
黄土丘陵区撂荒群落地上生物量和物种多样性关系
2017-09-15 16:01王雁南
水土保持研究 2017年3期
高 艳, 杜 峰, 王雁南
(1.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100; 2.中国科学院大学,北京 100049; 3.西北农林科技大学, 陕西 杨凌 712100; 4.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100)
黄土丘陵区撂荒群落地上生物量和物种多样性关系
高 艳1,2, 杜 峰1,4, 王雁南3
(1.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100; 2.中国科学院大学,北京 100049; 3.西北农林科技大学, 陕西 杨凌 712100; 4.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100)
研究黄土丘陵区撂荒群落生物量与物种多样性的关系及其影响机制,有助于明确群落演变的机理,可对该区植被恢复重建、群落演替、生物多样性保护和维持具有重要意义。为此,选取该区5个典型撂荒群落,在群落尺度上,分析地上生物量的月动态,重点研究了地上生物量与物种多样性的关系。结果表明:(1) 5个撂荒群落地上生物量总体呈上升趋势,月份之间差异极显著,群落生物量由大到小为白羊草(Bothriochloaischaemum)群落>茭蒿(Artemisiagiraldii)群落>铁杆蒿(Artemisiasacrorum)群落>猪毛蒿(Artemisiascoparia)群落>达乌里胡枝子(Lespedezadahurica)群落;(2) 群落地上生物量与物种多样性关系表现为线性增加和线性减少两种形式。猪毛蒿群落中二者呈线性增加。铁杆蒿和白羊草群落地上生物量随丰富度的增加线性增加。白羊草和达乌里胡枝子群落地上生物量随均匀度的增加线性减少。空白生态位的占有和利用、生态位互补效应、种间相互作用是解释二者关系的内在机制,冗余种假说效应可能会导致不显著关系。
物种多样性; 地上生物量; 影响机制; 黄土丘陵区
物种多样性作为生物多样性的重要内容,是物种丰富度和均匀度的综合反映,是群落结构、组织水平、发展阶段、稳定程度和生境差异的综合体现[1-3]。群落生物量是群落功能的重要表现形式,与物种多样性有着共同的影响因素,如环境生产力、植被的结构及动态、人为干扰和群落组成种的相互作用等。生物多样性的变化,可通过影响区域植物群落组成、结构和关键种是否缺失进而影响到对资源的利用方式和效率,导致群落生物量发生变化[4-5]。群落生物量和物种多样性通常随着演替的进展呈上升状态,直至演替后期达到相对稳定[6]。外界干扰对二者的影响普遍认为在中度干扰下相对较大[7]。群落优势种的塑造作用及组成种的相互作用对二者关系的影响与环境生产力也有关。综上所述,物种多样性与生物量的关系受多种因素的影响,对其研究是群落生态学的关键问题,也一直是生态学领域的研究热点。
自然群落中影响物种多样性与生产力关系的因素不同,其主导生态学过程也会有所不同。两者关系的表现形式有线性、单峰或没有显著关系[8]。主要表现形式:(1) 群落生物量随多样性的增加呈线形增加。影响机制其一是空白生态位的占有与利用,即群落组成种对空闲资源的利用增加,因而群落生物量也会相应的呈线性增加[4,9],群落属性表现为盖度较低,植被处在演替前期;其二可能是生态位互补效应[10-11],在时间、空间和资源类型上具有不同生态位需求的群落组成种各自利用不同的资源,整体上群落种丰富度的增加使得组成种对资源的利用能力也在增加,因而生物量也呈增加趋势,群落属性表现为盖度高,环境生产力高;其三是群落组成种的相互促进作用或种间正相互作用[12-13]。在严酷环境条件下,植物为应对环境胁迫而聚集生长。群落属性表现为聚集度高,斑块状生长,对土壤具有沃岛效应[14-15]。(2) 群落生物量随种多样性的增加线性减少。这种关系不常见,影响机制是优势种对其他种的竞争排除,且对生物量的贡献较大。优势种通常具有无性繁殖能力,种内聚集度高,丛生或斑块状生长,对其他种的竞争抑制强[16-17]。(3) 单峰关系[4,18-21]。即在中等生产力水平多样性最大,该假说是建立在竞争排斥原理的基础上,可以预测在有限资源的基础上竞争共存植物种的数量。(4) 生物量与物种多样性没有显著关系[22]。在小空间尺度上,单峰关系出现的相当频繁,占所有研究的41%~45%[4,8,23]。但目前,对3种关系格局的看法和解释仍存在很大的争议[24-25],在Waide等[26]的调查中,有32%关系不显著。所以还需在更广泛的领域开展研究,以解释各种关系格局形成的自然背景和机制。我国物种多样性与生产力关系的研究大多集中于典型草原生态系统[7,27-30],目前针对黄土丘陵区植物群落物种多样性与生产力的关系及其机制研究较为缺乏。
黄土丘陵区近些年环境得到一定的改善,但是生态系统依然较为脆弱,研究物种多样性与生物量的关系,对该区植被演替和恢复、维持生物多样性稳定和生态系统健康具有重要意义[31]。以往研究中,大多是生产力为因,多样性为果。生产力本是生产者,多样性反映生产者的一种属性,理应是因,生产力则应是果[17]。为此,我们以黄土丘陵区安塞县高桥乡为研究单元,以多样性为因、生物量为果,以群落尺度选取5个典型撂荒群落(猪毛蒿群落、铁杆蒿群落、茭蒿群落、白羊草群落、达乌里胡枝子群落),分析其地上生物量和物种多样性的关系。探讨二者的关系及影响机制,理解不同群落的塑造、演替机理,及植物多样性对黄土丘陵区生态系统功能的作用,为该区脆弱的环境的恢复和重建提供一定的决策依据。
1 材料与方法
1.1 研究区概况
研究区位于黄土丘陵区陕北安塞县高桥乡,其地理位置在东经109°11.168′—109°12.585′,北纬36°39.143′—36°40.226′,海拔高度为1 150~1 300 m。年平均日照时数为2 300~2 570 h,年平均日照总辐射量为497.7~553.4 kJ/cm2,年平均降水量为490.5~663.3 mm,其中6—8月降雨量占全年降水的60%~80%,年平均气温7.7~10.6℃,≥0℃积温3 733.1℃,≥10℃积温3 170.3℃,地带性土壤为黄绵土。地带性植被为森林草原,调查地区草本建群种有白羊草(Bothriochloaischaemum)、达乌里胡枝子(Lespedezadahurica)、铁杆蒿(Artemisiasacrorum)、猪毛蒿(Artemisiascoparia)、茭蒿(Artemisiagiraldii)、长芒草(Stipabungeana)和硬质早熟禾(Poasphondylodes)等。
1.2 群落调查
根据撂荒群落的勘查结果,选定在无人为干扰的21个撂荒群落样地进行群落调查。选取猪毛蒿群落、铁杆蒿群落、茭蒿群落、白羊草群落和达乌里胡枝子群落,5个群落分别选取9,5,2,3,2个样地(样地编号及其概况见表1)。调查时间为2003年、2005年,其中2003年是5—9月,每月每个样地随机取2个1 m×1 m典型样方。2005年是在生长旺季(8月、9月)调查,每月每个样地随机取3个1 m×1 m典型样方,共6个样方。测量样地内每种植物的盖度、高度、多度和地上生物量。
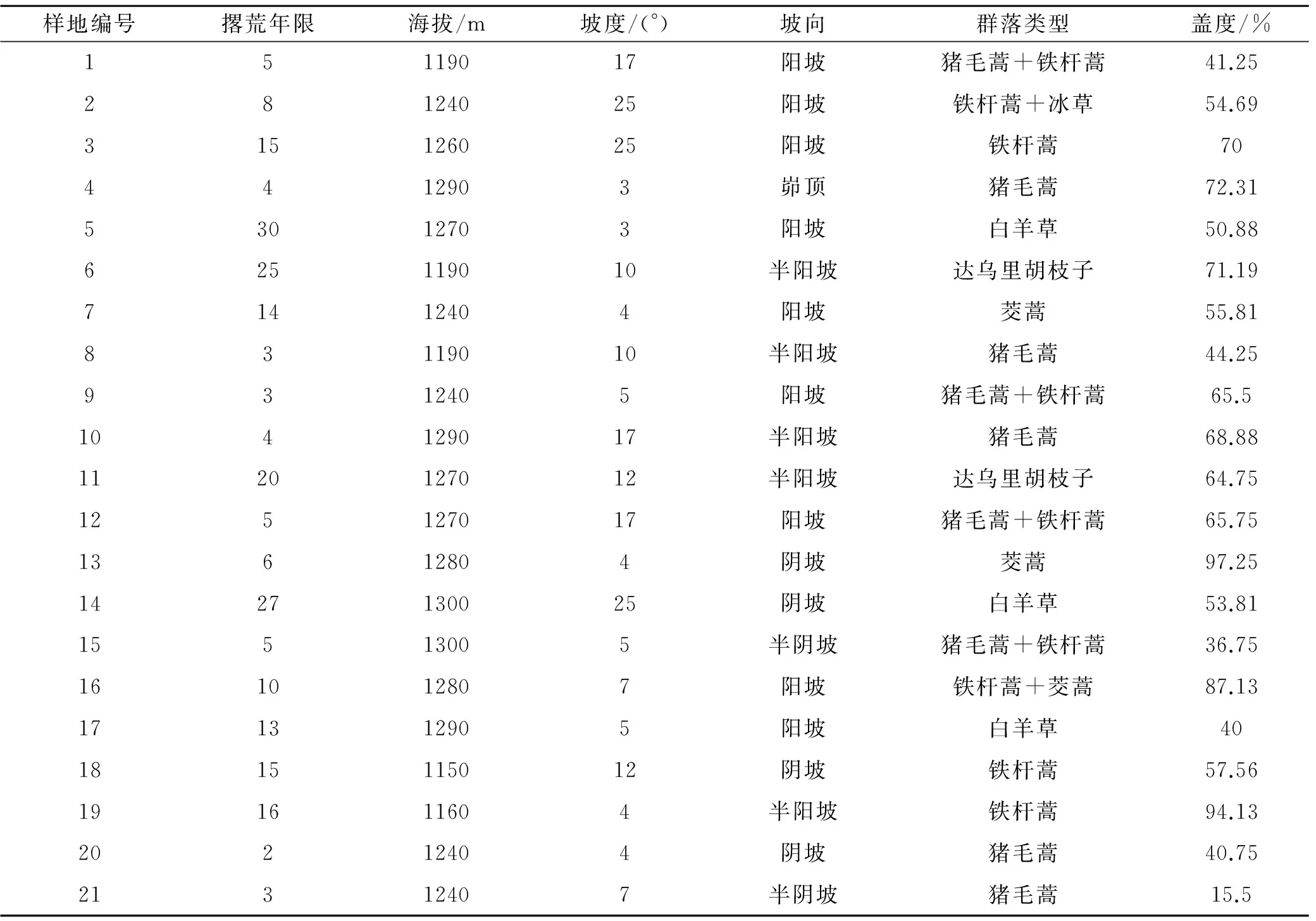
表1 样地概况
注:摞荒年限是2003年与农户走访调查数据。
1.3 数据分析
对摞荒群落进行地上生物量的月动态分析(用2003年群落调查数据),及地上生物量和多样性的关系分析(用2003年、2005年植物生长旺盛期群落调查数据)。各样方首先分种计算相对重要值,并计算多样性。选择以下多样性指数进行计算:Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数。
Shannon-Wiener指数(Hi):
Hi=-∑PilnPi
(1)
Margalef丰富度指数(Di):
(2)
Pielou均匀度指数(Ei):
Ei=Hi/lnS
(3)
式中:Pi为样方内i种的相对重要值;S代表样方内物种数目;N代表样方内物种个体数;相对重要值(Pi)=(相对盖度+相对高度+相对多度+相对生物量)/4。
采用Origin 8.0软件对群落地上生物量和物种多样性的关系进行拟合,选择剩余残差最小和显著性、拟合度最高且较为简单的曲线作为最终结果。使用SPSS 20.0进行数据统计分析。各群落不同月份生物量进行单变量二因素方差分析,使用最小显著差异法(LSD)进行多重比较。生物量和多样性的拟合使用F检验,二者相关性分析使用非参数Kendall的tau-b分析。
2 结果与分析
2.1 撂荒群落地上生物量月动态
2.1.1 不同月份各群落生物量的变化趋势 所有群落地上生物量在月份之间有极显著差异(p<0.001),生长季5—9月的总地上生物量均值分别为137.26,109.16,99.90,132.10,162.58 g/m2,呈上升趋势。各群落生物量在月份之间差异极显著(p<0.001),由大到小为白羊草群落(375.86 g/m2)>茭蒿群落(182.42 g/m2)>铁杆蒿群落(91.79 g/m2)>猪毛蒿群落(76.1 g/m2)>达乌里胡枝子群落(51.49 g/m2)。猪毛蒿、铁杆蒿群落地上生物量均呈上升趋势,9月份达到最大值。达乌里胡枝子群落变化平稳,7月增幅最大。白羊草、茭蒿群落的地上生物量总体呈下降趋势(图1)。
2.1.2 同月份间群落生物量的差异 5月、8月各群落生物量差异显著(p<0.001),其中白羊草群落最高,而6月、7月、9月3个月份各群落生物量差异不显著(p>0.05)。5月份白羊草群落的生物量分别是猪毛蒿群落、铁杆蒿群落、达乌里胡枝子群落、茭蒿群落的7.04,4.09,4.94,2.06倍,8月份分别是3.01,2.81,2.90,3.19倍(图1)。
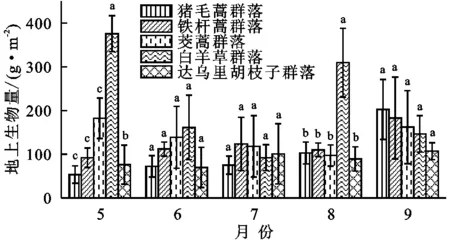
图1撂荒群落地上生物量月动态
2.2 不同撂荒群落地上生物量与物种多样性关系
图2表明撂荒群落地上生物量与物种多样性指数关系表现为线性增加和线性减少2种,不同群落的变化趋势差异显著。表2所示群落地上生物量由大到小为白羊草群落>铁杆蒿群落>茭蒿群落>猪毛蒿群落>达乌里胡枝子群落。铁杆蒿、茭蒿群落的物种多样性最高,白羊草群落次之,猪毛蒿、达乌里胡枝子群落最低。
由图2,表3可知,猪毛蒿群落的地上生物量随多样性指数的增加呈极显著的线性增加(p<0.01)。铁杆蒿群落地上生物量与丰富度指数呈极显著的线性增加(p<0.01),达乌里胡枝子群落其与均匀度指数呈显著的线性减少(p<0.05)。白羊草群落地上生物量随丰富度的增加是线性增加(p<0.01),随均匀度的增加线性减少(p<0.05)。茭蒿群落地上生物量与物种多样性关系不显著。
由表4可知,各个群落地上生物量与物种多样性指数的相关性差异较大。猪毛蒿群落二者呈极显著的正相关关系(p<0.01)。铁杆蒿和茭蒿群落地上生物量与Pielou指数呈显著的正相关关系,与Margalef指数没有相关关系。白羊草群落中,地上生物量与Pielou指数呈极显著的负相关关系(p<0.01),与Margalef指数是显著的正相关关系(p<0.05)。达乌里胡枝子群落中二者没有相关关系。
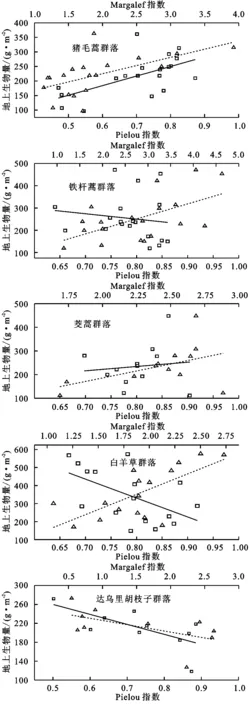
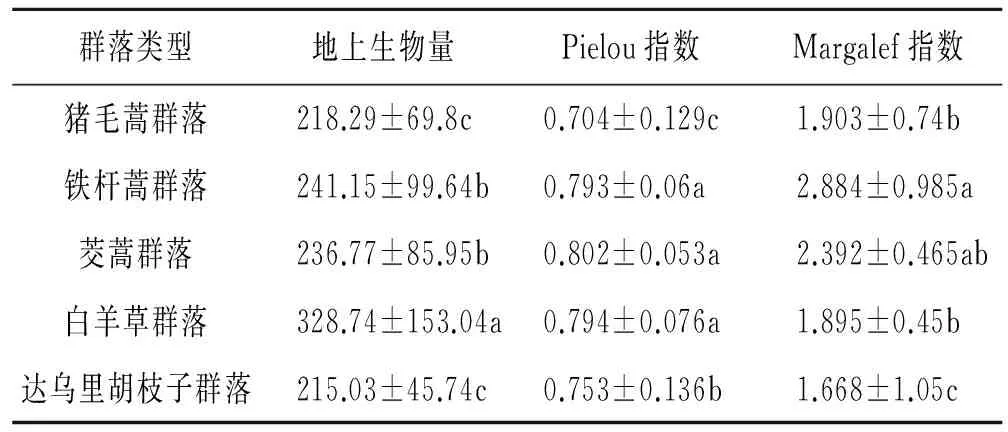
图2 典型群落地上生物量与物种多样性关系表2 5个典型群落的生物量与多样性指数
注:不同小写字母表示差异显著性。
3 讨 论
3.1 撂荒群落地上生物量的月动态
群落生物量是区域生态系统功能的物质基础,对区域初级、次级生产具有重要的指导意义。影响生物量的三大因子是气候、土壤、植被。调查样地中,区域气候背景一致、耕地弃耕前耕作条件相对均一,相对匀质的黄土母质,气候和土壤因素对二者关系影响较小。其中,水热条件的综合作用,决定了生长季不同阶段植物的生长发育[32]。
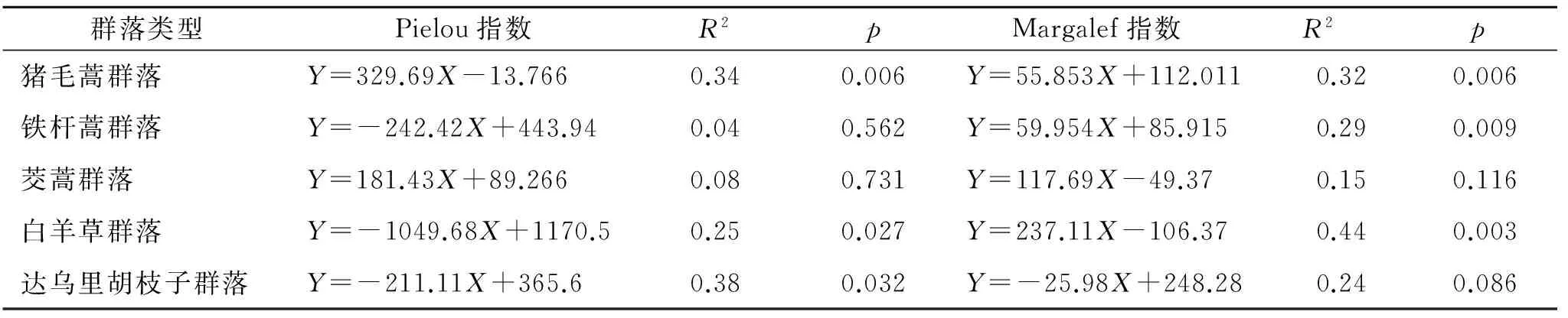
表3 典型群落的地上生物量与多样性指数拟合方程
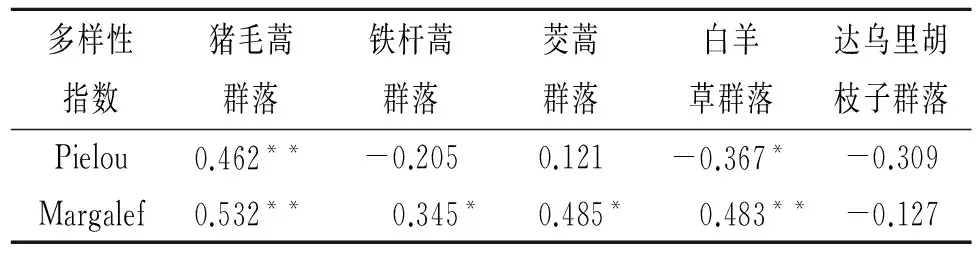
表4 典型群落地上生物量与种多样性指数之间的相关性分析
注:*代表显著水平是0.05,**代表显著水平是0.01。
本研究中地上生物量月动态变化显著。生长季初期,植物主要表现为营养生长,地上生物量低;生长季后期,植物开花结实,地上生物量明显增加,植物竞争强度增大,与初期相比变化明显。猪毛蒿群落在生长季初期,群落结构简单,生物量较低,但是群落在整个生长季期间总体盖度低,大量新个体利用空白生态位产生,繁殖率高,生物量呈不断上升趋势;铁杆蒿群落的地上生物量具有明显的季节性变化,在9月下旬达到峰值。由于铁杆蒿分枝多、根蘖性强,可从母株产生大量新株,易繁殖,在生长季前期主要表现为营养生长[33];达乌里胡枝子群落的地上生物量与黄土区其他植物群落相比明显较低。原因可能是样地位于阳坡、半阳坡,光照强,土壤含水量较低,为了适应干旱的立地条件做出的生理生态响应,总体地上生物量最低,与朱志诚等[34]的研究相一致;白羊草群落根系分布浅,在地上地下30 cm左右空间形成优势,土壤养分利用率高,总体生物量高。大多生长于阳坡,随着阳坡的干旱环境及放牧强度增加,可能导致6月、7月份生物量下降,8月份降雨量大,生长迅速[35]。茭蒿群落在环境资源充足、种内种间竞争尚不明显时迅速定居成为优势种,但是群落盖度较低,生物量低,竞争力弱。随着水热条件的改善、伴生种的增加,存活率下降,生物量下降[36]。
3.2 不同撂荒群落地上生物量与物种多样性关系机理
一般来说,区域尺度上生物量大体代表了环境生产力,物种多样性具有不同的变化规律。物种多样性与群落初级生产力的关系受到土壤水分、植被类型、海拔高度、人为干扰、演替阶段、生境条件差异、资源可利用性、种内种间竞争等因素的影响[2,11,37-39]。但在黄土丘陵区的群落小尺度上,群落内部的气候、土壤条件、撂荒期间的利用方式及撂荒历史等外部环境因素是相对均一的,二者关系变化的主要是由群落结构、物种组成和动态等内部因素影响[24-25,40]。
在撂荒演替的初期,群落环境水平较差,物种组成简单,多样性差,群落盖度较低,生物量较低。本文中猪毛蒿群落即处于演替恢复初期,盖度大约为40%~65%,物种数仅6~10种,演替中后期物种如铁杆蒿、达乌里胡枝子、冰草等开始出现并占有、利用大量的空白生态位,成为群落的主要次生种。随着群落物种数和个体数的不断增加,对空闲资源的利用增加,生物量也相应增加。因此对空白生态位的占有和利用是其主要影响机制。
铁杆蒿群落是在茭蒿群落演替3~4 a后,两群落均属撂荒演替中期典型群落,物种多样性指数最高,与郝文芳对黄土丘陵区物种多样性在演替中期最高一致[20]。铁杆蒿群落样地盖度平均在70%以上,物种数大致为20~25种,具有不同生态位需求的群落组成种各自利用不同的资源,丰富度的增加使得组成种对资源的利用能力也在增加,茭蒿、冰草等生物量较高的物种成为主要次生种,因而生物量也是增加的。生态位互补效应是主导生态学过程。以往的研究中,茭蒿群落处于黄蒿群落演替1~2 a后,环境相对较好条件下物种大致为15~25种,物种丰富度、均匀度高,主要次生种是生物量较低的铁杆蒿、达乌里胡枝子等[32,41]。本研究中茭蒿群落与铁杆蒿群落为演替序列同期,甚至一些样地演替较慢,样地盖度<60%,一般成熟茭蒿群落盖度是60%~75%[42],说明此群落为不稳定不成熟群落,种内种间竞争尚不明显,地上生物量与种多样性关系尚不显著。
白羊草和达乌里胡枝子为演替后期耐旱性较强物种,随着土壤的旱化,灌木比草本的根系更深、更发达[42],资源竞争力更强。本研究中白羊草群落主要分布在海拔较高的阳坡(海拔>1 270 m),种间正的相互作用导致地上生物量随物种数(丰富度)的增加而增加,白羊草在群落中所占的地位达到60%以上,竞争引起的自疏及他疏效应可能导致均匀度开始下降,且演替后期土壤水分减少,地上生物量随之下降;达乌里胡枝子群落主要分布在阳坡、半阳坡,由于其自身生物学特性决定的地上生物量较低,干旱条件下地下生物量明显高于地上生物量。物种数开始减少,由资源利用水平引起的种内种间竞争增大[25-29],地上生物量与多样性均呈下降趋势,是为了应对环境胁迫采取的策略。
由此可知,群落尺度上,地上生物量与物种多样性的关系主要由优势种的作用和演替阶段影响,可对群落生态演替及生态维护有一定的帮助。本文二者没有关系可能与各群落组成种对生物量的贡献有关。通常情况下优势种优势度越大,对生物量贡献越大,其他种的增加对生物量的贡献较小甚至是冗余的,即冗余种假说机制[16-17,22],即群落在特定的环境条件下物种数达到饱和,但随演替过程物种数的增加,丰富度和均匀度都变大,但对生物量贡献微弱,导致二者关系不显著。生物多样性本身不是一个独立变量,受到多种因素的影响,也影响着生态系统的维持和功能。本文仅仅在群落小尺度上浅谈二者的关系及影响机制,未涉及环境因素和地下部分。将来的研究还需综合考虑环境、人为干扰等因素的影响,以及在更大的空间尺度上进行,比如黄土高原草原、森林生态系统、流域等尺度,可能会得到更有普遍意义的结果。
4 结 论
(1) 撂荒群落在群落尺度上,地上生物量的变化由水热条件综合决定。不同群落的地上生物量大小为白羊草群落>茭蒿群落>铁杆蒿群落>猪毛蒿群落>达乌里胡枝子群落。
(2) 二者关系主要表现为线性增加和线性减少两种形式。在群落尺度上,地上生物量与物种多样性的关系主要受优势种的作用和演替阶段影响。
(3) 对空白生态位的占有和利用,影响猪毛蒿群落地上生物量与物种多样性关系的主要生态学过程;生态位互补效应是影响铁杆蒿群落二者关系的主要生态学过程;种间对资源利用竞争作用是影响白羊草群落和达乌里胡枝子群落二者关系的主要生态学过程;茭蒿群落尚处于不稳定阶段,二者关系尚不显著。
[1] Ruijven J V, Berendse F. Diversity-productivity relationships: Initial effects, long-term patterns, and underlying mechanisms[J]. Proceedings of the National Academy of Sciences,2005,102(3):695-700.
[2] Day M, Baldauf C, Rutishauser E, et al. Relationships between tree species diversity and above-ground biomass in Central African rainforests: implications for REDD[J]. Environmental Conservation,2013,41(1):64-72.
[3] Huston M A. Biological Diversity: the Coexistence of Species[M]. London: Cambridge University Press,1994.
[4] 漆良华,彭镇华,张旭东,等.退化土地植被恢复群落物种多样性与生物量分配格局[J].生态学杂志,2007,26(11):1697-1702.
[5] Tilman D. Resource Competition and Community Structure[D]. Princeton: Princeton University Press,1982.
[6] Tilman D. Causes, Consequences and ethics of biodiversity[J]. Nature,2000,405(6783):208-11.
[7] 杨殿林.呼伦贝尔草原群落植物多样性与生产力关系的研究[D].呼和浩特:内蒙古农业大学,2005.
[8] Rajaniemi T K. Explaining productivity-diversity relationships in plants[J]. Oikos,2003,101(3):449-457.
[9] Dodson S I, Arnott S E, Cottingham K L. The relationship in lake communities between primary productivity and species richness[J]. Ecology,2000,81(10):2662-2679.
[10] Mulder C P H, Uliassi D D, Doak D F. Physical stress and diversity-productivity relationships: the role of positive interactions.[J]. Proceedings of the National Academy of Sciences of the United States of America,2001,98(12):6704-6708.
[11] Hector A, Schmid B, Beierkuhnlein C, et al. Plant diversity and productivity experiments in european grasslands[J]. Science,1999,286(5442):1123-1127.
[12] Lambers J H R, Harpole W S, Tilman D, et al. Mechanisms responsible for the positive diversity-productivity relationship in Minnesota grasslands[J]. Ecology Letters,2004,7(8):661-668.
[13] 徐瑾.种间正相互作用机理及其对物种特性差异响应的模拟与实验研究[D].兰州:兰州大学,2009.
[14] 瞿王龙,杨小鹏,张存涛,等.干旱、半干旱地区天然草原灌木及其肥岛效应研究进展[J].草业学报,2015,24(4):201-207.
[15] 熊小刚.生态学中的新领域:沃岛效应与草原灌丛化[J].植物杂志,2003(2):45-46.
[16] 黄建辉,白永飞,韩兴国.物种多样性与生态系统功能:影响机制及有关假说[J].生物多样性,2001,9(1):1-7.
[17] 彭少麟,黄忠良.生产力与生物多样性之间的相互关系研究概述[J].生态科学,2000,19(1):1-9.
[18] Kassen R, Buckling A, Bell G, et al. Diversity peaks at intermediate productivity in a laboratory microcosm[J]. Nature,2000,406(6795):508-12.
[19] Abramsky Z, Rosenzweig M L. Tilman's predicted productivity-diversity relationship shown by desert rodents[J]. Nature,1984,309(5964):150-1.
[20] 郝文芳,梁宗锁,陈存根,等.黄土丘陵区弃耕地群落演替过程中的物种多样性研究[J].草业科学,2005,22(9):1-8.
[21] 韩大勇,杨永兴,杨杨.若尔盖高原退化沼泽群落植物多样性及种间相关性沿排水梯度的变化[J].植物生态学报,2012,36(5):411-419.
[22] Huston M A, Aarssen L W, Austin M P, et al. No consistent effect of plant diversity on productivity[J]. Science, 2000, 289(5483):1255-1255.
[23] Leibold M A. Biodiversity and nutrient enrichment in pond plankton communities[J]. Evolutionary Ecology Research, 1999.1(1):73-95.
[24] Fridley J D. Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities[J]. Oecologia, 2002, 132(2):271-277.
[25] Guo Q F, Berry W L. Species richness and biomass: Dissection of the hump-shaped relationships[J]. Ecology, 1998,79(7):2555-2559.
[26] Waide R B, Willig M R, Steiner C F, et al. The Relationship Between Productivity and Species Richness[J]. Ecology, Evolution, and Systematics, 1999, 30(30):257-300.
[27] 杨利民,周广胜,李建东.松嫩平原草地群落物种多样性与生产力关系的研究[J].植物生态学报,2002,26(5):589-593.
[28] 郭轶瑞,赵哈林,赵学勇,等.科尔沁沙质草地物种多样性与生产力的关系[J].干旱区研究,2007,24(2):198-203.
[29] 王长庭,龙瑞军,王启基,等.高寒草甸不同草地群落物种多样性与生产力关系研究[J].生态学杂志,2005,24(5):483-487.
[30] 臧岳铭.高寒矮嵩草草甸物种多样性、功能多样性与初级生产力关系的研究[D].西安:陕西师范大学,2009.
[31] 周萍,刘国彬,侯喜禄.黄土丘陵区不同坡向及坡位草本群落生物量及多样性研究[J].中国水土保持科学,2009,7(1):67-73.
[32] 朱志诚,贾东林.黄土高原中部草地群落初级生产[J].西北大学学报:自然科学版,1998,28(6):536-539.
[33] 朱志诚,贾东林.陕北黄土高原铁杆蒿群落生物量初步研究[J].生态学报,1993,13(3):243-251.
[34] 朱志诚,贾东林.达乌里胡枝子群落生物量初步研究[J].中国草地学报,1994(3):25-25.
[35] 朱志诚,贾东林.陕北黄土高原白羊草群落生物量初步研究[J]. 植物学报,1992,34(10):806-808.
[36] 刘芳,杨劼,宋炳煜,等.内蒙古鄂尔多斯黄土丘陵沟壑区茭蒿种群结构及群落特征[J].生态学杂志,2010,29(9):1685-1690.
[37] Oba G, Vetaas O R, Stenseth N C. Relationships between biomass and plant species richness in arid-zone grazing lands[J]. Journal of Applied Ecology, 2001,38(4):836-845.
[38] 刘江华,徐学选.黄土丘陵区小流域植物群落物种多样性空间分布特征[J].陕西林业科技,2009(1):9-12.
[39] 杜峰,梁宗锁,徐学选,等.陕北黄土丘陵区撂荒群落土壤养分与地上生物量空间异质性[J].生态学报,2008,28(1):13-22.
[40] Mittelbach G G, Gough L. What is the Observed Relationship between Species Richness and Productivity[J]. Ecology,2003,84(12):3384-3390.
[41] 杜峰.陕北黄土丘陵区撂荒演替及主要植物种内、种间竞争研究[D].陕西杨凌:西北农林科技大学,2004.
[42] 朱志诚,黄可,李继瓒.陕北黄土高原森林地带草本植物群落类型及其动态特征[J].中国草地学报,1989(3):18-24.
RelationshipBetweenAbovegroundBiomassandSpeciesDiversityofAbandonedCommunityinLoessHillyRegion
GAO Yan1,2, DU Feng1,4, WANG Yannan3
(1.InstituteofSoilandWaterConservation,CAS&MWR,Yangling,Shaanxi712100,China; 2.UniversityofChinaAcademyofSciences,Beijing100049,China; 3.NorthwestA&FUniversity,Yangling,Shaanxi712100,China; 4.InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling,Shaanxi712100,China)
The study on relationship between aboveground biomass and species diversity of abandoned old-field community and its mechanism in the hilly region of the Loess Plateau is helpful for understanding community succession process and is also important for vegetation reconstruction, biodiversity protection, and ecosystem health maintenance. In this study, five typical communities were selected to investigate the monthly dynamics of aboveground biomass and its correlation with biodiversity at community scale. The results showed that: (1) the aboveground biomass of the five communities increased from May to September and the difference between different months were significant (p<0.01); the community aboveground biomass was the highest inBothriochloaischaemumcommunity (375.86 g/m2), followed byArtemisiagiraldii(182.42 g/m2),Artemisiasacrorum(91.79g/m2),Artemisiascoparia(76.1 g/m2) communities, the lowest biomass was observed in Lespedeza dahurica (51.49 g/m2) community; (2) linearly positive and negative relations were found with respect to the relationships of aboveground biomass and species diversity index; inArtemisiascopariacommunity, aboveground biomass increased linearly with increase of species diversity index, while no relation was found in theArtemisiagiraldiicommunity; the aboveground biomass increased linearly with increase of Margalef index inBothriochloaischaemumandArtemisiasacrorumcommunities, and decreased linearly with increase of Pielou index inBothriochloaischaemumandLespedezadahuricacommunities. Furthermore, the influencing mechanism for the relation between aboveground biomass and biodiversity may result from occupation and utilization of blank niche, niche complementarity, interspecific interactions and competition and exclusion effect. The redundant species hypothesis perhaps resulted in the insignificant relations.
species diversity; aboveground biomass; influencing mechanism; Loess Hilly Region
2016-05-09
:2016-05-29
国家自然科学基金“黄土丘陵区植物竞争对土壤水分变化的响应”(41271526)
高艳(1989—),女,山西吕梁人,硕士研究生,研究方向为植物生态。E-mail:gaoyan923329@126.com
杜峰(1971—),男,内蒙古乌兰察布人,博士,副研究员,主要从事群落、种群生态及植被恢复生态研究。E-mail:dufeng@ms.iswc.ac.cn
Q145.2
:A
:1005-3409(2017)03-0096-07
猜你喜欢
园艺与种苗(2021年8期)2021-12-05
甘肃科技(2020年21期)2020-04-13
河南农业(2019年12期)2019-12-06
河南农业·综合版(2019年12期)2019-09-10
种子(2019年4期)2019-05-28
天然产物研究与开发(2018年10期)2018-11-06
意林·少年版(2018年16期)2018-09-12
中国民族民间医药(2018年13期)2018-07-31
中国民族医药杂志(2015年11期)2015-07-12
小火炬·阅读作文(2009年11期)2009-11-30
