
硝酸盐诱导蛋白与谷子抗旱性的关系研究
2017-10-10 02:03:19陈利青王雄郭展魏东禾璐候蕊韩渊怀
山西农业大学学报(自然科学版) 2017年10期
陈利青,王雄,郭展,魏东,禾璐,候蕊,韩渊怀,5*
(1.山西农业大学 生命科学学院,山西 太谷 030801;2.山西农业大学 农学院,山西 太谷 030801;3.山西省农业科学院 玉米研究所,山西 忻州 034000;4.山西省晋城市城区农业委员会,山西 晋城 048400;5.山西农业大学 生物工程研究所,山西 太谷 030801)
硝酸盐诱导蛋白与谷子抗旱性的关系研究
陈利青1,王雄2,郭展2,魏东2,禾璐3,候蕊4,韩渊怀2,5*
(1.山西农业大学 生命科学学院,山西 太谷 030801;2.山西农业大学 农学院,山西 太谷 030801;3.山西省农业科学院 玉米研究所,山西 忻州 034000;4.山西省晋城市城区农业委员会,山西 晋城 048400;5.山西农业大学 生物工程研究所,山西 太谷 030801)
[目的]硝酸盐诱导蛋白(NOI)是一类重要的抗逆蛋白,在多种植物中发挥作用。谷子作为重要的旱作植物,在农作物生产中占有重要地位,探究谷子中NOI与抗旱性的关系,对发掘谷子抗旱相关基因具有重要意义。[方法]运用生物信息学方法,对谷子中编码NOI的基因进行分析,并在PEG干旱胁迫下,探究抗旱品种勾勾母鸡咀(GG)和干旱敏感品种晋汾16(JF16)中编码硝酸盐诱导蛋白(NOI)的基因的相对表达情况。[结果]编码NOI的基因的启动子中存在与干旱相关的响应元件。在PEG干旱胁迫处理与非胁迫处理条件下,勾勾母鸡咀(GG)和晋汾16(JF16)中编码NOI的基因的表达模式差异显著。[结论]基于上述分析,推测硝酸盐诱导蛋白(NOI)可能参与谷子的抗旱调节,为深入研究谷子抗旱机理提供了理论基础和依据。
谷子;NOI;抗旱性
Abstract:[Objective]Nitrate-induced protein(NOI)is an important protein related to stress and works in variety of plants. As an important drought-tolerance crop, foxtail millet plays a significant role in crop production, so it is important sense to explore the connection between NOI and drought tolerance and the unique genetic resources.[Methods]Using bioinformatics,we analysed the genes coding NOI and studied the relative expression level in the drought-tolerant variety Go Go Mu Ji Ju (GG) and the drought sensitive variety Jin Fen 16 (JF16).[Results]The results showed that there were response elements associated with drought response in the gene’s protein promoter. Under PEG stress, Go Go Mu Ji Ju (GG) and Jin Fen 16 (JF16),showed a strikingly different expression pattern.[Conclusion]Based on the analysis above, we speculate that NOI could be involved in the regulation of drought tolerance in foxtail millet and have a different expression in varieties of foxtail millet. The study could provide a theoretical basis and foundation for further study of drought tolerance in foxtail millet.
Keywords:Foxtail millet, NOI, Drought tolerance
谷子(Setariaitalica),禾本科狗尾草属,一年生草本植物,原产于中国,是喜温、短日照耐旱作物,常种植于我国北方地区[1]。随着全球气候逐渐变暖,干旱问题日趋凸显,对粮食生产造成严重威胁。谷子作为一种耐旱作物,在可持续农业发展中地位日益突出。
硝酸盐诱导蛋白(Nitrate-induced protein,NOI)广泛存在于植物中,现已证明NOI参与多种植物抗逆性免疫机制。例如,RIN4的C末端和N末端的NOI域,包含两个保守序列(PXFGXW and Y/FTXXF),位于C末端PXFGXW含有AvrRpt2可识别切割位点(RCS)[2],而AvrRpt2裂解RCS2与RPS2活化诱导及囊泡运输相关[3,4],AvrRpt2还诱导激活ETI,从而增强PTI(PAMP-triggered immunity)反应[5]。当RPM1介导ETI时,需要C末端NOI域参与[6]。除此之外,NOI还参与气孔调节,通过控制细胞膜上H+-ATPases 活性控制气孔开合大小[7]。当H+-ATP酶活性降低时,气孔关闭,PTI应答增强,从而提高植物防御。
目前,在水稻,拟南芥[8]和大豆[9,10]等植物中,均发现存在NOI,并在抗逆性方面发挥重要作用,而关于NOI在谷子抗逆中的作用还缺乏研究。本文通过在PEG模拟干旱胁迫下,NOI在勾勾母鸡咀(GG,干旱品种)和晋汾16(JF16,干旱敏感品种)2个谷子品种的表达情况,探究该蛋白在谷子抗旱中的作用,为深入探索谷子抗旱机制奠定一定理论依据。
1 材料与方法
1.1 研究材料
本文研究材料勾勾母鸡咀(GG,干旱品种)及晋汾16(JF16,干旱敏感品种)均由山西农业大学农业生物工程研究所提供。
1.2 研究方法
1.2.1 生物信息学分析方法
利用Phytozomev12.0获取谷子参考基因组豫谷1号中编码NOI的基因序列信息。
利用plantCARE对编码NOI的基因起始密码子上游1 500 bp核苷酸序列进行启动子元件分析。
利用DNAMAN 6.0预测蛋白质分子量和等电点等理化性质。
利用软件MEGA 7.0构建进化树。
1.2.2 试验材料
在人工气候室中,将勾勾母鸡咀和晋汾16放置在营养土∶蛭石=3∶1混合的基质中培植,出苗前,进行光培养(14 h光照/28 ℃)和暗培养(10 h黑暗/23 ℃)。出苗后每周浇3次蒸馏水,3周后,釆用20% PEG-6000模拟干旱胁迫,处理材料0.5 h,对照用蒸馏水处理,每个处理取3株用于RNA的提取,进行表达谱测序。
2 结果与分析
2.1 谷子中编码NOI的基因的序列分析
利用水稻中NOI的氨基酸序列在豫谷1号数据库中进行blast比对,找到谷子中同源性较高的编码NOI的基因序列。如表1所示,Seita.8G049900.1,Seita.8G049900.2和Seita.8G049900.3来自于同一基因,由于mRNA剪切位点不同,出现3种转录本和核心编码序列,其差异出现在基因序列的起始端,而Seita.9G012000.1 和Seita.9G012000.2序列差异出现在末端。编码硝酸盐诱导蛋白(NOI)的基因的分布存在差异,位于第6,第8,第9号染色体上,且位于8号染色体的居多。5个基因中,基因组长度不一,介于2 739~15 523 bp间,最长的为Seita.8G039500.1,长度为15 523 bp,最短的为Seita.6G219800.1和Seita.6G219800.2,长度为2 739 bp。核心编码区长度在702~6 633 bp之间。其中,核心编码区长度最长的是Seita.8G039500.1,为6 633 bp,核心编码区最短长度为702 bp,是Seita.6G219800.2。从表1中可看出,除Seita.8G049900.3外,其他基因组长度与编码蛋白长度呈正相关性。除此之外,外显子数目也存在差异,在7~10个之间。
2.2 谷子中编码NOI的基因理化性质分析
从表2可以看出,5个基因的氨基酸序列长度存在明显的差异,在233~2 210 aa之间,其中Seita.8G039500.1的氨基酸序列最长,为2 210 aa,Seita.6G219800.2氨基酸序列最短,为233 aa。NOI分子量介于0.245 981~2.500 000 kDa间,其中分子量最大的是Seita.8G039500.1,为2.488 738 kDa,分子量最小的是Seita.6G219800.2,为0.245 981 kDa。编码蛋白质的氨基酸序列长度与分子量呈正相关。理论等电点(PI)跨度较大,介于6.50~9.74之间,最大的是Seita.6G219800.1和Seita.6G219800.2,为9.74,最小的为Seita.8G089100.1,为6.50。

表1 谷子中编码NOI的基因基本信息Table 1 Basic information of the genes coding NOI in foxtail millet
表2谷子中编码NOI的基因基本结构分析
Table2 Primary structure analysis of the genes coding NOI in foxtail millet
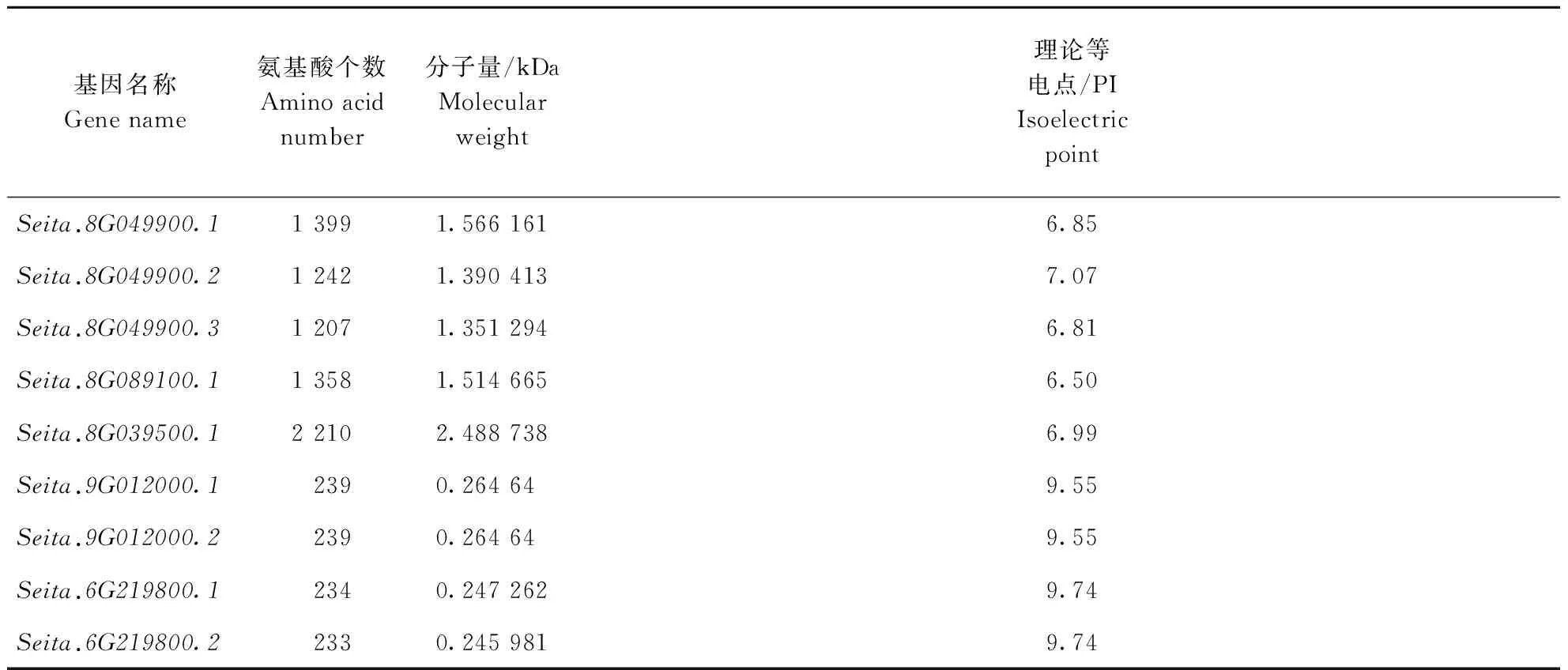
基因名称Genename氨基酸个数Aminoacidnumber分子量/kDaMolecularweight理论等电点/PIIsoelectricpointSeita.8G049900.113991.5661616.85Seita.8G049900.212421.3904137.07Seita.8G049900.312071.3512946.81Seita.8G089100.113581.5146656.50Seita.8G039500.122102.4887386.99Seita.9G012000.12390.264649.55Seita.9G012000.22390.264649.55Seita.6G219800.12340.2472629.74Seita.6G219800.22330.2459819.74
2.3 不同物种中NOI进化树分析
图1是不同物种间的NOI的蛋白序列运用MEGA7软件制作的进化树。从图1可看出,进化树主要有2个分支,然后又逐渐分成很多小枝:第一分支主要包括谷子(Seita.6G219800.1,Seita.6G219800.2,Seita.9G012000.1,Seita.9G012000.2),矮牵牛花(AAQ10002.1),玉米(NP_001148391.2),蓖麻(EEF30175.1),蚕豆基因(BAJ78239.1),拟南芥(OAO91965.1);第二分支由谷子(Seita.8G049900.1,Seita.8G049900.2,Seita.8G049900.3,Seita.8G039500.1,Seita.8G039500.2)和水稻(Os09g0253000)。图中也可以看出Seita.8G049900基因,Seita.9G012000基因和Seita.6G219800基因由于可变性剪切产生多种不同的基因片段。

图1 不同物种NOI进化树分析Fig.1 Phylogenetic tree analysis of NOI in different species
2.4 谷子中编码NOI的基因启动子元件分析
综合表3、表4可以看出,这些基因中,除大量基本应答元件外,还有许多不同类型的抗逆应答元件,如干旱相关的MYB结合位点MBS,胁迫响应相关的元件TC-rich repeats,以及各种激素响应元件ABRE(脱落酸响应元件、TGACG-motif(茉莉酸甲酯响应元件),TCA-element(水杨酸响应元件)等。其中光应答元件最多(10~24个),其次是逆境胁迫相关元件。由于基因不同,其含有的与抗旱相关元件种类与数目存在差异。5个基因中,Seita.8G049900中含有的总元件数目最多,为119个。Seita.8G039500.1最少,为79个。Seita.8G049900不含与MeJA,ETH和GA相关的应答元件;Seita.8G089100.1,Seita.8G039500.1中都不含有IAA应答元件。在这些基因中,Seita.6G219800含有与胁迫相关的调控元件的数目最多,Seita.9G012000最少。
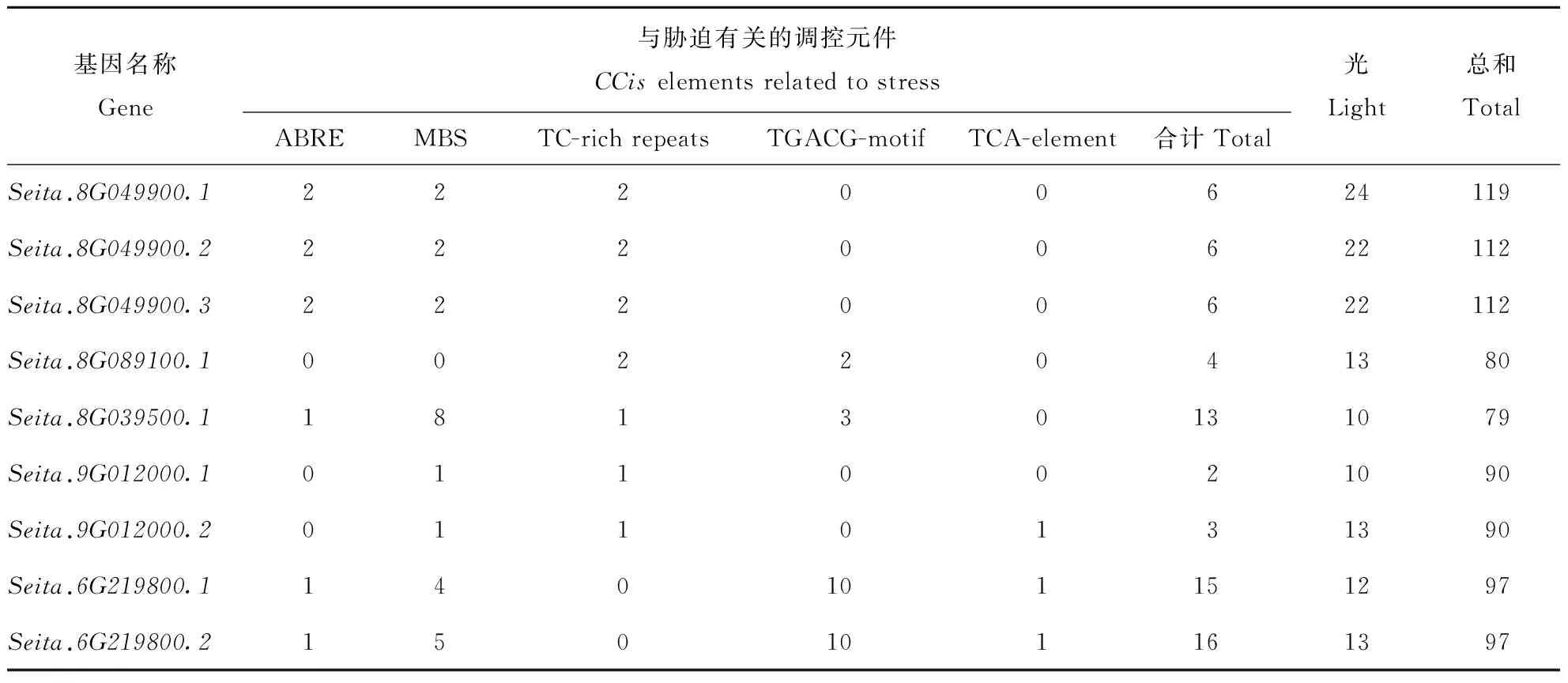
表3 谷子中编码NOI的基因启动子元件分析Table 3 The promoter element analysis of the genes coding NOI in foxtail millet
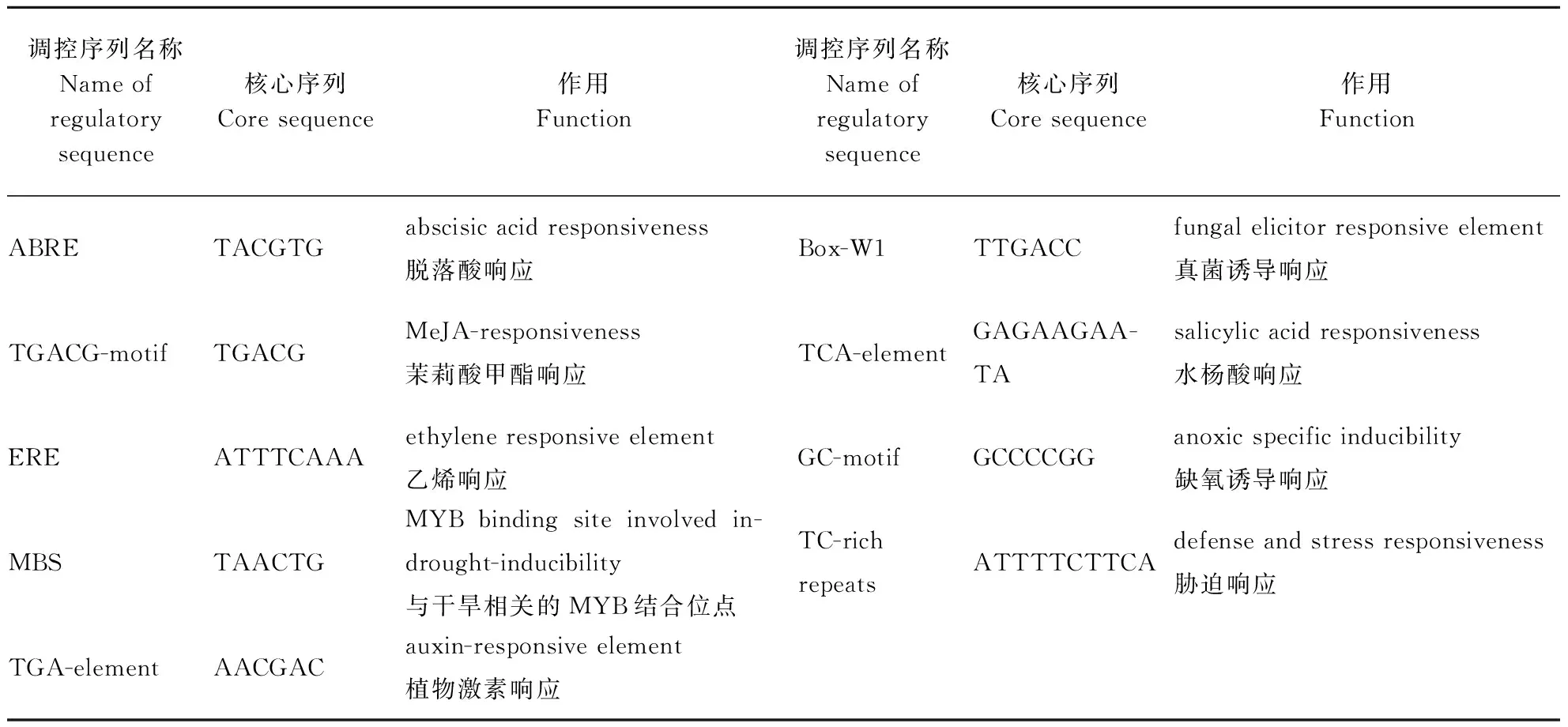
表4 启动子顺式元件Table 4 Promoter cis elements
2.5谷子Seita.8G049900.1基因在GG和JF中干旱胁迫后的表达模式分析
从图2中可看出,勾勾母鸡咀在干旱情况下Seita.8G049900.1基因表达水平高于正常情况下的表达水平29倍;而在晋汾16中,干旱情况下Seita.8G049900.1基因的表达水平较正常情况下低29倍。正常情况下,Seita.8G049900.1在晋汾16中的表达水平是勾勾母鸡咀中表达水平的30倍;干旱情况下,勾勾母鸡咀的基因相对表达水平显著提高,而晋汾16中Seita.8G049900.1基因相对表达水平出现明显的下调,且此时Seita.8G049900.1在勾勾母鸡咀中的表达水平是晋汾16中表达水平的30倍。
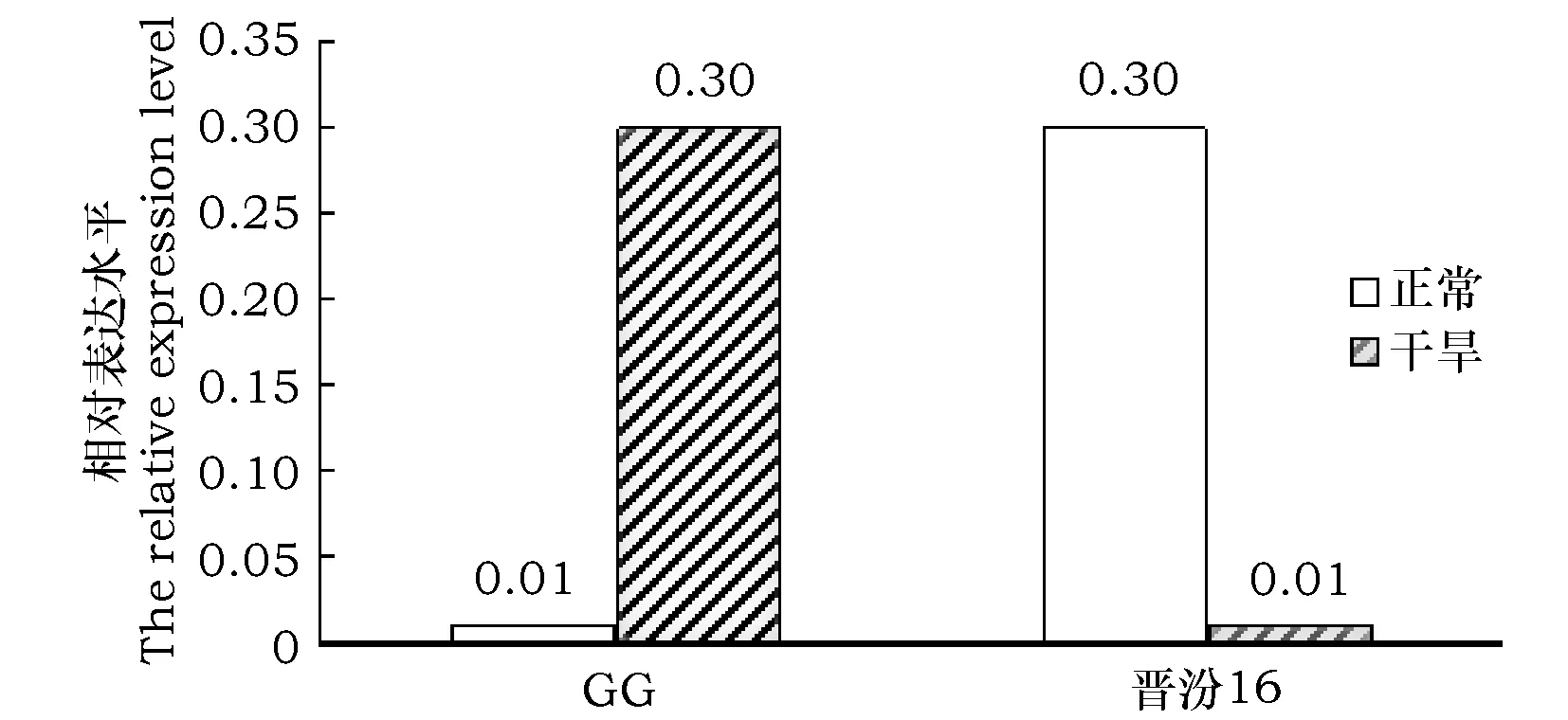
图2 GG和JF16中Seita.8G049900.1的表达水平分析Fig.2 Expression level analysis of Seita.8G049900.1 gene in GG and JF16
3 讨论与结论
NOI是植物中重要的抗逆蛋白。通过分析谷子中编码NOI的基因序列,发现其启动子序列中含有与抗逆相关的元件TC-rich repeats、干旱胁迫响应相关的MYB结合位点MBS、茉莉酸甲酯响应元件、水杨酸响应元件等。谷子具有较强的耐旱性,结合实际情况,推测谷子中存在抗旱调节机制。
通过试验在基因水平上分析谷子的抗旱机制。干旱情况下,勾勾母鸡咀中编码NOI的基因表达明显上调,且超过正常浇水时表达的30倍;而在晋汾16中,正常浇水情况下,Seita.8G049900.1基因表达量较高,干旱情况下,Seita.8G049900.1基因表达明显下调,说明不同环境下,不同谷子品种中编码NOI基因的表达模式不同。Afzal AJ[11]等人在研究NOI的免疫时发现不同试验条件下,每个NOI域具有不同的功能,且功能间表达情况不同[7],这与本试验结果相一致,即在勾勾母鸡咀和晋汾16的抗旱性的研究中发现,Seita.8G049900.1基因在2个品种中的表达量存在显著差异,类比NOI的免疫调节机制,推测编码NOI的基因在抗旱调节机制中表达具有差异,可能是由于末端NOI域作用不同引起的,具体调节机制需进一步探究。同时,在试验中勾勾母鸡咀和晋汾16在PEG胁迫和浇水情况下Seita.8G049900.1基因的表达趋势相反,这与Kim MG, da Cunha L等人[7]的研究结果一致,表明其在作物中的表达调节机制多样化,为进一步研究提供了理论基础与试验依据。
综上所述,通过试验预测谷子中编码NOI的基因可能参与谷子对逆境的应答,响应胁迫,且不同NOI域在调节抗旱中功能各异,为深入研究谷子抗旱机理提供了理论基础和依据。
[1]张雁明,刘晓东,马建萍,等.谷子抗旱研究进展[J].山西农业科学,2013,41(3):282-285.
[2]Afzal A J,Kim J H,Mackey D.The role of NOI-domain containing proteins in plant immune signaling[J].BMC Genomics,2013,14(1):327.
[3]Kim HS,Desveaux D,Singer AU,et al.The pseudomonas syringae effector AvrRpt2 cleaves its C-terminally acylated target,RIN4, from Arabidopsis membranes to block RPM1 activatio[J].Proceedings of the National Academy of Sciences of the United States of America,2005,102(18):6496-6501.
[4]Day B,Dahlbeck D,Huang J,et al.Molecular basis for the RIN4 negative regulation of RPS2 disease resistance[J].Plant Cell,2005,17(4):1292-1305.
[5]Chung EH,Da Cunha L,Wu AJ,et al.Specific threonine phosphorylation of a host target by two unrelated type III effectors activates a host innate immune receptor in plants[J].Cell Host and Microbe,2011,9(2):125-136.
[6]Nomura K,Mecey C,Lee Y N,et al.Effector-triggered immunity blocks pathogen degradation of an immunity-associated vesicle traffic regulator in Arabidopsis[J].Proceedings of the National Academy of Sciences of the United States of America,2011,108(26):10774-10779.
[7]Kim M G,Da Cunha L,McFall A J,et al.Two Pseudomonas syringae type III effectors inhibit RIN4-regulated basal defense in Arabidopsis[J].Cell,2005,121(5):749-759.
[8]Boyes D C,Nam J,Dangl J L.The protein that is degraded coincident with the hypersensitive response[J]. Proceedings of the National Academy of Sciences of the United States of America,1998,95(26):15849-15854.
[9]Selote D,Kachroo A.RPG1-B-der Arabidopsis thaliana RPM1 disease resistance gene product is a peripheral plasma membrane ived resistance to AvrB-expressing pseudomonas syringae requires RIN4-like proteins in soybean[J].Plant Physiology,2010,153(3):1199-1211.
[10]Melotto M,Underwood W,Koczan J,et al.Plant stomata function in innate immunity against bacterial invasion[J]. Cell,2006,126(5):969-980.
[11]Afzal A J,Da Cunha L,Mackey D.Separable fragments and membrane tethering of Arabidopsis RIN4 regulate its suppression of PAMP-triggered immunity[J].Plant Cell,2011,23(10):3798-3811.
(编辑:韩志强)
Thepreliminarystudyabouttherelationshipbetweennitrate-inducedproteinanddroughttoleranceinfoxtailmillet
Chen Liqing1, Wang Xiong2, Guo Zhan2, Wei Dong2, He Lu3, Hou Rui4, Han Yuanhuai2,5*
(1.CollegeofLifeScience,ShanxiAgriculturalUniversity,Taigu030801,China;2.CollegeofAgriculture,ShanxiAgriculturalUniversity,Taigu030801,China;3.MaizeResearchInstitutetoShanxiAcademyofAgriculturalSciences,Xinzhou034000,China;4.UrbanAgricultureCommitteeofShanxiJincheng,Jincheng048400,China; 5.InstituteofAgriculturalBioengineering,ShanxiAgriculturalUniversity,Taigu030801,China)
S515
A
1671-8151(2017)10-0685-05
2017-05-02
2017-05-31
陈利青(1990-),女(汉),山西晋城人,硕士,研究方向:生物化学与分子生物学
*通信作者:韩渊怀,教授,博士生导师,Tel:0354-6287239;E-mail:swgctd@163.com
国家自然科学基金(31371693)
猜你喜欢
小学生学习指导(中年级)(2022年5期)2022-06-01 03:16:58
当代陕西(2021年21期)2022-01-19 02:00:32
数学小灵通(1-2年级)(2021年10期)2021-11-05 07:20:30
艺术大观(2019年33期)2019-10-12 15:37:21
创新作文(小学版)(2019年7期)2019-10-12 08:37:10
文学少年(小学版)(2017年8期)2017-09-04 14:11:14
幼儿园(2016年6期)2016-04-26 10:57:10
现代农业(2015年5期)2015-02-28 18:40:53
都市家教·上半月(2014年4期)2014-05-30 19:24:19
祝你幸福·午后版(2013年9期)2013-09-22 09:56:54
