56.70a-28.42aEBSF55.81ab-28.44aEB53.47bc-28.92a树种类型Treespecies紫椴52.51b<0.001-28.92b0.039红松55.82a-29.04c水曲柳49.32c-27.39a蒙古栎49.36c-28.08b云杉56.75a-28.29b冷杉56.33a-28.41b岳桦54.06b-28.49b森林立地×树种类型Foreststands×treespecies0.490.046
BLKP:阔叶红松林broad-leaved Korean pine forest,KPSF:红松云冷杉林Korean pine-spruce-fir forest,EBSF:岳桦云冷杉林Ermans birch-spruce-fir forest,EB:岳桦林Ermans birch forest;同一列中不同字母表示均值的差异显著
2.2 土壤碳含量和δ13C丰度
4种森林土壤总碳含量随着土壤深度的增加呈指数下降,剖面变化格局相同,不同森林类型凋落物层和0—10 cm矿质表层土壤总碳含量差异显著(图2)。凋落物层碳含量随着海拔高度的增加逐渐下降,阔叶红松林(47.28%)显著高于岳桦林(33.84%)(表3)。矿质表层土壤碳含量也是阔叶红松林最高(5.79%),其次是高海拔的岳桦林(5.45%),而红松云冷杉林和岳桦云冷杉林最低(2.84%和3.02%)(表3)。
与土壤总有机碳变化趋势相反,土壤δ13C值随土壤深度的增加而增加(图2)。无论是凋落物层还是矿质土壤层,岳桦云冷杉林土壤δ13C值最高,岳桦林最低,不同林型间差异显著(表3)。就凋落物层而言,岳桦云冷杉林δ13C值平均为-27.15‰,分别比阔叶红松林、红松云冷杉林、岳桦林高2.5%、3.6%和5.6%(表3)。同样,岳桦云冷杉林矿质土壤层δ13C较其他3个森林分别高3.6%、3.1%和6.4%(表3)。
2.3 土壤碳含量与δ13C丰度之间的关系
长白山北坡垂直带4种森林土壤δ13C与logSOC均呈现极显著的负相关关系,logSOC能够解释土壤δ13C变异的95%以上(图3和表4)。回归方程斜率K的绝对值表示土壤SOC的周转速率,参数K的变化顺序依次为:岳桦林(2.92)>红松云冷杉林(1.85) >岳桦云冷杉林(0.97)>阔叶红松林(0.95)。研究结果表明,岳桦林土壤碳周转最快,而阔叶红松林土壤周转最慢。

图2 不同森林土壤有机碳含量和δ13C 丰度沿着土壤深度变化特征Fig.2 Variation of soil C contents and δ13C values among soil depths and forestsBLKP:阔叶红松林broad-leaved Korean pine forest,KPSF:红松云冷杉林Korean pine-spruce-fir forest,EBSF:岳桦云冷杉林Ermans birch-spruce-fir forest,EB:岳桦林Ermans birch forest;Oi: 未分解的凋落物层;Oa: 半分解的凋落物层;Oe:完全分解的凋落物层

土层Soilhorizons土壤属性Soilproperties森林类型ForesttypeBLKPKPSFEBSFEB凋落物层总碳/%47.28(1.33)a43.67(2.20)ab38.94(3.13)ab33.84(5.20)b有机层Ohorizonδ13C值/‰-27.82(0.18)b-28.14(0.32)bc-27.15(0.15)a-28.67(0.19)c矿质土壤层总碳/%5.79(1.67)a2.84(0.84)b3.02(0.98)ab5.45(0.68)aMineralsoillayerδ13C/‰-25.81(0.20)b-25.67(0.18)b-24.91(0.33)a-26.50(0.19)c
同一行中不同字母表示均值的差异显著(Turky′s HSD比较)

图3 4种森林土壤δ13C丰度与log SOC值之间回归关系Fig.3 Regression model between soil δ13C abundances and log SOC values in the four forests
3 讨论
3.1 植物叶片碳含量、δ13C丰度的分布格局及生态学意义
总体上,随着海拔高度的增加,植物叶片碳含量呈现先增加后降低的趋势(图1),部分证实了我们的研究假设。主要的原因是,随着海拔高度的增加光合有效辐射增加,气温下降,一定程度上增加了植物的光合碳固定,降低了植物线粒体的自养呼吸,导致植物体内的碳相对累积[22];但是,海拔1900 m左右的岳桦林叶片碳含量急剧下降,可能与低温、低大气CO2浓度以及云层遮挡等有关。此外,针叶树种叶片碳含量显著高于阔叶树种,与植物叶片的功能属性如比叶面积、叶寿命、叶氮浓度等密切相关。总体上,长寿命的针叶树种叶片叶氮浓度和光合能力均较低,必须更长时间进行光合作用以维持存在期间的碳平衡[22]。针叶树种叶片碳含量显著高于阔叶树种,说明针叶树种有机物含量较高,暗示其潜在的碳蓄积能力更强[23-24]。
在物种水平上,叶片δ13C可以反映植物光合作用过程中的水分利用效率(WUE),代表植物生长季期间植物与大气之间的碳水交换的综合效应[25]。水分利用效率的倒数即固碳耗水成本,即固定一定数量的碳所要消耗的水分[26]。不考虑乔木类型,植物叶片的δ13C随着海拔高度的增加而下降(图1),说明温度降低、降水增加会导致乔木建群种水分利用效率显著下降,暗示其固碳耗水成本将增加。另外,同一气候带,阔叶树种叶片的δ13C显著高于针叶树种,说明阔叶树种水分利用效率高于针叶树,其固碳耗水成本较低,与植物相关的功能属性有关。相对于针叶树种而言,阔叶树叶片薄而宽大,光合能力(在适宜条件下测得的光合速率)较强,水分蒸散也快,其净效应导致阔叶树种水分利用效率较高[22]。上述研究结果表明,未来气温升高、降水增加情景下,长白山温带森林区典型植物水分利用效率会下降,固定相同数量的碳所消耗的水分将增加。

表4 4种森林土壤δ13C丰度与log SOC值之间回归模型参数
3.2 土壤碳含量、δ13C丰度的分布格局及生态学意义
4个森林土壤SOC含量随深度呈指数下降,20 cm以下土层SOC差异不大,主要归因于叶片和根系凋落物归还量的差异。温带森林凋落物层较厚,微生物比较活跃,表层SOC累积明显;而且大约一半的植物地下生物量集中在土壤表层(0—10 cm)[27],20 cm以下土层有机质归还量少。与原假设相反,本研究并没有观测到SOC随着海拔高度的增加而增加。凋落物层碳含量随着海拔高度的增加下降,而0—20 cm矿质层土壤是阔叶红松林最高、其次是岳桦林,两个暗针叶林最低(图2,表3)。凋落物和矿质层碳含量的分布格局分别归因于植物功能型和土壤质地的差异。阔叶红松林和岳桦林土壤粘粉粒含量(50.1%—80.6%)显著高于两个暗针叶林土壤(24.07%—32.13%)(表1),一般质地较细的土壤更易于SOC的保持与稳定[28]。另外,阔叶树种较针叶树种光合作用强、叶片C/N较低,更易于分解和转化[22]。上述研究结果表明,与植物类型、土壤质地等因子相比,气温、降水等气候要素对长白山垂直带森林表层SOC含量的影响可能居次要地位。相似地,Zhang等[29]研究表明,对于主要来源于凋落物输入的SOC累积而言,植被比气候更为重要;植物凋落物输入和分解决定了森林SOC的储量[30]。但是,在区域或国家尺度上,气候、植被和土壤属性共同控制着SOC 储量的分布格局。Yang等[31]发现气候和植被是控制中国SOC储量分布的关键因子,分别能够解释SOC储量变异的36.2%和42.7%。总体而言,植物叶片与凋落物、土壤碳含量随海拔梯度的变化趋势不同,叶片碳含量主要受植物功能型、气候因子制约,而凋落物、土壤碳含量主要受土壤质地和植物功能型驱动,主控因子不尽相同。
4个森林SOM的δ13C值随土壤深度增加呈现富集的趋势,这与凋落物分解、土壤腐殖化过程中碳同位素分馏以及“Suess”效应有关[21,32]。相似的现象常见于其他的森林生态系统,从地表植被到土壤剖面底层δ13C富集度变化范围为1‰—3‰[33]。地表SOC基本上是较年轻、不稳定的化合物,深层SOM分解时间较长,多为年代较老、稳定性较高的有机化合物[6,34]。土壤δ13C的这种分布格局常用于评估土壤碳的周转以及迁移规律[6]。基于logSOC和δ13C的回归模型,我们发现岳桦林回归方程斜率K值最大,其次是两个暗针叶林,而阔叶红松林最低,表明岳桦林SOC周转最快,阔叶红松林SOC周转最慢,这与原假设不一致。可见,SOC周转并不单单受温度驱动,可能与地形坡度以及土壤溶解性有机碳(DOC)淋失有关。岳桦林坡度最大,降水最高,加之岳桦叶片(阔叶型)易于降解,产生的DOC随地表径流流失明显,土壤碳周转更快[17]。相反,阔叶红松林位于长白山垂直带谱的基带,地形平缓,土壤粘重,DOC垂直淋溶和水平迁移量均不大。利用logSOC和δ13C的SOM周转模型,Fang等[10]也评价外源性氮素输入对不同演替阶段亚热带森林SOC周转的影响,发现施氮抑制贫氮森林SOC周转,增加土壤碳储量,但是加剧富氮森林土壤DOC淋溶,不利于土壤碳截存。值得一提的是,logSOC和δ13C的SOM周转模型只能定性地概括不同生态系统类型或实验处理下SOM周转的相对快慢,无法准确量化SOM周转的速率或量级[35],未来的研究中需要结合室内培养实验和14C示踪技术来弥补这一研究不足。
4 结论
利用稳定性碳同位素技术研究了长白山垂直带4种森林植物叶片-凋落物-土壤连续体碳含量和δ13C丰度的分布格局,阐明SOM周转沿着海拔梯度的变化规律。研究结果表明,针叶树种叶片碳含量显著高于阔叶树种,而δ13C值表现相反,表明针叶树种潜在的碳蓄积能量更强,固碳耗水成本更高;凋落物碳含量随海拔增加而下降,矿质表层土壤SOC含量阔叶红松林和岳桦林显著高于两个暗针叶林,体现植被类型和土壤质地对SOC含量持留的支配作用。总的来说,高海拔岳桦林SOC周转最快,其次是暗针叶林,阔叶红松林最低,是气候、植被类型、土壤质地等多个因素综合作用的结果。研究结果暗示在探讨环境因子对陆地生态系统碳循环和碳平衡的影响时需考虑研究尺度,不同的尺度(局地、样带、区域、全球)驱动因子并不完全相同。然而,基于logSOC和δ13C的SOM周转模型只能定性地概括不同生态系统类型下SOM周转的快慢,并不能量化SOM周转速率的大小,未来的研究中需要结合室内培养实验和14C示踪技术来深入理解SOM周转及其驱动机制。
[1] 金峰, 杨浩, 赵其国. 土壤有机碳储量及影响因素研究进展. 土壤, 2000, 32(1): 11- 17.
[2] Schlesinger W H. Evidence from chronosequence studies for a low carbon-storage potential of soils. Nature, 1990, 348(6298): 232- 234.
[3] Lal R. Forest soils and carbon sequestration. Forest Ecology and Management, 2005, 220(1/3): 242- 258.
[4] Smolander A, Kitunen V. Soil microbial activities and characteristics of dissolved organic C and N in relation to tree species. Soil Biology and Biochemistry, 2002, 34(5): 651- 660.
[5] Deng X W, Han S J, Hu Y L, Zhou Y M. Carbon and nitrogen transformations in surface soils under Ermans birch and dark coniferous forests. Pedosphere, 2009, 19(2): 230- 237.
[6] Bernoux M, Cerri C C, Neill C, de Moraes J F L. The use of stable carbon isotopes for estimating soil organic matter turnover rates. Geoderma, 1998, 82(1/3): 43- 58.
[7] Jobbágy E G, Jackson R B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications, 2000, 10(2): 423- 436.
[8] Balesdent J, Girardin C, Mariotti A. Site-relatedδ13C of tree leaves and soil organic-matter in a temperate forest. Ecology, 1993, 74(6): 1713- 1721.
[9] Michener R, Lajtha K. Stable Isotopes in Ecology and Environmental Science. 2nd ed. Malden, MA: John Wiley & Sons, 2007: 1- 594.
[10] Fang H J, Yu G R, Cheng S L, Mo J M, Yan J H, Li S G.13C abundance, water-soluble and microbial biomass carbon as potential indicators of soil organic carbon dynamics in subtropical forests at different successional stages and subject to different nitrogen loads. Plant and Soil, 2009, 320(1/2): 243- 254.
[11] Fang H J, Cheng S L, Yu G R, Yang X M, Xu M J, Wang Y S, Li L S, Dang X S, Wang L, Li Y N. Nitrogen deposition impacts on the amount and stability of soil organic matter in an alpine meadow ecosystem depend on the form and rate of applied nitrogen. European Journal of Soil Science, 2014, 65(4): 510- 519.
[12] Del Galdo I, Six J, Peressotti A, Cotrufo M F. Assessing the impact of land-use change on soil C sequestration in agricultural soils by means of organic matter fractionation and stable C isotopes. Global Change Biology, 2003, 9(8): 1204- 1213.
[13] Gaudinski J B, Trumbore S E, Davidson E A, Zheng S H. Soil carbon cycling in a temperate forest: radiocarbon-based estimates of residence times, sequestration rates and partitioning of fluxes. Biogeochemistry, 2000, 51(1): 33- 69.
[14] Trumbore S, Chadwick O A, Amundson R. Rapid exchange between soil carbon and atmospheric carbon dioxide driven by temperature change. Science, 1996, 272(5260): 393- 396.
[15] Richter D D, Markewitz D, Trumbore S E, Wells C G. Rapid accumulation and turnover of soil carbon in a re-establishing forest. Nature, 1999, 400(6739): 56- 58.
[16] 于贵瑞. 人类活动与生态系统变化的前沿科学问题. 北京: 高等教育出版社, 2009: 1- 543.
[17] Cheng S L, Fang H J, Yu G R, Zhu T H, Zheng J J. Foliar and soil15N natural abundances provide field evidence on nitrogen dynamics in temperate and boreal forest ecosystems. Plant and Soil, 2010, 337(1/2): 285- 297.
[18] Wang Z, Xu Z B, Li X, Peng D, Tan Z. The main forest types and their features of community structure in northern slope of Changbai Mountain. Forest Ecosystem Research, 1980, (1): 25- 42.
[19] Amundson R, Baisden W T. Stable isotope tracers and mathematical models in soil organic matter studies // Sala O E, Jackson R B, Mooney H A, Howarth R W, eds. Methods in Ecosystem Science. New York: Springer, 2000: 117- 137.
[20] Garten C T Jr, Cooper L W, Post W M III, Hanson P J. Climate controls on forest soil C isotope ratios in the southern Appalachian Mountains. Ecology, 2000, 81(4): 1108- 1119.
[21] Powers J S, Schlesinger W H. Geographic and vertical patterns of stable carbon isotopes in tropical rain forest soils of Costa Rica. Geoderma, 2002, 109(1/2): 141- 160.
[22] Chapin F S III, Matson P A, Vitousek P. Principles of Terrestrial Ecosystem Ecology. New York: Springer, 2011: 1- 546.
[23] He J S, Fang J Y, Wang Z H, Guo D L, Flynn D F B, Geng Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia, 2006, 149(1): 115- 122.
[24] Bai Y F, Wu J G, Clark C M, Pan Q M, Zhang L X, Chen S P, Wang Q B, Han X G. Grazing alters ecosystem functioning and C: N: P stoichiometry of grasslands along a regional precipitation gradient. Journal of Applied Ecology, 2012, 49(6): 1204- 1215.
[25] Ripullone F, Lauteri M, Grassi G, Amato M, Borghetti M. Variation in nitrogen supply changes water-use efficiency ofPseudotsugamenziesiiandPopulusxeuroamericana; a comparison of three approaches to determine water-use efficiency. Tree Physiology, 2004, 24(6): 671- 679.
[26] Yu G R, Song X, Wang Q F, Liu Y F, Guan D X, Yan J H, Sun X M, Zhang L M, Wen X F. Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. New Phytologist, 2008, 177(4): 927- 937.
[27] Arai H, Tokuchi N. Factors contributing to greater soil organic carbon accumulation after afforestation in a Japanese coniferous plantation as determined by stable and radioactive isotopes. Geoderma, 2010, 157(3/4): 243- 251.
[28] 黄昌勇, 徐建明. 土壤学(第三版). 北京: 中国农业出版社, 2010: 1- 379.
[29] Zhang M, Zhang X K, Liang W J, Jiang Y, Dai G H, Wang X G, Han S J. Distribution of soil organic carbon fractions along the altitudinal gradient in Changbai Mountain, China. Pedosphere, 2011, 21(5): 615- 620.
[30] Blair G J, Lefroy R D B, Lisle L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Australian Journal of Agricultural Research, 1995, 46(7): 1459- 1466.
[31] Yang Y H, Mohammat A, Feng J M, Zhou R, Fang J Y. Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry, 2007, 84(2): 131- 141.
[32] Ehleringer J R, Buchmann N, Flanagan L B. Carbon isotope ratios in belowground carbon cycle processes. Ecological Applications, 2000, 10(2): 412- 422.
[33] Liu W G, Feng X H, Ning Y F, Zhang Q L, Cao Y N, An Z S.δ13C variation of C3and C4plants across an Asian monsoon rainfall gradient in arid northwestern China. Global Change Biology, 2005, 11(7): 1094- 1100.
[34] 于贵瑞, 王绍强, 陈泮勤, 李庆康. 碳同位素技术在土壤碳循环研究中的应用. 地球科学进展, 2005, 20(5): 568- 577.
[35] Gao J Q, Lei G C, Zhang X W, Wang G X. Canδ13C abundance, water-soluble carbon, and light fraction carbon be potential indicators of soil organic carbon dynamics in Zoigê wetland? Catena, 2014, 119: 21- 27.
Organiccarbondynamicsoftheleaf-litter-soilcontinuuminthetypicalforestsoftheChangbaiMountaintransect:ananalysisofstablecarbonisotopetechnology
SI Gaoyue1, LI Xiaoyu2, CHENG Shulan1,*, FANG Huajun1,2, YU Guirui2, GENG Jing2, HE Shun1, YU Guangxia1
1UniversityofChineseAcademyofSciences,Beijing100049,China2KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicalSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China
国家自然科学基金资助项目(41471212, 31470558, 31290222, 31290221);国家重点研发计划资助项目(2016YFC0500603,2016YFC0503603);青年创新研究团队资助项目(LENOM2016Q0004)
2016- 05- 19; < class="emphasis_bold">网络出版日期
日期:2017- 03- 27
*通讯作者Corresponding author.E-mail: slcheng@ucas.ac.cn
10.5846/stxb201605190967
司高月,李晓玉,程淑兰,方华军,于贵瑞,耿静,何舜,于光夏.长白山垂直带森林叶片-凋落物-土壤连续体有机碳动态——基于稳定性碳同位素分析.生态学报,2017,37(16):5285- 5293.
Si G Y, Li X Y, Cheng S L, Fang H J, Yu G R, Geng J, He S, Yu G X.Organic carbon dynamics of the leaf-litter-soil continuum in the typical forests of the Changbai Mountain transect: an analysis of stable carbon isotope technology.Acta Ecologica Sinica,2017,37(16):5285- 5293.
猜你喜欢
雷锋·强军号(2023年4期)2023-04-29 00:44:03
建筑与预算(2022年6期)2022-07-08 13:59:18
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
中国林副特产(2020年5期)2020-10-09 08:24:14
当代音乐(2020年1期)2020-05-07 01:50:42
现代园艺(2017年19期)2018-01-19 02:50:06
防护林科技(2017年7期)2017-09-01 01:08:43
建筑与预算(2014年7期)2014-10-14 13:28:52
满族文学(2014年5期)2014-09-22 23:50:36
建筑与预算(2014年5期)2014-04-11 07:29:28
