
烤烟几个主要根系生理特性的遗传效应分析
2017-09-12 10:22:54杨丙钊马文广彭玉富孙志浩杨铁钊
中国烟草科学 2017年4期
许 杰,杨丙钊,马文广,彭玉富,孙志浩,赵 科,杨铁钊*
(1.河南农业大学烟草学院,郑州 450002;2.河南中烟工业有限责任公司,郑州 450016;3.玉溪中烟种子公司, 云南 玉溪 653100)
烤烟几个主要根系生理特性的遗传效应分析
许 杰1,杨丙钊2,马文广3,彭玉富2,孙志浩1,赵 科1,杨铁钊1*
(1.河南农业大学烟草学院,郑州 450002;2.河南中烟工业有限责任公司,郑州 450016;3.玉溪中烟种子公司, 云南 玉溪 653100)
采用盆栽试验,选用5个烟叶钾含量不同的烤烟品种,按照Griffing Ⅱ组配成15个双列杂交组合,研究了充足供钾条件下烤烟旺长期4个主要根系生理特性的遗传效应。结果表明,4个根系生理性状在不同基因型间差异显著,一般配合力和特殊配合力方差也达到极显著水平,广义遗传率较高,均大于60%,性状的变异主要由基因效应控制。其中,烤烟根体积和根系阳离子交换量(CEC)的遗传以基因的加性效应为主,狭义遗传率较高,提高烤烟根系量和根系CEC育种在早代进行选择效果较好;烤烟根系活力和ATP酶活性的遗传以基因的显性效应为主,F1杂种优势较强,可以利用杂种优势获得根系活力大、ATP酶活性强的基因型;农大202、农大203和秦烟96的一般配合力均较高,是提高根系吸钾能力的较为理想的亲本;F1杂交组合中农大203×NC628和云烟85×NC628根系生理特性的综合表现较好,可作为选育烤烟钾素高效吸收基因型的材料。
烤烟;根系生理特性;配合力;遗传力;杂种优势
钾在维持烤烟的正常生长、提高抗逆性和改善烟叶品质等方面起着重要的作用,如何提高烟叶钾含量一直是北方烟区烟叶生产中的一个难题[1-2]。由于我国钾肥资源缺乏、成本较高,培育钾高效吸收的品种,提高烤烟营养效率就显得尤为迫切和重要。研究表明,水稻[3]、小麦[4]、玉米[5]钾高效吸收基因型表现出根系发达、根系体积和吸收面积大的特点;钾高效吸收的棉花[6]、番茄[7]、香蕉[8]的根系活力高于吸钾效率低的品种;不同基因型籽粒苋[9]和苎麻[10]吸钾能力强的具有更高的ATP酶活性和阳离子交换量(CEC);杨志晓等[11]研究发现,烤烟富钾基因型根系吸收钾素能力强于一般基因型[11];谢友[12]的研究有同样的发现,吸钾能力强的烤烟品种GK2和GK7具有根系量大、吸收面积大、根系活力强、ATP酶活性高以及根系CEC大等特点。
侯静[13]认为根体积和根系活力可以作为判别棉花钾吸收效率的指标;张宁等[14]发现根体积可以作为水稻苗期钾高效基因型的筛选指标;杨佳丽[7]、庄磊等[15]的研究表明,根系活力可以作为筛选番茄钾高效基因型的指标,低钾处理后21 d为最佳筛选时期;乙引等[16]的研究发现,高粱根中K+-ATP酶依赖K+的刺激或者直接转运K+,K+-ATP酶可能成为判断植物吸收转运K+能力的一个重要指标;崔国贤等[10]发现苎麻不同基因型的根系CEC与吸钾能力关系密切;李廷轩等[9]指出籽粒苋植株的富钾能力不仅与其根系CEC较大有一定关系,而且也与根系的ATP酶活性高度相关。
前人研究表明,根系生理特性与植物对钾的吸收密切相关,可以作为筛选钾高效吸收基因型的重要指标,但对根系生理特性遗传规律的研究报道并不多,在烟草上尚未见过相关报道。本研究选用5个大田烟叶钾含量不同的烤烟品种为材料,利用双列杂交设计研究了烤烟旺长期根体积、根系活力、根系CEC和ATP酶活性的遗传特性,以期为选育高效吸钾型烤烟提供科学依据。
1 材料与方法
1.1 试验材料
选用大田烟叶钾含量不同的农大202、秦烟96、农大203、NC628和云烟85为亲本(其中,农大202、秦烟96和农大203大田烟叶钾含量较高,吸钾能力较强,是高钾基因型,NC628和云烟85为钾含量和吸钾能力一般的基因型[17-18]),按照Griffing方法Ⅱ,组成5×5双列杂交设计[19],亲本及各杂交组合见表1。杂交种于2014年在河南农业大学科教园区制备。试验所用烟草种子全部由河南 农业大学烟草育种实验室提供。

表1 供试的5个亲本和杂交组合Table 1 The selected five parents and hybrid combinations
1.2 试验设计
盆栽试验于2015年在河南农业大学网室中进行,基质为钾长石。当各基因型烟草长至6~7片真叶时,移栽到高20 cm,直径26 cm的塑料盆中,每盆装钾长石约5 kg,盆底用200目尼龙网覆盖2层。每个基因型各种植12株。烟株生长所需营养由Hoagland营养液提供,根据烟株生长需求,还苗期每天浇灌营养液50 mL,伸根期每天浇灌100 mL,旺长期每天浇灌200 mL,常规管理。55 d后结束培养,每个处理取3株,设置3个重复,进行相关指标的测定。
1.3 测定项目与方法
1.3.1 根体积 采用排水法测定[20]。
1.3.2 根系活力 用TTC法测定[21]。
1.3.3 根系ATPase 活性 利用K+-ATPase试剂盒(南京建成生物公司)测定,每小时每毫克根系组织中ATP酶产生1 μmol无机磷的量即为一个ATP酶的活力单位。称取0.2 g鲜根与生理盐水按1∶9研磨、配制成10%的匀浆,4 ℃ 6000×g离心15 min,吸取上清液与生理盐水按1∶4稀释到2%浓度,之后根据试剂盒使用步骤进行测定。
1.3.4 根系阳离子交换量(CEC) 依据崔国贤等[10]的方法,将用蒸馏水冲洗过的根系在80 ℃烘至恒重,粉碎过40目筛。称取0.20 g样品,加入200 mL 0.01 mol/L HCl 处理5 min后过滤,用约200 mL去离子水冲洗滤渣,再用200 mL 1 mol/L KCl溶液(pH 7.0)冲洗滤渣到250 mL的烧杯中,用0.01 mol/L KOH滴定,以7~8滴酸碱混合指示剂(1 g/L中性红的乙醇溶液与1 g/L亚甲基蓝的乙醇溶液等体积混合,pH 7.0时为紫蓝色)指示滴定终点。
1.4 数据处理分析
试验数据采用DPS 7.05中LSD法进行多重比较;用Excel 2010进行作图分析;遗传力和配合力计算采用DPS 7.05中亲本及一组F1材料GRIFIFING配合力分析进行统计分析。
杂种优势计算:
平均优势=(F1-MP)/MP×100%
超高亲优势=(F1-HP)/HP×100%
超低亲优势=(F1-LP)/LP×100%
式中F1为杂种一代性状均值,MP、HP和LP分别为双亲均值、高值亲本和低值亲本。
2 结 果
2.1 亲本及杂交组合根系生理特性差异分析
移栽后55 d,烟株生长处于旺长期,此时烟株代谢旺盛,对矿质元素的需求量大,该时期是反映植株对矿质元素吸收基因型间差异的最佳时期[22]。各基因型4个根系生理性状的平均值及其变异列于表2。从表2可以看出,亲本和F1杂交组合各指标差异显著。其中,ATP酶活性的变异系数最大,为23.33%;CEC的变异系数最小,为8.77%;F1杂交组合各指标变异系数大小顺序为:ATP酶活性>根体积>根系活力>CEC。由此可知,根体积和ATP酶活性的变异范围较大,有利于选育不同根体积和ATP酶活性的烤烟品种,在烤烟育种中可以作为重要的选择目标。
2.2 烤烟根系生理特性配合力方差分析
由表3可以看出,烤烟根体积、根系活力、ATP酶活性和CEC一般配合力(GCA)方差、特殊配合力(SCA)方差分析结果均达到极显著水平,表 明烤烟这4个性状的遗传既有加性效应基因的作用,也有非加性效应基因的作用。

表2 烤烟基因型间根系生理特性差异Table 2 Differences in root physiological characteristics of flue-cured tobacco among genotypes
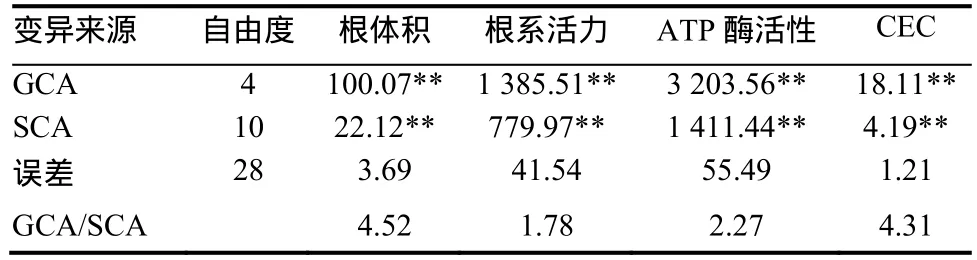
表3 烤烟根系生理特性配合力方差Table 3 Combining ability ANOVA of physiological characteristics of flue-cured tobacco
从GCA/SCA比值并结合表4中加性和显性方差值来看,烟草根体积和CEC的2个性状的GCA/SCA均大于4,且加性方差>显性方差,说明这2个性状的遗传主要以基因的加性效应为主,改良这两个性状在早代选择效果较好。根系活力和ATP酶活性的GCA/SCA分别为1.78和2.27,均小于3但大于1,显性方差>加性方差,表明这2个性状的遗传以基因的显性效应为主,基因的加性效应也占有相当大的比重,可以利用杂种优势来改良这些性状,但要获得稳定的品系需要进行连续定向选择。
2.3 烤烟根系生理特性的遗传参数估计
烤烟根系生理性状的遗传参数列于表4。由表4可知,4个性状的广义遗传率均较高,都在60%以上;各性状广义遗传率hB2的大小顺序为:根系活力>ATP酶活性>根体积>阳离子交换量。根体积和CEC的狭义遗传率hN2值较高,分别为54.81%和46.18%;根系活力最低,小于20%;各性状狭义遗传率hN2的大小顺序为:根体积>阳离子交换量> ATP酶活性>根系活力。由此可知,广义遗传率高的性状,其狭义遗传率不一定高,如根系活力的hB2高达87.97%,但是hN2仅为18.22%;而根体积的 hB2虽低于根系活力,但其hN2大于50%;因此在烤烟杂种后代的选择过程中,根体积和CEC这2个性状可以在早代进行单株选择,而根系活力和ATP酶活性可以进行连续定向选择。
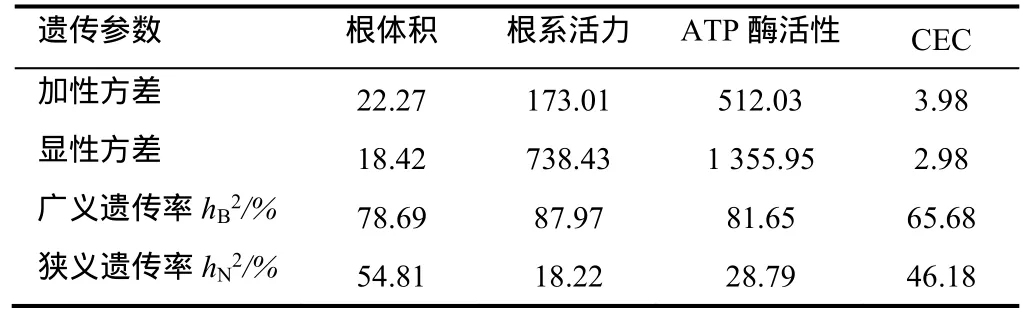
表4 烤烟根系生理特性的遗传参数Table 4 Heritability of physiological characteristics of flue-cured tobacco
2.4 烤烟根系生理特性的一般配合力和特殊配合力分析
亲本及组合的配合力效应值列于表5中。从表5可以看出,除了秦烟96在根体积的遗传上一般配合力为负值,农大202、农大203和秦烟96在4个根系生理性状上的一般配合力均为正值,表现出正向效应;而NC628和云烟85这4个性状的一般配合力均为负值,表现出负向效应。5个亲本一般配合力效应值大小差异显著,农大203根系活力和CEC这2个性状一般配合力最大;农大202根体积一般配合力值最大;而秦烟96 ATP酶活性一般配合力值最大。农大202、农大203和秦烟96在提高根系养分吸收能力上是较为理想的亲本。
从表5可以看出,烤烟同一性状不同杂交组合的特殊配合力效应存在较大的差异。根体积SCA正向效应较大的组合有NC628×农大202、云烟85× NC628和农大203×农大202;根系活力SCA正向效应较大的组合有农大203×NC628、秦烟96×农大202、云烟85×NC628和云烟85×农大203;ATP酶活性SCA正向效应较大的组合有秦烟96×农大202、农大203×NC628和秦烟96×云烟85;根系CEC SCA正向效应较大的组合有农大203×NC628和云烟85×NC628。说明这些组合的亲本表现较好和(或)双亲遗传差异较大,是加性和非加性效应互作的结果。
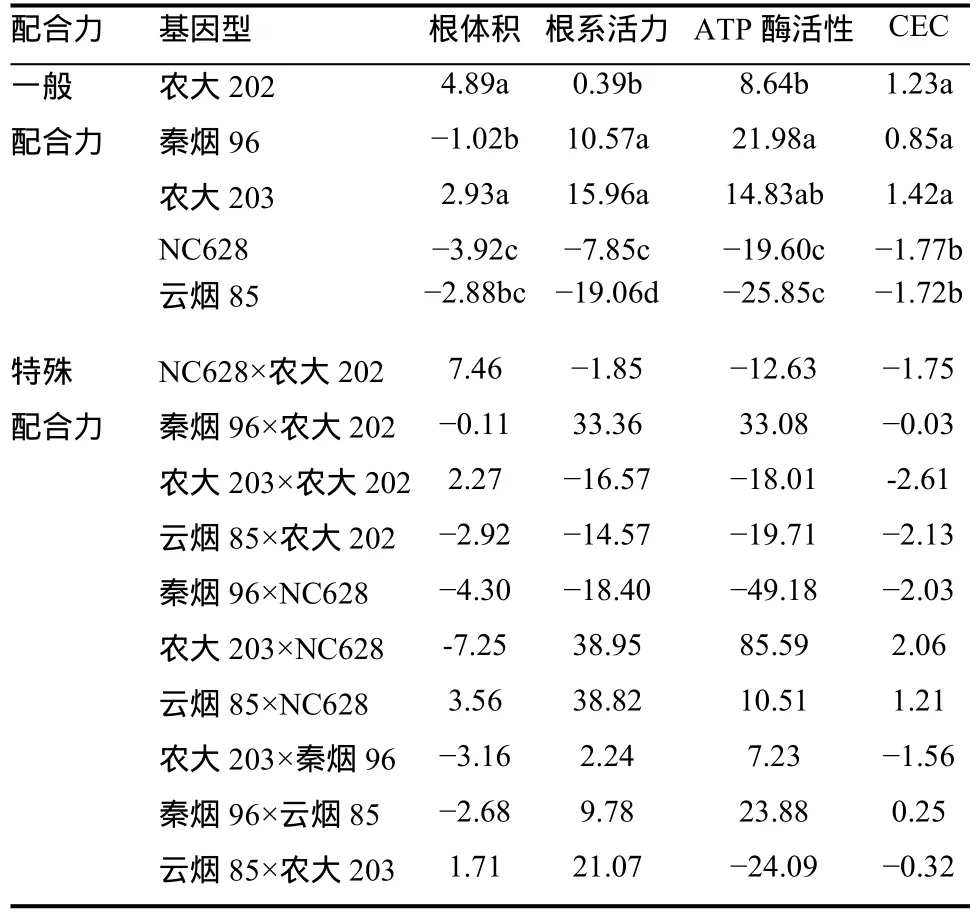
表5 烤烟基因型间根系生理特性的一般配合力 和特殊配合力Table 5 GCA and SCA effects of physiological characteristics of flue-cured tobacco among genotypes
2.5 烤烟根系生理特性的杂种优势分析
2.5.1 烤烟根体积杂种优势分析 F1杂交组合根体积的杂种优势表现列于表6。从表6可以看出,F1根体积表现出中亲优势和超高亲优势的组合有3个,占全部组合的30%;表现出超低亲优势的组合有6个,占全部组合的60%。其中,农大202的杂交后代杂种优势较强,结合表4和表5,农大202可作为优良亲本,配置增加根体积的杂交F1代。

表6 烤烟根体积杂种优势Table 6 Heterosis of root volume of flue-cured tobacco %
2.5.2 烤烟根系活力杂种优势分析 F1杂交组合根系活力的杂种优势表现列于表7。结果表明,F1根系活力有80%的杂交组合表现出中亲优势,50%的组合表现出超高亲优势,90%的组合表现出超低亲优势。这表明F1根系活力杂种优势很强,可以利用杂种优势获得根系活力较高的烤烟基因型。
2.5.3 烤烟根系ATP酶活性杂种优势分析 F1杂交组合的根系ATP酶活性杂种优势表现列于表8。由表8可知,10个F1杂交组合的根系ATP酶活性有50%表现出中亲优势,40%的组合表现出超高亲优势,100%的组合表现出超低亲优势。这表明F1根系ATP酶活性杂种优势较强,杂种优势可以利用。 2.5.4 烤烟根系CEC杂种优势分析 F1杂交组合的根系CEC杂种优势表现列于表9中。结果显示,10个F1组合根系CEC仅2组表现出中亲优势,仅1组表现出超高亲优势。这表明F1根系CEC杂种 优势并不强,在育种上很难通过杂种优势获得烟株根系CEC大的杂交组合。

表7 烤烟根系活力杂种优势分析Table 7 Heterosis of root vigor of flue-cured tobacco %
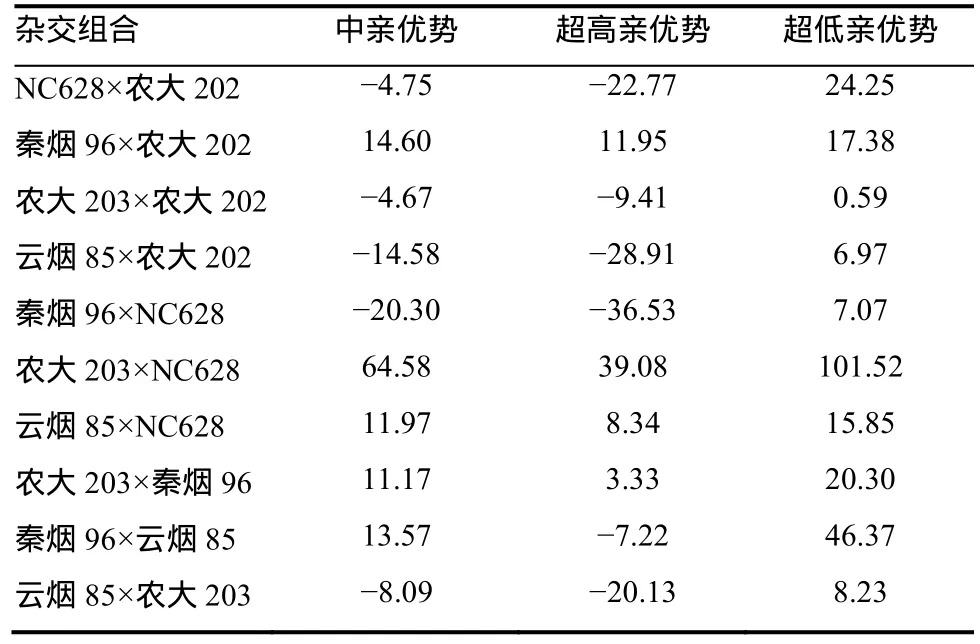
表8 烤烟根系ATP酶活性杂种优势Table 8 Heterosis of root ATPase activity of flue-cured tobacco %
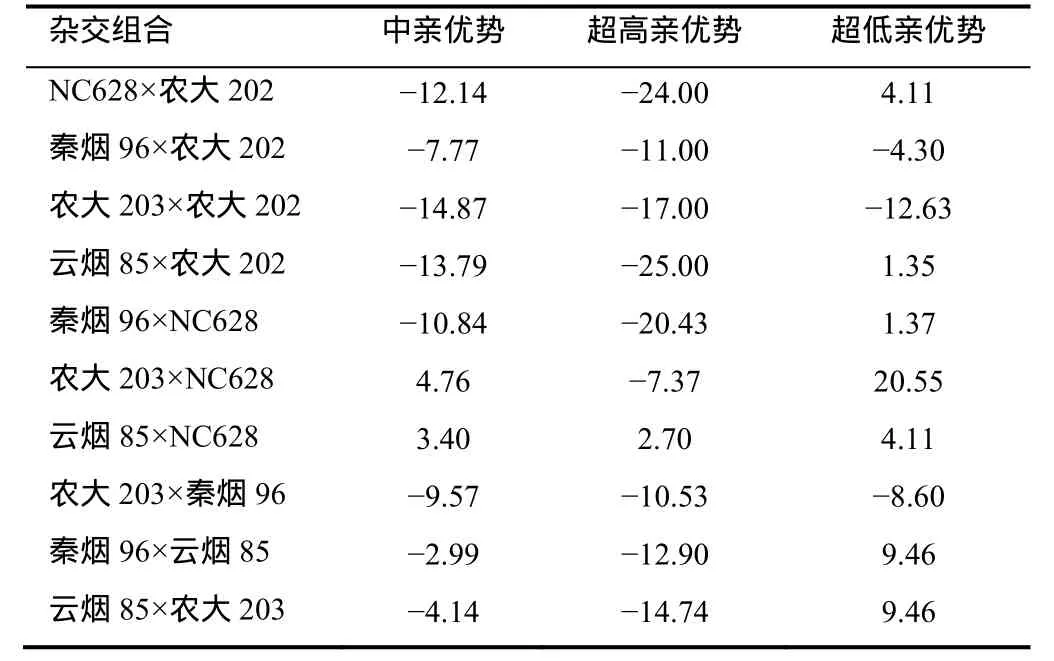
表9 烤烟根系CEC杂种优势Table 9 Heterosis of root CEC of flue-cured tobacco %
3 讨 论
根系是作物吸收和运输养分的主要器官,根系功能的发挥与根系生理特性关系密切[23-24]。根系活力是根系代谢状况的表征,反应了根系吸收矿质营养和水分的能力,根系活力越高,吸收能力就越强,对地上部分水和营养的供给量就越多[25-26];ATP酶是能量代谢的关键酶,它通过水解ATP为根系吸收和转运矿质营养提供能量,对根系吸收养分有直接的影响,其活性越高则根系代谢就越旺盛[27-28];根系CEC与作物吸收矿质元素密切相关,随着根系CEC的增大,作物吸收矿质养分也增多[29]。
王秀全等[30]在研究了玉米大田腊熟期的根系特性遗传表现后指出,根体积、鲜干重,根条数及总长的遗传以加性效应为主;陈加敏等[31]的研究发现,干旱胁迫下大豆苗期根系主根长、根系总长以及根干重的遗传都具有多基因效应,并且都受2对主基因控制;扈光辉等[32]的研究表明,低温逆境条件下非加性效应在不同耐冷玉米种质相对根干重和根总长的遗传中起重要作用,两个指标狭义遗传力均低于50%且相对根干重>相对根总长,在晚代进行选择较为有利。春亮等[33]的研究结果表明,不同基因型玉米苗期根系性状在不同供氮水平下其GCA、SCA效应存在显著的差异,各根系性状在高供氮水平下以非加性效应遗传为主(轴根长除外),低供氮水平下则以加性遗传为主;根系的干重,总、侧根长度在不同供氮水平下的广义遗传率均较高,狭义遗传力在低氮胁迫下表现出上升趋势。本研究结果表明烟草根体积的广义遗传率很高,基因效应在遗传中起重要作用,且主要受基因的加性效应控制,选育根系量较大的品种在早代选择效果较好。
刘红梅等[34]的研究表明,水稻根系活力广义遗传率很高,且主要受非加性效应的影响,根系活力的竞争优势与母本GCA呈现显著的正相关关系,母本一般配合力高的杂交稻组合其根系活力的竞争优势也更强。曹树青等[35]利用4×4(不育系和恢复系)不完全双列杂交设计,研究了籼型水稻根系活力及其衰退值的遗传规律,结果显示,根系活力 和它的衰退值的广义遗传力都大于85%,GCA和SCA方差达到极显著水平,两个指标均表现正向的杂种优势;根系活力遗传以非加性效应为主,而根系活力衰退值的遗传以加性效应为主,两者均可作为有效的育种选择指标。周小平等[36]的研究发现,在玉米经济产量形成过程中,杂交种F1的根系活力有明显的杂种优势,且与光合特性有显著的正相关关系,根系活力可以作为玉米高产杂交种选育的指标。赵全志等[37]在杂交小麦的研究过程中得出了类似的结论。本研究结果表明,烤烟根系活力的遗传与前人研究结果一致,广义遗传率较高,且以基因的显性效应为主,杂种优势较强,可以利用杂种优势获得根系活力较高的烤烟基因型。
研究表明,栽培大麦和野生大麦液泡H+-ATP酶活性在对盐胁迫的响应上差异显著,野生大麦XZ16的耐盐能力明显强于栽培大麦YERONG和GAIRDNER[38]。金蕙芬等[39]在研究了玉米杂交及其亲本自交在授粉后不同时期的ATP酶活性后发现,授粉后1~2 d杂种子房的ATP酶活性显著高于两亲本,授粉后第3天,杂种子房的ATP酶活性呈现出先持续下降,随后又上升的趋势;授粉后15~45 d杂种胚中ATP酶活性均高于两亲本。玉米杂交种苗期根尖细胞ATP酶活性大于双亲或者在双亲之间,杂种幼苗根系发达、吸收水分和矿质营养的能力增强[40]。郭培国等[41]的研究表明,水稻杂交种F1的Ca2+-ATP酶和Mg2+-ATP酶活性和叶中ATP含量都显著高于亲本,两个ATP酶的杂种优势也很明显。本研究结果表明,烤烟根系ATP酶活性遗传以基因的显性效应为主,遗传力较高,杂种优势较强,可以利用杂种优势获得ATP酶活性较高的烤烟基因型,与前人研究结论一致。
DABHOLKAR[42]利用3个地方品种和3个有潜力的品种,采用双列杂交设计研究了高粱阳离子交换量。CEC这一性状由加性和非加性效应控制,CEC具有一定的杂种优势。本研究结果表明,烤烟根系CEC的广义遗传率较高,遗传以基因的加性效应为主,选育根系CEC较大的品种在早代就可以进行单株选择。这与DABHOLKAR研究结果不一致,可能是由于作物种类不同导致的。
由于4个根系性状的特殊配合力基因型方差达极显著水平,所以在钾高效吸收烤烟育种中亲本一般配合力和组合特殊配合力均应考虑。本试验中,除了秦烟96在根体积的遗传上一般配合力为负值,农大202、农大203和秦烟96在4个根系生理性状上的一般配合力均为正值,表现出正向的效应,这3个亲本在钾高效吸收品种选育上具有较大的潜力,通过测交组配有可能选育出钾吸收效率超亲品系材料。
4 结 论
烤烟4个主要根系生理性状的遗传受到加性和非加性效应基因的共同控制,广义遗传率较高。其中,烤烟根体积和根系CEC的遗传以基因的加性效应为主;烤烟根系活力和ATP酶活性的遗传以基因的显性效应为主,F1杂种优势较强,杂种优势可以利用。
[1] 刘国顺. 烟草栽培学[M]. 北京:中国农业出版社,2003:143-145.
[2] HAWKS S N, COLLINS W K. Principles of flue-cured tobacco production[M]. Raleigh: North Carolina State University, 1983.
[3] JIA Y B, YANG X E, FENG Y, et al. Differential response of root morphology to potassium deficient stress among rice genotypes varying in potassium efficiency[J]. Journal of Zhejiang University-Science B(Biomedicine & Biotechnology),2008, 9(5): 427-434.
[4] 张永清,毕润成,庞春华,等. 不同品种春小麦根系对低钾胁迫的生物学响应[J]. 西北植物学报,2006,26(6):1190-1194.
[5] 王朝东,李迎春,王西志,等. 不同基因型玉米适应低钾胁迫的根系响应机理[J]. 河北农业大学学报,2012,35(6):1-7.
[6] 王晓茹,董合林,李永旗,等. 棉花不同品种钾吸收效率差异的根系形态学和生理学机理[J]. 棉花学报,2016(2):152-159.
[7] 杨佳丽,许向阳,李景富. 番茄苗期钾高效基因型筛选方法研究[J]. 北方园艺,2015(12):40-42.
[8] 饶宝蓉,杜中军,罗海燕,等. 不同基因型香蕉对钾高效吸收的根系形态学和生理学特征[J]. 热带作物学报,2010(9):1480-1486.
[9] 李廷轩,张锡洲,王昌全,等. 根系CEC、ATP酶活性和根际微生物对籽粒苋富钾能力的影响[J]. 四川农业大学学报,2002(4):354-356.
[10] 崔国贤,李宗道. 苎麻不同基因型的吸钾能力及其与根系参数的关系[J]. 农业现代化研究,2000(6):371-375.
[11] 杨铁钊,杨志晓,聂红资,等. 富钾基因型烤烟的钾积累及根系生理特性[J]. 作物学报,2009,35(3):535-540.
[12] 谢友. 烤烟钾高效基因型新品系钾素营养特性及其机理研究[D]. 长沙:湖南农业大学,2010.
[13] 侯静. 棉花钾高效基因型筛选及机理的初步研究[D]. 乌鲁木齐:新疆农业大学,2007.
[14] 张宁,郭荣发. 不同钾效率水稻品种苗期的根系形态与生理指标[J]. 江苏农业学报,2014(4):716-720.
[15] 庄磊,许向阳,李景富. 番茄钾高效鉴定指标的研究[J]. 东北农业大学学报,2009(4):25-28.
[16] 乙引,汤章城. 经渗透胁迫高梁根的质膜囊泡K+-ATP酶及其运输特征[J]. 植物生理学报,1996,22(3):231-236.
[17] 何冰. 烟草基因型间钾吸收特性差异及其吸收机理的探讨[D]. 郑州:河南农业大学,2015.
[18] 孙渭,陈志强,马英明,等. 烤烟新品种秦烟96的选育及其特征特性[J]. 中国烟草科学,2012,33(2):28-33.
[19] 王毅,林国平,黄文昌,等. 白肋烟烟碱、总氮含量及氮碱比的配合力与遗传力分析[J]. 中国烟草学报,2007(3):52-56.
[20] 张志良. 植物生理学实验指导[M]. 3版. 北京:高等教育出版社,2003:267-269.
[21] 邹琦. 植物生理学实验指导[M]. 北京:中国农业科技出版社,2000:120-124.
[22] 杨铁钊,彭玉富. 富钾基因型烤烟钾积累特征研究[J].植物营养与肥料学报,2006(5):750-753.
[23] LYNCH J P. Root architecture and plant productivity[J]. Plant physiol, 1995, 109: 7-13
[24] GLASS A D M, SIDDIQI M Y, GILES K L. Correlation between potassium uptake and hydrogen flux in barley varieties: A potential screening method for the isolation of nutrient efficient lines[J]. Plant physiol, 1981, 68: 457-459.
[25] 谭勇,梁宗锁,王渭玲,等. 氮、磷、钾营养对膜荚黄芪幼苗根系活力和游离氨基酸含量的影响[J]. 西北植物学报,2006(3):478-483.
[26] MENGL K. Response ofvarious crop speciesand cultivars to fertilizer application[J]. Plant and soil, 1983, 72: 305-319.
[27] 王空军,董树亭,胡昌浩,等. 我国玉米品种更替过程中根系生理特性的演进Ⅰ.根系活性与ATPase活性的变化[J]. 作物学报,2002(2):185-189.
[28] MARSHNER H, ROMHELD V, CAKMAK I. Mineral Nutrition of Higher Plants[M]. London: Academic Press, 1995: 270-274
[29] 李廷轩,马国瑞. 籽粒苋不同富钾基因型根际钾营养与根系特性研究[J]. 水土保持学报,2004(3):90-93.
[30] 王秀全,陈光明,何丹,等. 玉米根系性状的遗传及相关分析[J]. 中国农业科学,2000,33(增刊):119-123.
[31] 陈加敏. 大豆苗期耐旱性的鉴定及苗期耐旱性和根系性状的遗传研究[D]. 南京:南京农业大学,2004.
[32] 扈光辉,王天宇,苏俊,等. 玉米种质苗期耐冷性状的遗传分析[J]. 中国农学通报,2009(6):101-106.
[33] 春亮,陈范骏,米国华,等. 玉米苗期根系对氮胁迫反应的配合力分析[J]. 植物营养与肥料学报,2005(6):44-50,57.
[34] 刘红梅. 杂交稻米质与生理生化特性的遗传及相互关系研究[D]. 长沙:湖南农业大学,2013.
[35] 曹树青,邓志瑞,翟虎渠,等. 籼型杂交水稻根系活力及其衰退特性的配合力及杂种优势分析[J]. 中国水稻科学,2002(1):20-24.
[36] 周小平,张岁岐,杨晓青,等. 玉米根系活力杂种优势及其与光合特性的关系[J]. 西北农业学报,2008,17(4):84-90.
[37] 赵全志,吕德彬,程西永,等. 杂种小麦群体光合速率及伤流强度优势研究[J]. 中国农业科学,2002,35(8):925-928.
[38] 佐拉(ZAHRA JABEEN). 野生大麦与栽培大麦耐盐性的生理及遗传差异研究[D]. 杭州:浙江大学,2014.
[39] JIN H F. A comparative study on morphology and ATPase activities of maize hybrid and its parents during early embryogeny[J]. Journal of integrative plant biology, 1986(2): 161-168.
[40] 金蕙芬,王国芳,陆春祥. 玉米杂种与其亲本间根系活力及ATP酶RNA酶活性的比较研究[J]. 浙江农村技术师专学报,1992(3):22-28.
[41] 郭培国,李明启. 杂交水稻及其亲本光合特性的研究Ⅱ.功能叶片的希尔反应、光合磷酸化、ATP酶活性和ATP含量[J]. 热带亚热带植物学报,1997(1):65-70.
[42] DABHOLKAR A R, 石春海. 高粱阴阳离子交换量的配合力分析[J]. 国外农学-杂粮作物,1988(4):15-16.
Analysis of Genetic Effects of Physiological Characteristics of Root in Flue-cured Tobacco
XU Jie1, YANG Bingzhao2, MA Wenguang3, PENG Yufu2, SUN Zhihao1, ZHAO Ke1, YANG Tiezhao1*
(1. College of Tobacco Science, Henan Agricultural University, Zhengzhou 450002, China; 2. Henan Tobacco Industrial Corporation, Zhengzhou 450016, China; 3. Yuxi Zhongyan Tobacco Seed Company, Yuxi, Yunnan 653100, China)
Fifteen hybrids were obtained by crossing 5 different genotypes of flue-cured tobacco with different potassium content selected according to the Griffing Ⅱ model. Genetic effects of 4 main physiological characteristics of root were analyzed at the fast-growing stage of flue-cured tobacco under plenty of potash via pot cultivating method. According to the results, there were significant differences between general combining ability and special combining ability of the 4 traits. Their broad-sense heritability was over 60%, indicating that variance in these traits was mainly controlled by genetic factors. The heritability of root volume and CEC of flue-cured tobacco was mainly controlled by additive effects, suggesting that in breeding tobacco cultivars for high-volume and high-CEC, selection at early generations is more efficient. The heritability of root vigor and ATPase activity of flue-cured tobacco was mainly controlled by dominance effects, and the heterosis of F1 generation was also stronger. High-vigor and high-activity of ATPase could be obtained by heterosis utilization. Nongda203×NC628 and Yunyan85×NC628 could be chosen as materials to breed for nutrient efficient flue-cured tobacco varieties (lines) for their good comprehensive performance.
flue-cured tobacco; physiological characteristics of root; combining ability; heritability; heterosis
S572.03
1007-5119(2017)04-0001-08
10.13496/j.issn.1007-5119.2017.04.001
河南中烟工业有限责任公司科技项目“河南浓香型优质烤烟品种筛选与工业应用研究”(HNZY102013002)
许 杰(1989-),在读硕士研究生,主要从事烟草遗传育种与品质改良研究。E-mail:xujie2025@126.com *通信作者,E-mail:yangtiezhao@126.com
2017-02-24
2017-06-19
猜你喜欢
中国糖料(2023年4期)2023-11-01 09:34:46
农业科技通讯(2023年1期)2023-02-12 07:09:28
蔬菜(2021年7期)2021-11-27 03:45:25
中国果业信息(2019年11期)2019-01-05 20:47:24
猪业科学(2018年5期)2018-07-17 05:56:14
河北农业科学(2016年2期)2016-09-09 01:41:31
西南农业学报(2016年5期)2016-05-17 05:42:23
山西农经(2016年5期)2016-02-28 14:24:37
新疆农垦科技(2014年10期)2014-02-28 19:21:08
茶叶通讯(2014年2期)2014-02-27 07:55:39
