
从岩石学及微区同位素探讨四川盆地灯影组皮壳-葡萄状白云石成因
2017-09-15 07:21:24钱一雄冯菊芳何治亮张克银董少峰尤东华张永东
石油与天然气地质 2017年4期
钱一雄,冯菊芳,何治亮,张克银,金 婷,董少峰,尤东华,张永东
(1.中国石化 石油勘探开发研究院 无锡石油地质研究所,江苏 无锡 214126; 2.中国石化 石油勘探开发研究院,北京 100083; 3.中国石化 西南油气分公司 石油勘探开发研究院,四川 成都 610081; 4.中国石化 江苏油田分公司 采油二厂,江苏 金湖 211600)
从岩石学及微区同位素探讨四川盆地灯影组皮壳-葡萄状白云石成因
钱一雄1,冯菊芳2,何治亮2,张克银3,金 婷4,董少峰1,尤东华1,张永东1
(1.中国石化 石油勘探开发研究院 无锡石油地质研究所,江苏 无锡 214126; 2.中国石化 石油勘探开发研究院,北京 100083; 3.中国石化 西南油气分公司 石油勘探开发研究院,四川 成都 610081; 4.中国石化 江苏油田分公司 采油二厂,江苏 金湖 211600)
四川盆地灯影组中的“皮壳、栉壳及葡萄状”白云石是一种非常特殊结构白云石。对四川盆地灯影组葡萄-皮壳状及基岩岩石学、碳氧与锶同位素等研究表明:皮壳-葡萄状白云石早期由内纤状(AC)、外为束状-放射状海底纤柱状(RFC)的文石及犬牙状高镁方解石或粒状方解石构成。不同尺度 “球状-哑铃”或菱形的泥微晶白云石沿C轴呈台阶式相向生长,反映了海水潜流带为主的生长模式。微区同位素揭示了环带中的粉细晶白云石要比黑、白相间纤状白云石中的δ13C,δ18O负偏明显;黑色纤状又较白色纤状白云石δ13C,δ18O正偏。环带至少有3种变化情形:① δ18O(PDB),δ13C(PDB)负偏,87Sr/86Sr较高,较宽的明、暗阴极条带,指示了周期性弱氧化与弱还原环境的变化海水并叠加了大气淡水作用;②有序度变化大,从环带核部至边缘,δ13C(PDB)缓慢下降,δ18O(PDB)缓慢升高,87Sr/86Sr接近同时代的海水平均值,核部中等橙红向外为不发光-暗淡发光,反映了核部弱还原向外变为弱氧化、海水并有持续大气淡水的补给;③有序度较低、从环带核部至边缘,δ13C(PDB)缓慢上升,δ18O(PDB)先升后降,边缘稍为偏负,中等橙红色、间隔有相对暗淡阴极发光,推测为停滞浓缩海水为主、后有弱氧化大气淡水补给。据此推断,皮壳-葡萄状白云石是在新元古南华纪冰期的特殊海水背景下,经历了过饱和海水中微生物中细菌参于下成核、准稳态含镁矿物沉淀(AC-RFC)、大气淡水下水岩作用粒状方解石(RFC-PC)和浓缩海水中的准同生云化作用过程。
岩石学;微区同位素;白云石;灯影组;四川盆地
皮壳-葡萄状白云石是固定生长质点且发育等厚皮壳层的半球体[1]。直径大者为8~10 cm,小者不到1 cm,沿层面或缝洞呈“钙华”状分布。发育玛瑙纹带或条带状(皮壳),由波状暗色层和浅色层交替组成,且从内到外纹层颜色变浅,纹层厚1~2 mm。
灯影组皮壳-葡萄状白云石成因与储层发育密切相关。前人对灯影组的“皮壳或栉壳状”白云岩成因已进行了较为详细地岩相学及地球化学研究[1-14],大致可归纳成3种认识:①原生海水或生物化学成因;②表生大气水岩溶产物;③沉积或与成岩作用叠加产物,强调其既可形成于沉积期,也可发育于早期成岩、表生成岩至埋藏期成岩改造。
不同观点认识的分岐在于以下3个关键问题:一是皮壳-葡萄状白云石(自泥晶化或纤状、环带白云石)是否经历了早期蓝细菌参与的微生物调制作用;二是皮壳-葡萄状白云石化过程中(原生或准同生?)的流体性质及环境;三是如何协调岩石学与地球化学资料中解释中出现的矛盾?
本文选择了林1井、金石1井和乐山沙湾范店、金沙岩孔剖面中(图1)中灯(灯影组)二段至及灯一段及少量灯四段的基岩白云岩与皮壳-葡萄状白云石样品,在详细的岩相学研究的基础上,采用了微区碳氧同位素、扫描电镜等方法,进一步厘定了皮壳-葡萄状白云石的海水为主并有大气淡水等叠加介质中的沉积-早期成岩产物的地质地球化学依据。
1 采样分析
本次样品包括乐山沙湾范店(样品编号FD)、金沙岩孔(样品编号JSYK)、林1井(样品编号Lin1)、金石1井(样品编号JS1)的灯影组薄片288件,阴极发光35件,白云岩及缝洞白云石有序度58件,岩石及缝洞充填物(白云石和方解石)的碳、氧同位素共120件(微区29件)和锶同位素54件。其中,白云石有序度分析由成都理工大学油气藏地质及开发工程国家重点实验室完成,仪器为D/max-2500型X-射线衍射仪。阴极发光及扫描电镜是在中国石化石油勘探开发研究院构造与沉积储层实验室完成,仪器分别为BLM-3RX型阴极发光仪和Oxford扫描电镜。碳、氧同位素分析是由国家地质实验测试中心矿床所完成,采用McCrea(1950)100%正磷酸法和Finngan-MAT252气体质谱仪,分析精度±0.2‰。微区碳、氧同位素分析采用微钻取样,直径为1 mm,并标定原位阴极发光图像,样品研磨小于200 目,测试方法同上,由中国科学院地质与地球物理研究所完成。锶同位素分析是由核工业地质研究院测试中心完成。采用IsoProbeT热电离质谱仪,精度高于3×10-6。

图1 川东南震旦系灯影组露头地质剖面及部分井位Fig.1 Location of outcrop and wells of Dengying Formation in the southeast Sichuan Basin
2 岩石学
2.1 岩性特征
四川盆地灯影组由下至上可划分为4段[15],皮壳-葡萄状白云石主要出现在灯二段及灯一段。由于受桐湾运动影响,抬升暴露使得灯四段和灯三段部分受到了一定程度剥蚀。皮壳-葡萄状白云石主要呈层状、穿层状或缝洞中产出,外形呈三角状,外圈为灰黑色条带,中心为白色-乳白色的中晶白云石及残留孔洞(图2a)。分布较集中处,形成生长纹相连、半球状或席状(图2b);而围岩为以潮上-潮间带富含菌藻的藻纹层、叠层石、藻凝块或砂屑白云岩[16](图2c,d)或含菌藻微晶、泥粉晶白云岩。
2.2 结构与成分
皮壳-葡萄状白云石往往具有相向的对称生长并发育了等厚纤状环边(图2f,i)以及底面凹凸不平甚至发育海底硬底构造[1]。明暗相间的“同圈层-环带”构造较为常见。其中,浅色层是由纤柱状一粒状亮晶白云石组成,呈放射状或晶簇状垂直层纹生长;暗色层具致密泥晶和纤状白云石构成,富含有机质。中心常可见纤柱状、粒状或少量悬垂型、刀刃、鞍形的白云石或乳白色石英、黄铁矿、沥青等充填及残留的溶蚀孔洞(图2)。根据纤状、纤柱状、刀刃状岩石结构和束状消光特征以及同心圈层构造,以及“球形-哑铃”晶形(图3a)等特征推断,明暗纹层皮壳-葡萄状白云石早期应为文石和高镁方解石,细菌及藻类可能在皮壳-葡萄状白云石形成中起重要作用。
2.3 胶结世代-阴极发光
浅色纤(柱)状白云石发光相对较弱,部分纤状及粒状白云石不发光,暗色层(富藻层)白云石发光强度增加[6,11,14]。
皮壳-葡萄石状白云石的阴极发光呈现出多种类型或组合(图2e—i)。从阴极发光特征分析,可划分以下4类环带:①较宽的明、暗条带(样品JSYK-016)(图4a);②极细的纹层组(15~20 条/mm),总体为中等-暗橙红色明暗相间,以亮橙-桔红色的窄带和中等至暗桔、橙红色宽带相间为特点(样品FD-003)(图4b);③自核部至外环带,发光从亮-暗-中等桔红色变化,核部为泥(细)晶白云石发中亮橙红色,较细密集的纹层条带发暗橙红色或不发光(纤状白云石),颗粒状、横纹发育较宽的外环带发中等暗红夹不发光(样品FD-20)(图4c);④与上述②结构与变化趋势相似,即较宽的暗橙-桔红色和较窄的亮橙-桔红色环带,但具有向外壳发光强度减弱的趋势(样品JS1-2)(图4d)。

图2 四川盆地震旦系灯二、灯三段皮壳-葡萄状白云石岩相学特征Fig.2 Petrography of grape-like dolomite in the 2nd and 3rd member of Dengying Formation in the Sichuan Basina.样品JSYK-10,暗色纹层状-半球状皮壳-葡萄状泥晶白云石,构成三角形,核部发育菱形中晶白云石及少量石英;b.样品JSYK-23,边缘是明暗藻纹层泥晶云岩,中心为半球状皮壳-葡萄状泥晶白云石;c.样品FD-023,藻云岩,藻丘,大量肾形藻,边缘暗色泥晶化,含球状有机质团粒(SM);d.样品FD-020,藻纹层-藻团粒云岩,发育暗色微晶质有机球粒(SM);e.样品FD-003,中心为长快波的束状光性的纤状(AC),向外为慢波束状-放射状纤状(RFC或束状FAS?)-犬牙状泥晶白云石,并发育穿心的收缩微缝,海水潜流及大气淡水带;f.样品JSYK-002,相向生长的针状-纤状泥晶白云石(原为文石AC)及纤柱状BC(原为高镁方解石),中心为斑块粒状白云石(原为粒状方解石PC),微生物膜(MB),似角砾状-示底构造,藻纹层中泥晶套由暗色的针状-纤状泥晶白云石胶结物构成,海水潜流及大气淡水带;g.样品JSYK-023,类似于卷曲状-角砾状及空隙,斑块粒状白云石(PC),明暗的纤状纹层中的藻-细菌活动遗迹或干裂,潮上带;h.样品JSYK-034,基本同e,空隙(BC)周围为同心圈层的明暗纤状泥晶白云石,从纤状(AC)向外为海底纤状((RFC)至刀刃状-束状白云石(FAS),发育收缩微缝SCF;i.样品JSYK-033,相对厚的暗色藻纹层泥晶云岩AC,向中心明暗的纹层(MC),出现了暗色核心“豆荚状或胡桃状钙豆粒” (右下角);或片状空隙收缩微缝SCF,潮上带,海水潜流及大气淡水带

图3 四川盆地金沙岩孔灯影组藻团粒-皮壳-葡萄状云岩 (JSYK-4)扫描电镜图像Fig.3 SEM photographs of algal pellet and grape-like dolomite (JSYK-4) of the Dengying Formation in Jinshayankong section in the Sichuan Basina.藻纹层-藻球粒泥粉晶云岩中“球形-哑铃”及菱形泥微晶白云石;b.明暗色层中的不同粒级、组构白云石,顺层生呈球形、环带、水平条带;c.台阶式生长,溶液中的晶体生长模式
皮壳-葡萄石状白云石的阴极发光变化较大。暗色为主代表了海水的咸水还原环境,中等甚至较亮的桔、橙红色指示了淡水-半咸水停滞间歇弱氧化环境。总体上,皮壳状-葡萄石状白云石自核部至外圈,一般呈发光减弱,但同时也有增强再减弱,或减弱后再增强,反映了海水及咸水的还原至弱氧化的变化过程,主要受控于沉积-早期成岩流体性质变化。并不对应于一般的准同生期—浅埋藏—构造暴露大气水—埋藏过程的成岩作用序列。
2.4 形成环境及世代
葡萄石状白云石是沿C超微菱形的粒状白云石构成(50~100 μm)(图3)。金沙岩孔剖面中的灯影组中(样品JSYK-4)藻团粒云岩-皮壳-葡萄状白云石中的白云石呈多级次“球形-哑铃”及菱形等多种形态泥微晶白云石(图3a),围绕较大的藻球形呈平行的条带状或韵律纹层的超微白云石生长(图3b),具有典型溶液中生长台阶纹层,反映它在过饱和溶液沿C轴结晶生长,或溶解再沉积“拟晶化白云化”[17-18],即胶结物自泥晶化过程[11]。
对金石1井、沙湾范店灯影组中的葡萄-皮壳状白云岩、粉细晶、中细晶白云石以及缝洞中白云石共计58件样品的有序度(δ)分析结果表明: 4件葡萄-皮壳状、纤维状白云石的δ值分别为0.57,0.57 ,1.0和1.0。其中,2个较低δ值比向芳等[5]10件纤维状白云岩(δ=0.77~1.00)和张杰等[11](n=1,δ=1.00)均要低;也比向芳等(n=4,δ=0.70~1.00)和张杰等[11](n=4,δ=0.640~0.832)中的泥晶或粉细晶白云石有序度低。12件粉细晶白云岩(基质)δ值为0.38~1.00,据此可划分为3类:①δ≤0.50;② 0.50≤δ≤0.80;③ 0.80<δ≤1.0。围岩中白云石的结晶程度存在较大的差异。有3种或3种以上结晶程度不同的白云石。①原生或准同生云化产物;②浅埋藏白云岩化及重结晶作用产物;③经历了埋藏或热液改造下的白云岩化产物。葡萄-皮壳状白云石除于保留原始结构外后受成岩叠加改造(图5)。

图4 四川盆地灯影组皮壳、葡萄状云岩阴极发光(CL)及微区碳、氧同位素值Fig.4 The banding (fibrous and shrub textures) of grape-like dolomite under cathodluminescence microscopy and data of δ13C(PDB), δ18O(PDB) of micro-drill samples from the Dengying Formation in the Sichuan Basina.样品JSYK-016;b.样品FD-003;c.样品FD-20;d.样品JS1-2

图5 四川盆地灯影组基质、皮壳-葡萄状白云石与缝洞中白云石有序度分布Fig.5 Sequence of matrix dolomites,grape-like dolomites,vugs and fractures of the Dengying Formation in the Sichuan Basin
3 碳、氧和锶同位素
3.1 围岩及葡萄石状白云石
海相碳酸盐岩中的δ13C主要反映了非有机溶解的碳源,与产率、大洋循环、风化作用及外来的碳源有关。成岩过程中δ13C趋于减小,高的生物产率和有机碳埋藏速率能促进溶解碳酸盐和生物有机质之间的同位素分馏,使海水中溶解碳酸盐相对于生物有机质富集δ13C;反之。由于其波动变化曲线与生物地层或界线吻合较好,常被与用于全球碳循环相对应的对比及校验[19-21]。而影响氧同位素变化有同位素分馏、温度、交换反应、成岩改造等。
若葡萄石状白云石的先驱物是文石或镁方解石,经历了早期白云岩化后,可能已改变了稳定同位素组份;相对于同成因方解石,白云石δ18O(PDB)要高3‰~4‰[22]。为对比它与围岩及白云岩化过程的异同,需要获得不同剖面或钻井中同层段的围岩稳定同位素数据作为参考。统计中,需删除灯影组顶、底部极少量δ18O(PDB)小于-7.5‰;已证实其存在δ13C(PDB)正漂移和负漂移[15]。
林1井、金石1井、沙湾范店、金沙岩孔中灯影组粉泥晶、粉细晶云岩δ13C(PDB)平均为3.5‰(n=53),δ18O(PDB)平均为-4.3‰(n=53),87Sr/86Sr平均为0.710 000(n=23)。不同剖面略有所差异,详见表1。取白云岩基岩中δ13C(PDB)为3.5‰,δ18O(PDB)为4.3‰(对应方解石为0)作为原始海水参考值。
由表1和图6可见,不同剖面中、不同层段由于沉积亚微相及环境的差异,可能是由细菌或藻构成的藻席、灰泥丘等亚相的有机生物产率不同;δ13C也不同。δ13C(PDB)值呈金沙岩孔>金石1井>林1井>沙湾范店,这与岩石学观察相似;即发育富有机质的藻席、藻丘的层段中的皮壳-葡萄石状白云石相对更发育,对应δ13C(PDB)相对较高。
从皮壳-葡萄状白云石与基岩碳、氧同位素变化的关系性来看(图6):①金沙岩孔中δ13C(PDB),δ18O(PDB)值变化范围较大,皮壳-葡萄状白云石中δ13C(PDB),δ18O(PDB)既有正相关又有负相关的变化趋势,反映有大气淡水参于的复杂沉积-成岩流体;②沙湾范店中δ13C和δ18O相对变化较小,皮壳-葡萄状白云石中δ13C(PDB)和δ18O(PDB)有较弱的正相关,可能指示了海水与大气淡水混合带的特征;③金石1井中皮壳-葡萄状白云石δ13C(PDB)和δ18O(PDB)变化最小,两者不相关,反映其不受大气淡水的影响。
若按灯影组不同结构岩石类型的白云岩来划分:①泥粉-粉细晶白云岩:δ13C(PDB)平均为3.16‰(n=30,1.10‰~6.21‰),δ18O(PDB)平均为-4.46‰(n=30,-6.70‰~ -2.27‰),87Sr/86Sr平均为0.709 860(n=17,0.708 498~0.710 967);②细晶及少量中细晶白云岩:δ13C(PDB)平均为3.09‰(n=18,1.10‰~5.31‰),δ18O(PDB)平均为-3.35‰(n=18,-7.10‰~-0.71‰),87Sr/86Sr平均为0.710 170(n=7,0.708 953~0.711 931);③皮壳-葡萄状白云石(微区碳、氧同位素:δ13C(PDB)平均为3.20‰(n=24,1.10‰~4.74‰),δ18O(PDB)平均为-4.10‰(n=24,-8.10‰~-0.47‰),87Sr/86Sr平均为0.709 770(n=7,0.708 560~0.711 930)。
由此可见(图6),皮壳-葡萄状白云石δ13C(PDB)和δ18O(PDB)平均值略高于泥粉-粉细晶白云岩对应值;δ13C(PDB)随结晶程度增加,呈现出δ13C(PDB)平均值减小,受成岩改造影响;
皮壳-葡萄状白云岩与泥粉-粉细晶白云岩87Sr/86Sr值相似,稍高于当时海水87Sr/86Sr[24](0.708 700~0.709 400);但细晶及少量中细晶白云岩中的87Sr/86Sr增高明显,符合白云岩化过程的变化趋势。因此,皮壳-葡萄状白云石更多地保留早期海水-碳酸盐岩同位素交换的信息。皮壳-葡萄状白云石中87Sr/86Sr呈:金沙岩孔>金石1井>沙湾范店,反映了金沙岩孔受陆源或大气淡水影响较大(表2)。这与δ13C和δ18O得出的结论相似。

表1 四川盆地灯影组粉泥晶、粉细晶云岩碳、氧和锶同位素
注:“—”代表未分析或未提供。

图6 四川盆地灯影组皮壳-葡萄状与晶质白云岩碳、氧和锶同位素对比Fig.6 Comparison of δ13C(PDB), δ18O(PDB) and 87Sr/86Sr between matrix dolomite and grape-like dolomite of the Dengying Formation in the Sichuan Basina.δ13C(PDB)-δ18O(PDB)关系;b.δ13C(PDB)-87Sr/86Sr关系;c.δ18O(PDB)-87Sr/86Sr关系M.泥-粉晶白云岩基岩;G.皮壳-葡萄状白云岩
3.2 皮壳-葡萄石状白云石微区同位素
波纹状、明暗条带的皮壳状白云石的微区同位素分析表明:①样品JS1-3-1:δ13C(PDB)=4.38‰(白色纤状泥粉晶)~3.83‰(斑块粉细晶),δ18O(PDB)=-2.41‰(白色纤状泥粉晶)~-4.50‰(斑块粉细晶);②样品FD-008-1:δ13C(PDB)=2.1‰(暗色纤状泥粉晶)-1.4‰(灰白色粉细晶),δ18O(PDB)=-4.0‰(暗色纤状泥粉晶)~-5.40‰(灰白色粉细晶);③样品FD-011-2(黑色纤状泥晶):δ13C(PDB)=4.6‰,δ18O(PDB)=-2.42‰;④样品JSYK-009-1:δ13C(PDB)=6.2‰(白色纤状粉晶)~4.8‰(黑色纤状泥粉晶),δ18O(PDB)=-5.74‰(白色纤状粉晶)~1.27‰(黑色纤状泥粉晶);⑤样品JSYK-019-1:δ13C(PDB)=3.28‰(白色中细晶)~5.31‰(黑色纤状泥晶),δ18O(PDB)=-0.71‰(白色中细晶)~0.69‰(黑色纤状泥晶)。
微区δ13C(PDB)和δ18O(PDB)值有以下变化趋势:①黑色纤状泥粉晶比白色粒状白云石的δ13C(PDB)平均值要高,且δ18O(PDB)正偏或高正偏;反映了早期藻菌类或有机质微影响,不支持暗色体是经大气水残留交代的产物的认识[7];②粉细晶白云石环带比皮壳状中的黑、白色纤状的泥粉晶白云石δ13C(PDB)对应值低、δ18O(PDB)偏负,反映了两者在流体性质、有机质含量、温度等方面的差异;③不同剖面中的黑、白色纤状的泥粉晶白云石的δ13C(PDB)和δ18O(PDB)有一定的变化,但与对应剖面中的白云岩基质有相似的变化趋势。δ13C(PDB)值:岩孔剖面>金石1井>林1井>范店剖面,与上述不同剖面中的基质白云岩的δ13C(PDB)变化一致,反映出δ13C(PDB)受控于沉积亚微相及有机相的变化,另一方面也揭示了葡萄石状白云岩与基岩有相似沉积与成岩过程。
皮壳-葡萄状白云石微区碳氧同位素值分析结果表明:在JS1-2,FD-3,FD-20和JSYK-016样品中(图4,图6),δ13C(PDB)总体变化较平缓(一般<±1.5‰)。从核部至边缘略有升高的趋势,如样品JS1-2和FD-3;或稍有减小,如样品FD-20;或呈“之”字形变化(图7)。如样品JSYK-016和 FD-003的δ13C(PDB)低于基岩平均值;其他样品稍高于基岩δ13C(PDB)平均值(样品JSYK-016),但均接近于所在井段(或剖面)平均值,反映了皮壳-葡萄状白云石主要受海水控制。氧同位素变化较大、且较复杂。如样品JSYK-016和FD-3为先降后升;而样品FD-20中部为溶洞白云石,样品JSYK-016为粒状白云石充填;受大气淡水影响,但总体呈上升趋势。样品JS1-2两端低中间变化较平缓。与本区或剖面的岩的δ18O(PDB)平均值对比发现(表1):样品FD-003和FD-20偏正;样品JS1-2内、外壳负偏;而样品JSYK-016内部至核部正偏明显,这充分说明了葡萄石状白云岩氧同位素变化受海水控制为主、同时受又受后期大气水作用影响。
4 讨论
4.1 海水成岩发育的环境
海水成岩作用往往发育于高孔、高渗、较大的沉积水通量等环境。如大量海水成岩胶结作用易在如礁、潮间带等亚环境发育。海水中的方解石往往沿C轴生长为针状、纤状或皮壳-葡萄状,富含Mg,Sr,Na而贫Fe和Mn;一般不发光或暗淡。低镁/钙的大气淡水中,往往发育等轴粒状、叶片和等厚环边(结壳),在潜流带,处于还原带中Fe,Mn增加,发光性增加;若镁方解石矿物稳定化发育于大气淡水的开放体系中,会损失原始海水的微量或同位素特征;反之,与海水接触的封闭体系,会保留较重的碳、氧同位素值以及相对较高Sr,Na。
葡萄石状或皮壳状白云石多发育于潮坪的潮上-潮间带为主的富含菌藻的藻纹层、叠层石、藻凝块云岩。发育大量针状、纤状泥晶及放射状或束状的纤柱状泥粉晶白云石;等粒的镶嵌粒状、刀刃-犬牙状等也较常见,暗淡-桔红色为主韵律的阴极发光,据此推断,早期应为海水中的文石及高镁方解石或白云石的先驱物,同时受早期大气淡水作用的影响。
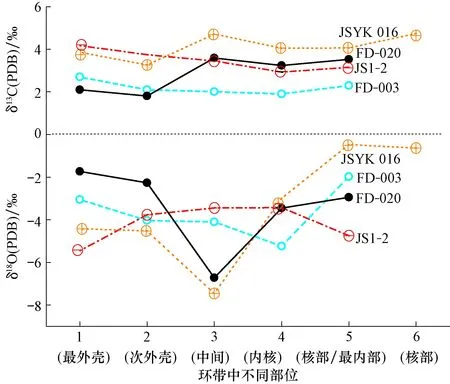
图7 四川盆地灯影组皮壳-葡萄状云岩(环带)微区碳、氧同位素变化Fig.7 Variation of δ13C(PDB) and δ18O(PDB) of micro-drill samples across the banding (fibrous and shrub textures) of grape-like dolomite in the Dengying Formation in the Sichuan Basin
4.2 与微生物岩成因关系
微生物岩是底栖微生物通过捕获、粘结或障积沉积物颗粒和(或)微生物自身矿化而形成的有机沉积构造[25]。如形成微生物藻席[26-27]埃及Solar Lake生物席之中驱动着球状和丝状蓝细菌的优势性变化,交互式的丝状蓝细菌占主导的暗色纹层与球状蓝细菌占优势浅色纹层的纹泥样式[28-29]。硫酸盐还原细菌(SRB)、硝酸盐还原、氨化作用以及甲烷厌氧细菌均可促进海底、天然气水合物环境中的白云石沉淀[30-31],可能起催化剂作用。
灯影组中早已报道了存在由肾形藻构成的灰泥丘[16]、微生物粘结丝带体的发现[1]为藻菌微生物的活动提供了进一步的证据。本次研究中,在金沙岩孔剖面中与藻纹层-团粒白云岩及叠层石共生的皮壳-葡萄状白云石广泛发育的明暗层-环带结构中发现了大量藻菌活动遗迹,虽然还不能完全确定离开藻的活动,就不能形成皮壳状及环带,后者可在远离平衡条件下、通过流体-岩石作用中自组织过程形成;但“球形-哑铃”的泥晶白云石等现象均揭示出微生物调制作用在皮壳-葡萄状白云石形成中起关键作用。
4.3 特殊的海水成分与白云石沉淀
原生白云石形成可能与元古代古海洋环境及化学性质有关,新元古代可能存在的“文石或白云岩海”[32-33]。发生于718~680 Ma的Sturtian全球的冰川事件促进了地壳风化作用、厌氧、海洋分层和滞流,减弱了海洋的流通,气候温暖。海水中Mg/Ca大于2,富含铁、铜金属元素及有机质[34-36];在碳酸盐岩过饱和环境下,可直接沉淀出白云石或先形成大量Mg-Ca水合物(一水方解石及白云石先驱物)[37],并与文石或白云石共生,并很块发生拟晶白云岩化而转为白云石[17,38,36],据此了解释了新元古早期的潮间-潮上带-藻云岩或叠层石中发育的卷曲-角砾状、网状-层状-片状的空隙及大量纤状白云石胶结物、示底构造、收缩微裂隙和去藻纹层化等多种复杂结构[36]。
葡萄石状或皮壳状白云石发育了与新元古早期十分相似的沉积与组构,可能指示了相对的低氧浓度(或较高的CO2分压), Mg/Ca或者碱度更高的相似的海洋环境,古甲烷菌等微生物作用较为普遍;与Sturtian期不同的是,它主要受651~635 Ma Marimoan(相当于南沱组)冰期影响,灯三段中的薄层凝灰岩层存在,虽指示了周缘火山喷发活动;但对灯二和灯一段影响较小,与热液来源有关的Co,Cu,Fe,Mn,Pb,Zn,S元素含量较低。另外,从灯影组剖面中,δ13C和δ34S总体相对稳定的向上减少变化,较高的有机生产率和向上增强氧化程度[23]也得到佐证。相对应地,葡萄石状或皮壳状白云石的发育程度向上减弱,这可能是金沙岩孔剖面的中、下段葡萄石状或皮壳状白云石相对发育的原因之一。
4.4 岩相学、地球化学数据及其解释中矛盾
正由于葡萄石状或皮壳状白云石与围岩中δ13C(PDB),δ18O(PDB)等值的差异,可能使部分学者得出大气水岩溶产物的认识。如向芳[5]中的皮壳-葡萄状白云石δ13C(PDB)=-1.190‰~0.671‰,δ18O(PDB)=-11.033‰~-6.684‰;施泽进等[10]δ13C(PDB)=-1.906‰~0.83‰(平均为-0.385‰,n=10),δ18O(PDB)=-11.033‰~-6.684‰(平均为-7.994‰,n=10)等。本次微区δ13C(PDB),δ18O(PDB)分析数据与上述部分学者提供数据有较大的差异,但与张杰等[11]及郝毅等[14]的提供的少量数据相似(表3)。矛盾的原因:①不同剖面或层段岩相存在差异,与基岩对比时,需考虑同一层位中的相似的沉积亚微相[21];②样品纯度不同,常规选样中,不可避免存在样品混合的现象,即胶结物与基岩难以完全分开;微区取样更纯、分析结果更可靠;③现代稳定同位素分析技术的进步,使原位、微区高精度分析成为可能。
4.5 成因模式探讨
综上所述,皮壳-葡萄石状白云岩中的明暗纹层、相向的等厚纤状胶结、暗淡-桔红色为主韵律的阴极发光特征、台阶式生长纹层,与泥粉晶云岩相似或稍为正偏的δ13C(PDB),δ18O(PDB)以及87Sr/86Sr值,均指示了海水成因为主的特征。但皮壳-葡萄石状白云岩具有穿层或沿断裂-裂隙及溶洞中发育,尤其是刀刃-犬牙状、镶嵌粒状胶结物以及“钙豆粒”的存在;反映了早期大气淡水作用影响;与葡萄石状白云石有关的缝洞充填中有中晶白云石、石英或残留孔洞,反映出葡萄石状白云石形成之后的成岩体系是相对开放的,具有叠加了构造-大气水或埋藏流体作用的地质与地球化学条件。
从皮壳-葡萄状白云石的多种产状、内部结构相似的环带状、孔洞中心充填物差异较大,从暗与明亮层阴极发光变化、暗色纤状比亮晶纤状的δ13C(PDB),δ18O(PDB)正偏等现象;反映了流体介质中咸水弱氧化与还原条件(Eh)规律改变,推断出以下的反应机制:“过饱和海水-微生物中细菌参于下成核-准稳态含镁矿物沉淀(AC)与快速转变-大气淡水渗流至非平衡态-过饱和沉淀(RFC-MC)”,即随着富藻类文石或少量高镁方解石沉淀,介质环境中pH,Eh和盐度等发生变化、生物活性下降,非有机高镁方解石逐渐进入过饱和,形成黑白相间韵律条带或环带白云石。

表2 四川盆地灯影组基质白云岩与皮壳-葡萄状白云石的有序度及碳、氧、锶同位素值对比
注:“—”代表未分析
5 结论
1) 皮壳-葡萄状白云石的基岩藻为-叠层石、凝块岩等微生物岩构成的藻席或灰泥丘等微生物岩发育有关。
2) 不同剖面中δ13C(PDB)和δ18O(PDB)是受原始沉积岩相及流体性质及其作用控制,环带中至少有3类的δ13C(PDB)和δ18O(PDB)具有不同变化趋势,主要反映了海水作用为主、但大气淡水参与环境。
3) 皮壳-葡萄状白云石在Marimoan(南沱组)冰期后氧化程度逐渐升高的海水中,通过 “过饱和海水-微生物中细菌参于下成核-准稳态含镁矿物沉淀(AC-RFC)形成-大气淡水渗入水岩反应形成粒状方解石(RFC-PC)与在浓缩海水中准同生云化(Dol)”中形成。
致谢:两位审稿人提出了中肯的意见,樊云鹤同志清绘了部分图件,在此一并感谢!
[1] 牟传龙,王秀平,梁薇,等.上扬子区灯影组白云岩葡萄体特征及成因初探—以南江杨坝地区灯影组一段为例[J].沉积学报,2015,33(6):1097-1110. Mou Chuanlong,Wang Xiuping,LiangWei,et al.Characteristics and genesis of grape-like stone of dolomite in Sinian DengYing formation in YangZe region:A case from the first section DengYing formation in YangBa,Nanjiang,Sichuan Province[J].Acta Sedimentologica Sinica,2015,33(6):1097-1110.
[2] 张荫本.震旦纪白云岩中的葡萄状构造成因初探[J].石油实验地质,1980,4:40-43. Zhang Yinben.Preliminary study on the botryoidal structures in the Sinian dolomites[J].Geology & Experiment,1980,4:40-43.
[3] 刘怀仁,刘明星,胡登新,等.川西南上震旦统灯影组沉积期的暴露标志及其意义[J].岩相古地理,1999,11(5):1-10. Liu Huairen,Liu Mingxing,Hu Dengxin,et al.The exposure indicators formed during the deposition of the Upper Sinian Dengying Formation in southwestern Sichuan and their significance[J].Sedimentary Geology and Tethyan Geology,1991.11 (5):1-10.
[4] 雷怀彦,朱莲芳.四川盆地震旦系白云岩成因研究[J].沉积学报,1992,10(2):69-77. Lei,Huaiyan,Zhu Lianfang .Study of origin of the Sinian algal and Non-algal dolomites in Sichuan basin[J].Acta Sedimentologica Sinica,1992,10(2):69-77.
[5] 向芳,陈洪德,张锦泉.资阳地区震旦系充填白云石与古岩溶[J].成都理工学院学报,1998,25(3):436-441. Xiang Fang,Zhang Jingquan,Chen Hongde.Filling dolostone and paleokarst of Sinian in ZiYang area[J].Journal of ChengDu Universtiy of Technology ,1998,25(3):436-441.
[6] 王兴志,穆曙,方少仙,等.四川盆地西南部震旦系白云岩成岩过程中的孔隙演化[J].沉积学报,2000,18(4):549-554. Wang Xingzhi,Mu Shuguang,Fang Shaoxian,et al.Evolution of porosity in the process of Sinian dolostone diagenesis in southwest Sichuan[J] .Acta Sedimentologica Sinica,2000,18(4):549-554.
[7] 陈明,许效松,万方,等.上扬子台地晚震旦世灯影组中葡萄状雪花状白云岩的成因意义[J].矿物岩石,2002,22(4):33-37. Chen Ming,Xu Xiaosong,Wan Fang,et al.Gensis and significance of grape and show flake-shaped dolomite from Dengying Formation of Upper Sinian in Upper Yangtze platform region[J].Journal Mineral and Petrology,2002,22(4):33-37.
[8] 方少仙,侯方浩,董兆雄.上震旦统灯影组中非叠层石生态系兰细菌白云岩[J].沉积学报,2003,21(1):96-105 Fang Shaoxian,Hou Fanghao ,Dong Zhaoxiong.Non-stromatoltite ecologic system cyanobacteria dolostone in Dengying Formation of Upper-Sinian[J].Acta Sedimentologica Sinica,2003,21(1):96-105.
[9] 黄文明,刘树根,张长俊,等.四川盆地震旦系储层孔洞形成机理与胶结充填物特征研究[J].石油实验地质,2009,31(5):449-461. Huang Wenming,Liu Shugen,Zhang Changjun,et al.The formation mechanisms of cavity and characteristics of fillings of Sinian system in Sichuan Basin[J].Petroleum Geology & Experiment,2009,31(5):449-461.
[10] 施泽进,梁平,王勇,等.川东南地区灯影组葡萄石地球化学特征及成因分析[J].岩石学报,2011,27(8):2263-2271. Shi Zejin,Liang ping,Wang yong.,et al.Geochemical characteristics and genesis of grapestone in Sinian Dengying Formation in south-eastern Sichuan basin[J].Acta Petrologica Sinica,2011,27(8):2263 -2271.
[11] 张杰,Brian Jones,潘立银,等.四川盆地震旦系灯影组葡萄状白云石成因[J].古地理学报,2014,16(5):715-725. Zhang Jie,Brian Jones,Pan Liyin,et al.Origin of botryoidal dolostone of the Sinian Dengying Formation in Sichuan Basin[J].Journal of Palaeogeography,2014,16(5):715-725.
[12] 王国芝,刘树根,李娜,等.四川盆地北缘灯影组深埋白云岩优质储层形成与保存机制[J].岩石学报,2014,30(3):667-678. Wang guozhi,Liushugen,Li Na,et al.Formation and preservation mechanism of high quality reservoir in deep burial dolomite in the DenYing formation on the Northern margin of the Sichuan[J].Acta Petrologica Sinica,2014,30(3):667-678.
[13] 周进高,姚根顺,杨光,等.四川盆地安岳大气田震旦系一寒武系储层的发育机制[J].天然气工业,2015,35(1):36-44 Zhou Jinggao,Yao Genshun,Yang Guang,et al.Genesis mechanism of the Sinian-Cambrian reservoirs in the AnYue Gas field Sichuan Basin[J].Nature Gas IND.,2015,35(1):36-44.
[14] 郝毅,周进高,陈旭,等.四川盆地灯影组“葡萄花边”状白云岩成因及地质意义[J].海相油气地质,2015,20(4):57-64 HaoYi,Zhou Jingao,ChenXu,et al.Genesis and geological significance of Upper Sinian DengYing dolostone with grape-lace shaped cement,Sichuan Basin[J].Marine Origin Petroleum Geology,2015,20(4):57-64
[15] 邓胜徽,樊茹,李鑫,等.四川盆地及周缘地区震旦(埃迪卡拉)系划分与对比[J].地层学杂志,2015,39(3):239-254 Deng Shenghui,Fan Ru,Li Xin ,et al.Subdivision and correlation of the Sinian(Ediacaran)system in the Sichuan Basin and its adjacent area[J].Journal of stratigraphy,2015,39(3):239-254
[16] 朱同兴,罗安屏.一个最古老的肾形藻泥丘相的首次发现及地质意义[J].岩相古地理,1992,12(4):20-28. Zhu Tongxing,Luo Anping.First discovery of an oldest renalcis moundfacies and its geological significance-an example from the Uppet Sinian Denying formation in Northeastern YunNan[J].Sedimentary facies and Palaeogeography,1992,12(4):20-28.
[17] Sibly D F,Dedoes,R E,Bartlett T R.Kinetics of dolomitization[J].Geology,1987,15,1112-1114.
[18] Sibly D F.Secular changes in the amount and texture of dolomite[J].Geology,1991,19(2):151-154.
[19] Kaufman A J,Knoll A H.Neoprototerozoic variation in the C-isotope composition of seswater:stratigraphic and biogeochemical implications[J].Precambrian Res.,1995,73:27-49.
[20] Des Maaris J.Isotopic evolution of the biogeochemical carbon cycle during the Precambrian.In:Valley JW,Cole DR.(eds.).Stable Isotope geochemistry Reviews in Mineralogy &geochemistry,2001,43:55-578.
[21] Jonathan A G,Malcolm W W.Facies-dependent δ13C variation from a Cryogenian platform margin,South Australia:Evidence for stratified Neoproterozoic oceans[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2009,271 :196-214.
[22] Land L S.the isotopic and trace element geochemistry of dolomite:the state of the art,in DH Zenger,J B Dunham,and R.L.Ethington,eds.s,Concepts and Modela of Dolomitization,Tulsa[J],1980,SEPM Special Publication28:87-110.
[23] 张同钢,储雪蕾,张启锐,等.扬子地台灯影组碳酸盐岩中的硫和碳同位素记录[J].岩石学报,2004,20(3):717-724 Zhang Tonggnag,Chu Xuelei,Zhang Qirui,et al.The sulfur and carbon isotopic records in carbonates of DengYing formation in the Yangtze platform,Chian[J].Acta Petrologica Sinica,2004,20(3):717-724.
[24] Halverson G P,Dud S F,Maloof A C,et al.Evolution of the87Sr /86Sr composition of Neoproterozoic seawater[J].Palaeongeography,Palaeoclimatology,Palaeoecology,2007,256(3-4):103-129.
[25] Perri E,Tucker M.Bacterial fossils and microbial dolomite in Triassic stromatolites[J].Geology,2007,35:207-210.
[26] Riding R.Microbial Carbonates:The geological record of calcified bacterial-algal mats and biofilms[J].Sedimentology,2000,47 (Suppl1):179-214.
[27] 梅冥相.微生物席的特征和属性:微生物席沉积学的理论基础[J].古地理学报,2014,16(3):285-304. Mei Mingxiang.Feature and nature of microbial-mat:Theoretical basis of microbial-mat sedimentology[J] .Journal of Palaeogeography,2014,16(3):285-304.
[28] Gerdes G.Structures left by modern microbial mats in their host sediments.In:Schieber J,Bose PK,Eriksson PG,et al (eds).Atlas of Microbial Mat Features Preserved within the Siliciclastic Rock Record[M].2007,Amsterdam:Elsevier,5-38.
[29] Gerdes G.What are microbial mats? [A].In:Seckbach J,Oren A(eds).Microbial Mats:Modern and Ancient Microorganisms in Stratified Systems.Cellular Origin,Life in Extreme Habitats and Astrobiology,2010,14[M].Berlin:Springer-Verlag,5-25.
[30] Vasconcelos C,Mckenzie J A,Bernasconi S,et al.Microbial mediation as a possible mechanism for natural dolomite formation at low temperatures[J].Nature,1995,377:220-222.
[31] Vasconcelos C,McKenzie J A.Microbial mediation of modern dolomite precipitation and diagenesis under anoxic conditions (Lagoa Verm elha,Rio de Janeiro,Brazil)[J].Journal of Sedimentary Research,1997,67(3):378-390.
[32] Hardie LA.Secular variations in Precambrian seawater chemistry and the timing of Precambrian aragonite seas and calcite seas[J].Geology,2003,31:785-788.
[33] Hood A S,Wallace M W,Drysdale R N.Neoproterozoic aragonite-dolomite seas? Widespread marine dolomite precipitation inCryogenian reef complexes[J].Geology,2011,39(9):871-874.
[34] Donald E C,Bernard P B,Alfonso M.The early diagenetic formation of organic sulfur in the sediments of Mangrove Lake,Bermuda[J].Geochimica et Cosmochimica Acta,1998,62(5):767-781.
[35] Giddings J A,Wallace M W.Sedimentology and C-isotope geochemistryof the ‘Sturtian’cap carbonate,South Australia[J].Sedimentary Geology,2009,216 (1-2):1-14.
[36] Hood A S,Wallace M W,Reedb,C P.et al.Enigmatic carbonates of the Ombombo Subgroup,Otavi Fold Belt,Namibia:A prelude to extreme Cryogenian anoxia[J]Sedimentary Geology,2015,324:12-31
[37] Rodriguez-Blanco J D,Shaw S B,Roncal H T,et al.The role of Mg in the crystallization of monohydrocalcite[J].Geochimica et Cosmochimica Acta ,2014,127,204-220.
[38] Higgins J A,Fischer W W,Schrag D P.Oxygenation of the ocean and sediments:consequences for the seafloor carbonate factory[J].Earth and Planetary Science Letters ,2009,284 (1),25-33.
(编辑 张玉银)
Applications of petrography and isotope analysis of micro-drill samples to the study of genesis of grape-like dolomite of the Dengying Formation in the Sichuan Basin
Qian Yixiong1,Feng Jufang2,He Zhiliang2,Zhang Keyin3,Jin Ting4,Dong Shaofeng1,You Donghua1,Zhang Yongdong1
(1.WuxiResearchInstituteofPetroleumGeology,PetroleumExploration&ProductionResearchInstitute,SINOPEC,Wuxi,Jiangsu214126,China;2.PetroleumExploration&ProductionResearchInstitute,SINOPEC,Beijing100083,China;3.Exploration&ProductionResearchInstitute,SouthwestOil&GasCompany,SINOPEC,Chengdu,Sichuan610081,China;4.No.2OilProductionFactory,JiangsuOilfieldCompany,SINOPEC,Jinhu,Jiangsu211600,China)
The shell-comb and grape-like dolomite of the Dengying Formation in the Sichuan Basin has an unusual textures.The petrographic data,carbon,oxygen and strontium isotope analysis on matrix and grape-like dolomites microdrill-samples have been used to investigate the origin of shell comb and grape-like dolomite.The grape-like dolomite is characterized by length-fast fascicular-optic fibrous aragonite cement,and bladed radial slow high-Mg calcites as well as granular calcites.The micritic dolomite in opposite direction growth,which consists of different size of either spherical and dumbbell forms or rhombic shape,has formed in pattern rhythm layers of banding or laminated along the terrace ofC-axis of crystal.The zonation,which consists of microcrystalline,fine crystalline dolomite,has generally a more distinctive negative values of δ13C(PDB) and δ18O(PDB) than that of the black and white fibrous dolomite of grape-like dolomite.The black or dull fibrous dolomite has a positive δ13C(PDB) and δ18O(PDB),compared with that of white fibrous dolomite.It may mainly indicate a marine origin for shell comb and grape-like dolomite.At least three different fluid environment oscillation to generate the banding of shell comb grape-like dolomite:① One of them is characteristic of lower negative of δ13C(PDB)and δ18O(PDB) and much higher 87Sr/86Sr compared to that of matrix dolomite or seawater.The wide medium bright orange luminescent bands with interval of dull bright zones is supposed to form in seawater overprinted with fresh water,indicate an periodic environment changes between weak oxidation and weak reduction;② The second has a sharp variation of sequences,with slow decrease of δ13C(PDB) and increase of δ18O(PDB) from its centre to rim of zonation.Normal seawater value of87Sr/86Sr,medium bright orange luminescent in the centre to dull or non-luminescent in outer zones indicates it generated in a seawater with continuous recharge of fresh water,and weak reduction in the center shifted to weak oxidation in the outer of zone;③ The third one does not have very clear zonation,and negative18O(PDB) in the rim with a slow increase of δ13C(PDB) and increasing and later decreasing of δ18O(PDB).It also shows a cathodoluminescence feature similar to that of matrix dolomite,which has been interpreted as a weak reduction and later oxidation environment.The environment first developed in condense seawater and was ventilated by fresh water in later diagenesis periods.In conclusion,the shell comb and grape-like dolomite has been developed in an unusual Neoproterzoic seawater characterized by very high Mg/Ca.It is coeval with the precipitation of algal laminated stromatolite and thrombolite,in forms of the microbial mat,mound,biofilms and reef.The environment model can most likely account for rapid growth of isopachous,fibrous aragonite and bladed high-Mg calcites cement,and growth of later granular calcites induced or promoted by bacterial microbial modulation,and para-contemporaneous dolomitization in condense seawater.The fluid is oversaturated seawater at the beginning,and later superimposed by a cyclic subaerial exposure of fresh water.
petrology,isotope of micro-drill samples,dolomite,Dengying Formation,Sichuan Basin
2016-11-03;
2017-04-05。
钱一雄(1962—),男,教授级高级工程师,碳酸盐岩沉积与储层。E-mail:qyx9167@vip.sina.com。
国家自然科学基金联合基金项目(U1663209);国家重点基础研究发展计划(973计划)项目(2012CB214802);国家科技重大专项(2011ZX05005-0023,2011ZX05008-003-11)。
0253-9985(2017)04-0665-12
10.11743/ogg20170404
TE122.1
A
猜你喜欢
工业炉(2023年6期)2023-03-29 05:29:18
河北地质(2022年2期)2022-08-22 06:23:14
黑龙江水利科技(2020年8期)2021-01-21 09:27:44
高校地质学报(2020年2期)2020-05-07 03:20:26
石油地质与工程(2019年3期)2019-09-10 08:27:42
趣味(语文)(2018年11期)2019-01-14 01:12:54
小学生作文(低年级适用)(2017年5期)2017-07-07 10:15:06
录井工程(2017年4期)2017-03-16 06:10:28
天然气勘探与开发(2015年3期)2015-12-08 08:28:25
安徽冶金科技职业学院学报(2015年3期)2015-12-02 03:46:39
