
基于氮素效应的烤烟叶面积指数动态模拟*
2017-09-03 08:42邵惠芳刘志宏崔登科黄五星许自成
中国生态农业学报(中英文) 2017年9期
邵惠芳, 刘志宏, 崔登科, 穆 童, 黄五星, 许自成
基于氮素效应的烤烟叶面积指数动态模拟*
邵惠芳1, 刘志宏1, 崔登科2, 穆 童1, 黄五星1, 许自成1
(1. 河南农业大学烟草学院 郑州 450002; 2.天昌国际烟草有限公司宝丰复烤厂 平顶山 467400)
为明确不同施氮水平下烤烟群体叶面积指数的动态特征以及其与活动积温的关系, 本研究以‘豫烟12’、‘秦烟96’、‘云烟87’为供试材料, 设4个施氮水平, 分别为N0(0 kg·hm-2)、N1(30 kg·hm-2)、N2(60 kg·hm-2)、N3(90 kg·hm-2), 测定分析不同积温下烤烟群体叶面积指数及其动态特征, 利用Curve Expert 1.40软件模拟并通过极限值法筛选建立了归一化积温模型, 为烤烟群体光合结构的改善提供一定的理论依据。结果表明: 1)烤烟群体叶面积指数随移栽后活动积温呈偏度<0的单峰曲线变化, 而随着施氮水平的增加呈现增加趋势, 相同施氮水平下烤烟群体叶面积指数峰值大小表现为‘秦烟96’>‘云烟87’>‘豫烟12’。2)有理函数模型=(+)/(1++2)具有较好的模拟效果和生物学意义, 能够很好地模拟烤烟群体相对叶面积指数随相对活动积温的变化, 决定系数为0.980 7**; 利用2015年试验数据对模型进行检验, 模拟准确度()均大于0.958, 精确度(2)均大于0.95, 均方根误差RMSE均小于6.04%。3)模型参数在某些品种和施氮水平之间表现出显著差异性, 品种和施氮量主要通过调节模型参数、、实现对整个模拟模型的调节。4)烤烟群体相对叶面积指数变化速率曲线呈“N”型变化, 反映了烤烟群体叶面积指数的实际变化趋势。5)施氮量对模型次级参数具有调节作用, 随着施氮量增加烤烟群体平均叶面积指数、叶面积指数最大值呈增加趋势, 可作为烤烟群体叶面积指数氮素调节的重要参考指标。该模型的建立可以为烤烟群体发育动态监测以及烤烟群体叶片光合特性的提升提供理论依据和决策支持。
烤烟; 叶面积指数; 活动积温; 施氮量; 有理函数模型
叶片是作物进行光合作用、蒸腾作用及有机物质合成的主要器官[1], 其功能性状可以直接反映植物的遗传特性和对资源的有效利用率[2-3]。光合作用是作物产量和品质形成的基础, 叶面积作为重要的功能性状, 对作物群体的光合作用以及整个生长过程有重要的影响[4-5]。叶面积指数(leaf area index, LAI)是量化群体结构的重要参数[6], 其大小反映了作物利用太阳辐射能量进行光合作用的强弱, 在一定范围内, 作物产量会随着LAI的增加而提升, 而当LAI达到一定限度后, 作物叶片交叉面积过大, 使田间郁闭, 光照强度减弱, 光合作用速率减小, 进而作物产量降低[7]。因此, 模拟LAI的动态特征是获取作物生长发育状况的重要手段[1], 其对定量分析作物整个生长过程至关重要。同时理想的LAI是培育作物高产群体结构的合理基础[8-9]。为明确作物群体LAI的变化规律, 以建立模型方程为手段对作物群体LAI动态特征进行模拟的定量研究成为热点[10-12]。
烤烟(L.)是以收获叶片为目的的经济作物, 烤烟产量的形成过程就是叶面积增长及干物质积累的过程[13-14], 通过定量分析烤烟LAI的动态变化可以对烤烟的生长状况、病虫害监测、田间管理等起到重要作用[15]。烤烟上有关LAI的研究比较少, 且多与冠层光谱相结合进行研究。刘国顺等[16]利用高光谱参数与烟草LAI等参数相结合建立了逐步回归模型, 经检验模拟效果较好。张正杨等[17]研究了不同高光谱模型检测LAI的精度, 认为主成分分析和神经网络的方法可以提高烟草LAI的反演精度, 验证模型确定性系数分别达0.938和0.889。温度是影响作物叶面积增长的主要生态因子之一[18-19], 而目前有关烟草LAI与生态因子相结合的研究鲜见报道。张明达等[13]建立了基于综合光温指标的烟叶干物质生产模型, 为烤烟产量的预测提供了一条科学的途径。烤烟作为一种特殊的作物, 其在成熟之后即被采收, 且为分次采收, 许多研究只考虑烤烟成熟期之前的变化, 并没有考虑进入成熟期开始采收之后的LAI的变化, 具有一定的局限性。因此, 建立烤烟全生育期内温度和LAI的直接动态模型可为掌握烤烟群体发育动态提供理论依据和决策支持。为了进一步研究烤烟LAI的动态变化规律, 同时由于某一品种烤烟在整个生长发育过程中对活动积温的需求相对稳定, 本文以活动积温代替移栽后天数作为衡量尺度, 参考其他作物有关LAI的动态模拟模型, 采用相对归一化的方法[20]以相对活动积温为自变量, 建立相对活动积温与不同施氮水平下烤烟群体相对LAI关系的积温模型, 通过模型分析不同氮素调控水平下烤烟群体LAI的动态特征以及特征参数的变化, 探讨烤烟群体LAI动态特征模拟作为烤烟田间生长状况监测有效手段的可行性。本研究成果对改善烤烟群体光合效能以及烤烟生长过程的动态监测具有重要的意义。
1 材料与方法
1.1 试验地概况
试验于2015年和2016年在河南省洛阳市洛宁县小界乡烟叶基地单元试验田(111°38′N, 34°26′E, 海拔567 m)进行。2015年烟叶大田生长期(5—9月)均温为24.36 ℃, 降雨量为437.15 mm, 日照时数为835.2 h, 2016年烟叶大田生长期(5—9月)均温为23.59 ℃, 降雨量为415.56 mm, 日照时数为867.3 h。试验地土壤为黄壤土, 前茬作物为小麦(L.), 其地势平坦, 光照充足, 2015年和2016年的土壤基本理化性质见表1。
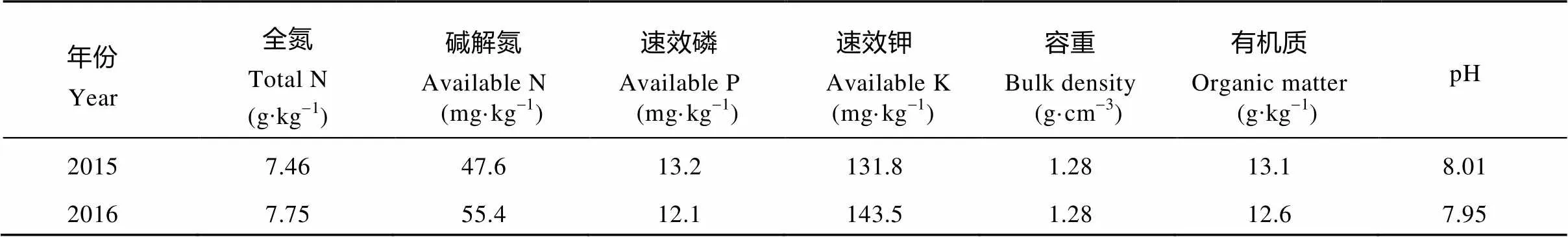
表1 试验地土壤理化性质
1.2 试验处理
2015年和2016年的试验均以施氮量和品种为因素, 裂区设计。主区为施氮量, 施氮量分别设N0(0 kg·hm-2)、N1(30 kg·hm-2)、N2(60 kg·hm-2)、N3(90 kg·hm-2)4个水平; 副区为品种处理, 分别为‘豫烟12’(YU12)、‘秦烟96’(QIN96)、‘云烟87’(YUN87); 共设12个处理小区, 小区面积为120 m2, 每个小区种植烟株200株, 行株距为1.20 m×0.55 m。施氮量处理的70%氮肥在起垄时作为基肥于垄底条施, 剩余30%于团棵期追施。所用肥料为烟草专用复合肥、芝麻饼肥、硫酸钾、硝酸钾、过磷酸钙, N∶P2O5∶K2O=1∶1.5∶3。2015、2016年烤烟移栽期分别为5月13日、5月4日。打顶、去除底脚叶以及成熟烟叶分次采收等是烤烟生产必要的农艺措施, 同时也是烤烟叶面积指数误差产生的重要来源, 且无法避免, 进而需要尽量将这些操作误差降到最低, 以提高分析结果的准确性。因此, 在打顶和去除底脚叶时做到标准一致, 同时烟叶成熟后及时采收, 且只采收成熟的烟叶。除此之外, 各小区其他农事操作保持一致且均按照当地优质烟叶生产技术规范进行。
1.3 测定项目与方法
1.3.1 烤烟群体LAI测定
在烤烟进入团棵期时, 每小区选取生长均匀一致的烟株10株挂牌标记, 并且从挂牌之日起开始对烟株进行测量, 每个处理每次取3株测量所有展开叶(所有着生于烟株上的未成熟的展开烟叶)的长和宽。前期烟叶生长较快, 每隔3~5 d测量一次, 后期每隔8~10 d测量一次。打顶及去除底脚叶等农艺措施进行之前加测。
利用公式(1)计算烤烟LAI:

式中:为烤烟叶面积的校正系数0.634 5[21],为烤烟的种植密度(本试验烤烟种植密度为1.65株·m-2),为测量株数,LW分别代表第株烤烟的第片叶的最大长度和最大宽度。
1.3.2 气象数据测定
采用试验基地的自动气象站记录烤烟大田生育期逐日的气象资料, 气象与试验田直线距离为600 m, 烤烟移栽后的活动积温(TEM)参考严定春等[22]的方法计算。
1.4 数据处理
1.4.1 数据归一化
采用Microsoft Excel 2016对2015年和2016年的LAI、TEM数据进行整理并采用归一化方法进行处理, 得到相对叶面积指数(RLAI)、相对活动积温(RTEM), 计算公式为:

(3)
式中: RLAI、RTEM分别为烤烟不同生育期的相对叶面积指数、相对活动积温, LAI、TEM分别为烤烟不同生育期的叶面积指数、活动积温, LAImax、TEMmax分别为烤烟移栽期到成熟期内的叶面积指数最大值、活动积温最大值
1.4.2 模型建立及检验
利用Curve Expert 1.40软件对2016年的RLAI和RTEM数据进行动态模拟, 得到RLAI与RTEM的动态模拟方程, 并且利用极限值法[20,23]进行筛选, 筛选出具有生物学意义的RLAI动态模型。
利用2015年的数据检验所建立的RLAI模型的精确度和准确度, 由于烤烟生长对积温的需求相对稳定, 将任意时刻的RTEM值代入方程即可得到对应的RLAI模拟值, 同时将实测的LAI值进行归一化处理, 得到RLAI实测值, 用RLAI模拟值和RLAI实测值进行模型检验, 横坐标表示RLAI模拟值, 纵坐标表示RLAI实测值, 两者比较建立的线性方程用=表示, 方程系数与1的接近程度反映了RLAI动态模型模拟结果的准确性, 方程决定系数2的大小反映了RLAI动态模型模拟结果的精确度[20]。
利用SPSS 21.0进行单因素方差分析, 选用最小差异显著法(LSD)进行多重比较, 利用Microsoft Excel 2016作图和制表。
2 结果与分析
2.1 烤烟群体叶面积指数(LAI)动态变化特征分析
由LAI随TEM的变化曲线可以看出(图1), 不同施氮水平下3个烤烟品种的LAI随TEM的变化趋势基本一致, 均表现为LAI先随着移栽后TEM的累积增大, 在移栽后TEM达到1 200 ℃左右时出现峰值, 之后缓慢下降, 在整个生育期内呈偏度<0的单峰曲线变化。与‘QIN96’相比, ‘YU12’和‘YUN87’的波峰较为平缓, 相同施氮水平下, LAI峰值大小表现为‘QIN96’>‘YU12’>‘YUN87’; 在TEM较低时, 同一品种不同施氮量的群体间LAI差值较小, 但随着TEM的累积群体间LAI差值有所增大。同时, 曲线在峰值之后下降部分趋势的陡峭程度反映成熟采收烟叶的多少, 进而反映其成熟速率, 可以看出, 不同品种间的成熟速率为‘QIN96’>‘YUN87’> ‘YU12’。
N0、N1、N2和N3指0 kg·hm-2、30 kg·hm-2、60 kg·hm-2和90 kg·hm-24个施氮水平。N0, N1, N2 and N3 represent the N application of 0 kg·hm-2, 30 kg·hm-2, 60 kg·hm-2and 90 kg·hm-2, respectively.
2.2 烤烟群体相对叶面积指数(RLAI)动态优化模型的建立
2.2.1 烤烟群体相对叶面积指数(RLAI)动态变化
将烤烟LAI和移栽后TEM分别进行归一化处理, 得到各处理归一化后的RLAI与RTEM, 其能够更好地模拟烤烟群体LAI的动态变化, 同时消除了量纲的差异以方便比较。不同施氮水平下RLAI随RTEM的变化趋势与实测烤烟群体LAI变化趋势一致, 均为偏度<0的单峰曲线(图2)。对于不同品种而言, ‘YU12’波峰最为平缓, 其次为‘YUN87’和‘QIN96’。波峰平缓的烤烟群体LAI保持在较高水平的时间较长, 该段时间烤烟的光合生产潜能达到较高水平, 但该段时间其成熟速率较慢。对于不同施氮水平而言, 在相对活动积温较小时, 施氮水平间的差异不太明显; 相对活动积温较大时, 施氮量间差异开始显现, 且以N0施氮水平的相对叶面积指数降低最为明显。
2.2.2 烤烟群体相对叶面积指数(RLAI)动态模拟
选取Curve Expert 1.40软件建立了模拟度较好的8个模型(表2), 并利用极限值法对其进行筛选。当→∞时,值应该趋近于0,从表2可以看出, 模型2、3、6、7、8的值在→∞时却趋近于无穷大或常数, 这并不能够很好地解释RLAI随RTEM的动态变化过程, 没有实际意义; 而模型4、5在当=0时,值不存在, 也不能够做出合理解释, 因此将2~8模型舍弃, 选用模型1对RLAI随RTEM的动态变化进行模拟。
模型1为有理函数模型, 其通式为=(+)/(1++2), 当=0时,==0.020 1, 即为烤烟移栽时的RLAI值,→∞时,=0, 烤烟烟叶完全成熟采收后LAI也为0, 且该模型只有1个峰值, 对应烤烟最大RLAI, 因此, 有理函数模型能够很好地模拟烤烟RLAI随RTEM的变化, 模拟方程的系数分别为=0.020 1,=0.278 5,=-3.312 2,=3.301 9, 标准差为0.042 5, 决定系数2=0.980 7, 达到极显著水平, 其对应的模拟曲线见图3。
2.2.3 烤烟群体相对叶面积指数(RLAI)动态模型关键参数分析
利用2016年不同处理的RLAI和RTEM数据, 分别建立归一化动态模拟方程(表3), 其决定系数均达0.922 0以上, 说明RLAI动态模型能够对不同烤烟品种和施氮量下LAI进行模拟, 对各模拟方程的、、、参数差异的显著性进行分析, 结果表明, 品种和施氮量对各个参数有不同程度的影响。对不同品种而言, 参数在不同品种间无显著差异, 参数在‘YUN87’和‘YU12’之间无显著差异, 参数、在‘QIN96’和‘YUN87’之间无差异; 除此之外, 各参数在品种间差异显著, 且达到0.05显著水平。对于不同施氮水平而言, 参数在不同施氮水平之间差异不显著, 参数、在N0、N1、N2之间差异不显著, 参数在N0与N1以及 N1与N2之间无差异; 除此之外, 各参数在不同施氮量间差异显著, 且达到0.05显著水平。值得注意的是, 参数虽然在不同品种及施氮水平间存在差异, 但是其所代表的是移栽时烤烟群体RLAI值, 且均接近于0, 因此, 参数对模拟方程的影响较小, 品种和施氮量主要通过调节参数、、对动态模拟方程进行调控。
N0、N1、N2和N3指0 kg·hm-2、30 kg·hm-2、60 kg·hm-2和90 kg·hm-24个施氮水平。N0, N1, N2 and N3 represent the N application of 0 kg·hm-2, 30 kg·hm-2, 60 kg·hm-2and 90 kg·hm-2, respectively.

表2 烤烟归一化相对叶面积指数动态共性模型
模型中为相对积温,为相对叶面积指数。**表示达0.01显著水平。andin the model denote relative accumulated temperature and relative leaf area index. ** means significance at 0.01 probability level.
N0、N1、N2和N3指0 kg·hm-2、30 kg·hm-2、60 kg·hm-2和90 kg·hm-24个施氮水平。N0, N1, N2 and N3 represent the N application of 0 kg·hm-2, 30 kg·hm-2, 60 kg·hm-2and 90 kg·hm-2, respectively.

表3 不同施氮水平下不同烤烟品种相对叶面积指数动态优化模型[y=(a+bx)/(1+cx+dx2)]参数
N0、N1、N2和N3指0 kg·hm-2、30 kg·hm-2、60 kg·hm-2和90 kg·hm-24个施氮水平。模型中为相对活动积温,为相对叶面积指数; 括号外小写字母和括号内小写字母不同分别表示烤烟品种间和氮素水平间在<0.05水平差异显著; **表示差异达0.01显著水平。N0, N1, N2 and N3 represent the N application of 0 kg·hm-2, 30 kg·hm-2, 60 kg·hm-2and 90 kg·hm-2, respectively.andin the model denote relative accumulated temperature and relative leaf area index. Different letters outside and inside brackets indicate significant differences (< 0.05) among tobacco varieties and N application levels, respectively. ** means significant difference at 0.01 probability level.
2.2.4 烤烟群体相对叶面积系数(RLAI)动态模型变化速率分析
对烤烟群体RLAI动态模拟方程=(+)/ (1++2)求一阶导数可以得到烤烟群体RLAI变化速率模拟方程¢=(--2-2)/(1++2)2, 将RTEM代入方程得到不同烤烟群体RLAI变化速率的模拟曲线(图4)。曲线结果表明, 不同烤烟群体RLAI变化速率均呈“N”型变化, 其以“0”为分界线, 将曲线分成两部分, 其中大于0的部分表示烤烟RLAI呈增加趋势, 且增加速率先增大后降低, 烤烟群体RLAI增加速率最大时的RTEM为0.4左右, 此时烤烟正处于旺长期; 当烤烟群体RLAI变化速率等于0时, RTEM为0.5左右, 此时烤烟正进入成熟期; 小于0的部分则表示烤烟RLAI呈降低趋势, 且降低速率先增大后减小。
施氮水平为N0时RLAI在‘QIN96’和‘YU12’中的增加和衰减速率的最大值高于其他施氮水平, 且在不同施氮水平之间差异不明显; 而在‘YUN87’中N1水平的RLAI的增加和衰减速率的最大值均高于其他施氮水平, 且在不同施氮水平之间较为发散。
2.3 烤烟群体相对叶面积指数(RLAI)动态优化模型的检验
采用2015年的试验数据对建立的烤烟群体RLAI动态优化模型进行检验。模型检验结果见图5。由检验结果可以看出, 3个品种烤烟群体RLAI模拟值和实测值比较接近, 模拟准确度分别为0.981、0.988、0.975、0.958, 均接近于1, 能够较好地反映模型的准确性; 模拟的精确度2分别为0.978 5、0.974 8、0.960 7、0.967 2, 均达0.95以上, 说明线性方程对实测值的解释率较高; 均方根误差RMSE分别为4.52%、4.71%、6.04%、5.91%, 均比较小, 说明模拟值和实测值比较接近, 模型能够较好地对数据进行还原。总体而言, 模型的精确度和准确度均比较高, 能够较好地反映烤烟群体LAI的动态变化, 满足对烤烟群体LAI预测的需求。
2.4 基本模型的施氮量对烤烟群体最大叶面积指数(LAImax)和平均叶面积指数(MLAI)的影响分析
烤烟群体RLAI动态优化模型能够较好地反映烤烟群体LAI的动态变化特征, 能够很好地对烤烟大田生产进行监测, 对模型数据进行充分挖掘, 例如烤烟群体最大叶面积指数(LAImax)和平均叶面积指数(MLAI)等参数的大小可以间接反映烤烟光合能力, 同时辅助其他一些参数可以更深层次地对烤烟生长动态进行分析, 例如平均净同化率(NAR)、平均作物生长率(CGR)等参数能够从生理角度对烤烟光合性能进行综合分析。由于缺少相应的参数, 本文只对LAImax和MLAI进行分析。
2.4.1 烤烟群体最大叶面积指数(LAImax)与施氮量
群体最大叶面积指数(LAImax)可以指示作物群体的最大同化能力[24], 其在动态模型曲线上与峰值相对应, 施氮水平对其有较大的影响。由图6可知, 在施氮量为0~90 kg·hm-2的范围内, 烤烟群体LAImax与施氮量呈显著线性正相关关系, 施氮量大的处理LAImax相对应较高。‘QIN96’、‘YU12’及‘YUN87’的LAImax与施氮量的回归方程分别为=0.013 4+3.389 5 (2=0.972 4*)、=0.011 4+3.405 3 (2=0.979 8*)和=0.009 5+2.884 7 (2=0.956 3*)。不同品种间LAImax存在差异, ‘YUN87’的LAImax明显小于‘QIN96’和‘YU12’, 而‘QIN96’与‘YU12’的LAImax之间差异并不显著。
2.4.2 烤烟群体平均叶面积指数(MLAI)与施氮量
烤烟群体MLAI在一定程度上可以反映烤烟整个生育期内的物质积累状况, 对烤烟RLAI进行积分可以计算得到平均相对叶面积指数(MRLAI), MRLAI与LAImax的乘积即为MLAI[8], 其可以通过公式(4)、(5)计算得到:
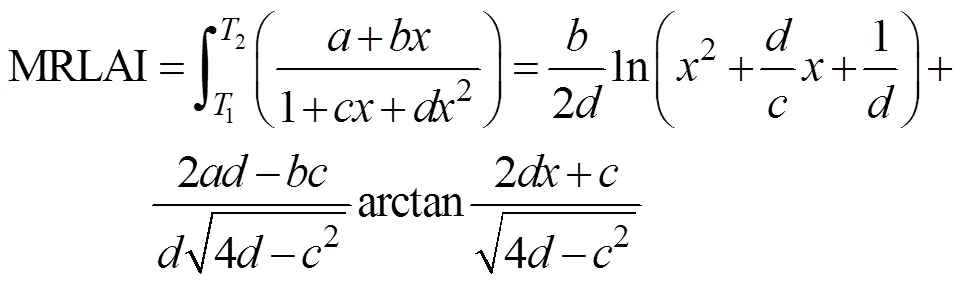
(5)
式中:1、2为相对活动积温, 当1=0,2=1时, 即可求得整个生育期内的平均相对叶面积指数。
由图6可知, 在整个烤烟生育期内MLAI随施氮量增加呈显著线性增加趋势, ‘YU12’、‘YUN87’和‘QIN96’的回归方程分别为=0.007 3+1.757 1 (2=0.993**)、=0.005 8+1.444 6(2=0.994*)和= 0.005 2+1.506 7 (2=0.999*), 说明施氮量的增加会显著影响烤烟生育期内MLAI的增加, 有利于提高烤烟群体的生产潜力; 同时对于不同品种而言, ‘YU12’的MLAI与‘QIN96’和‘YUN87’之间存在显著差异, 而‘QIN96’与‘YUN87’之间差异并不显著。
3 讨论
烤烟作为一种喜温作物, 其对温度反映非常敏感[25]。研究表明,高温和低温都会对植物的正常生长和光合作用产生一定的影响[26-28]。由于烤烟对积温的需求相对稳定, 因此本试验研究了LAI与积温的关系, 随移栽后天数的增加, 有效活动积温增加, 同时烤烟群体LAI随活动积温的累积表现为先增大后减小的变化趋势, 并且呈单峰曲线变化, 这与在玉米(L.)LAI与活动积温研究以及棉花(L.)LAI与辐热积的研究中结果一致[1,9]。
建立作物生长动态变化的数学模型, 应用现代信息技术进行定量化分析和模拟研究, 是获取作物生长发育状况的重要手段[1]。利用数学模型, 能够对作物LAI进行预测, 科学监测作物的生长发育, 及时掌握大面积作物生长状况信息, 为作物估产和种植栽培提供科学决策支持。聂志刚等[29]采用回归正交组合设计, 运用APSIM模型模拟了光照和温度对小麦生长发育的影响, 模型检验精度较高, 对旱地小麦叶片发生规律起到了重要作用。吕新[30]利用积温等生态因素变量代替了以时间变量作为衡量玉米生长发育过程的时间标尺, 比用天数更具有代表性, 能更好地反映玉米的生长状况。因此, 研究生态因子与LAI之间的关系并建立动态模型意义重大。本文从施氮量调控以及积温生态因子调控两方面研究了烤烟LAI动态变化, 并且建立了动态模型, 认为不同施氮水平下烤烟RLAI随RTEM动态变化符合有理数函数=(+)/(1++2)变化, 其能够很好地模拟及解释烤烟RLAI的动态变化, 同时该模型参数少、计算简便, 只需要知道某一时刻的TEM即可反演出任意时刻的LAI, 能够及时掌握LAI的动态变化, 为优质烟叶生产提供理论依据。
氮是烟株生长所有必需营养元素中限制烟草生长和品质的首要因素[31], 其供应量直接决定着烟株的营养状况和烟叶的产量及品质[32]。研究表明, 烤烟单叶质量在一定程度上会随着施氮水平的增加而增加[33]。而单叶质量在很大程度上取决于其叶面积的大小, 因此, 施氮量对烤烟群体叶面积也会产生相应的影响, 施氮量适宜烟株生长发育协调, 烤烟净光合速率会提高, 有利于烟叶良好品质的形成[31]; 施氮量过大, 烟株叶色浓绿, 贪青晚熟[34], 会使得烤烟群体LAImax出现的时间延迟, 其成熟的速率也会有所降低; 施氮量不足, 烟株生长缓慢, 烤烟LAImax与正常水平相比较低。本研究中从图1和图6可以看出LAImax会随着施氮量的增加而增大, 同时其成熟速率有所降低, ‘YUN87’表现最为明显, 这与前人研究结果相一致。因此, 适当使用氮肥, 可以改善叶片光合性能, 为后期烟叶化学成分的转化提供物质基础。
烤烟RLAI动态模型主要通过参数调控实现对烤烟LAI变化的动态模拟, 参数接近于零, 品种和施氮量对其的调控作用较小, 品种和施氮量主要对参数、、调控实现对烤烟LAI的动态模拟。利用烤烟RLAI动态模型还可以计算出LAImax、MLAI等次级特征参数, 实现对烤烟生长状况的综合评价。MLAI是衡量作物整个生育期内物质生产状况的次级特征参数, 随着MLAI的提高, 平均作物生长率也随之提高[35]。本研究认为烤烟群体MLAI随施氮量的增加而增加, 且增加效应在后期表现明显。同时, 烤烟MLAI、LAImax随着施氮量的增加呈现增加趋势, 不同品种间存在一定的差异, 表现为‘YU12’在MLAI上显著高于‘YUN87’、‘QIN96’, ‘YUN87’在LAImax上显著低于‘QIN96’、‘YU12’, 引起这种差异的原因可能是品种特性或者是不同品种对氮素的利用效率不同。因此, 确保适宜的施氮水平才能保证烤烟群体物质生产水平和烤烟最大生产潜力之间达到平衡。同时‘YUN87’能达到的LAImax明显小于‘QIN96’和‘YUN87’, 这可能是与‘YUN87’自身的品种特性以及其在当地的适应性有关。而有关LAImax出现时间的问题, 本文中没有具体体现, 因为各处理LAI的测定时间是一致的, 同时各处理LAImax出现的时间会有所不同, 不可能实现正好在LAImax最大时进行测量, 所以文章中所测量的LAImax是近似值。因此, 可以在旺长期增大测量密度, 增加测量次数, 对测定值进行曲线拟合, 所对应的曲线最大值即为LAImax, 同时LAImax出现的时间也可以确定。
作物群体LAI的变化速率可以准确反映作物在不同生育期的生长快慢。孙锐等[24]在春玉米上研究发现, 不同密度群体LAI变化速率呈“N”型单峰曲线变化; 李向岭等[9]研究发现不同播期玉米LAI变化速率呈“N”型变化趋势。本研究表明, 烤烟群体RLAI的变化速率随积温累积呈“N”型变化, 速率曲线是由烤烟群体RLAI动态模型求导而来, 有其实际的生产意义, 曲线最大值代表RLAI增大速率最大值, 其对应活动积温为900 ℃左右, 此时处于烤烟移栽后65 d左右, 正处于烤烟旺长期, 此时烤烟群体RLAI变化速率应该达到最大; 曲线值为0时, 活动积温达到1 100 ℃, 此时处于烤烟移栽后80 d左右, 烤烟进入成熟期, 此时的RLAI变化速率变为0, 并且开始向负值转化。不同施氮量对‘YU12’、‘QIN96’影响较小, 对‘YUN87’影响较大, 不同施氮量间速率曲线较为分散, 这可能与不同品种对氮素的响应程度有关。利用速率曲线能够及时掌握烤烟群体RLAI变化速率及LAI的动态变化。
本研究只探索了不同施氮水平下烤烟群体RLAI和RTEM之间的关系, 并对该关系进行了初步验证, 烤烟叶片生长还会受到水分、肥料、种植密度等栽培措施的影响, 以及温度、辐射、光照等生态因子的制约, 在这种综合因子调控下的烤烟生长模型还需要进一步的研究和探讨, 而本研究将烟叶的成熟采收纳入到研究对象的范围内, 这是之前的研究所没有涉及的。
4 结论
烤烟群体LAI随TEM呈单峰曲线变化, LAI随施氮量增加而增加, 以‘QIN96’的峰值最大, 同时以‘QIN96’的成熟速率最快。而RLAI随RTEM也呈单峰曲线变化, 以‘YU12’波峰最为平缓, 成熟采收后, 以N0水平的下降趋势最快。采用RTEM为自变量, 利用归一化方法建立RLAI和RTEM的动态模拟模型, 并利用试验数据进行检验, 结果表明,=(0.020 1+0.278 5)/(1-3.312 2+3.301 92)能够很好地预测烤烟叶面积的变化动态, 其决定系数为0.980 7, 模拟准确度()均大于0.958, 精确度(2)均大于0.95, 均方根误差RMSE均小于6.04%。品种和施氮量对模型参数有不同程度的影响, 主要通过对模型参数、、的影响实现对整个模型的调节, 同时模型的次级特征参数能够较好地对烤烟生长状况进行综合评价, 烤烟群体MLAI、LAImax、RLAI变化速率等参数均对氮素会产生一定程度的响应, MLAI、LAImax均随着施氮量的增加而增大, 其中以‘QIN96’的LAImax最大, 以‘YU12’的MLAI最大。而在整个生育期内RLAI变化速率曲线呈“N”变化, 其在不同品种和施氮量间也表现出不同程度差异。因此, 氮施用量可以作为改善烤烟群体叶片光合结构以及协调烤烟产量与质量的重要指标。
[1] 贾彪, 钱瑾, 马富裕. 氮素对膜下滴灌棉花叶面积指数的影响[J]. 农业机械学报, 2015, 46(2): 79–87 Jia B, Qian J, Ma F Y. Simulating effects of nitrogen on leaf area index of cotton under mulched drip irrigation[J]. Transactions of the Chinese Society for Agricultural Machinery, 2015, 46(2): 79–87
[2] 王玉平, 陶建平, 刘晋仙, 等. 不同光环境下6种常绿阔叶林树种苗期的叶片功能性状[J]. 林业科学, 2012, 48(11): 23–29 Wang Y P, Tao J P, Liu J X, et al. Response of leaf functional traits to different light regimes in an evergreen broad-leaved forest in the Jinyun mountain[J]. Scientia Silvae Sinicae, 2012, 48(11): 23–29
[3] 宋彦涛, 周道玮, 王平, 等. 松嫩草地66种草本植物叶片性状特征[J]. 生态学报, 2013, 33(1): 79–88 Song Y T, Zhou D W, Wang P, et al. Leaf traits of 66 herbaceous species in Songnen grassland in Northeast China[J]. Acta Ecologica Sinica, 2013, 33(1): 79–88
[4] 王峰, 陈玉真, 王秀萍, 等. 茶树不同叶位叶片功能性状与光合特性研究[J]. 茶叶科学, 2016, 36(1): 77–84 Wang F, Chen Y Z, Wang X P, et al. Leaf functional and photosynthetic characteristics in different leaves positions of tea plant[J]. Journal of Tea Science, 2016, 36(1): 77–84
[5] Royer D L, Kooyman R M, Little S A, et al. Ecology of leaf teeth: A multi-site analysis from an Australian subtropical rainforest[J]. American Journal of Botany, 2009, 96(4): 738–750
[6] 陆艳婷, 张小明, 黄福灯, 等. 氮素处理对水稻浙粳22产量形成的影响及其生育后期与叶片光合特性间的关系[J]. 核农学报, 2013, 27(3): 373–378 Lu Y T, Zhang X M, Huang F D, et al. Influence of nitrogen treatment on yield and its relation to leaf photosynthesis characteristics of japonica rice variety Zhejing22 at late growing stage[J]. Journal of Nuclear Agricultural Sciences, 2013, 27(3): 373–378
[7] 高鑫. 春玉米LAI和叶片氮素营养及产量的高光谱估测模型研究[D]. 呼和浩特: 内蒙古农业大学, 2016: 45–60 Gao X. The research on the hyperspectral monitoring model of LAI, leaf nitrogen and yield in spring maize[D]. Huhhot: Inner Mongolia Agricultural University, 2016: 45–60
[8] 吴立峰, 张富仓, 王海东, 等. 新疆棉花亏缺灌溉叶面积指数模拟研究[J]. 农业机械学报, 2015, 46(1): 249–258 Wu L F, Zhang F C, Wang H D, et al. Simulation of cotton leaf area index under deficit irrigation in Xinjiang[J]. Transactions of the Chinese Society for Agricultural Machinery, 2015, 46(1): 249–258
[9] 李向岭, 赵明, 李从锋, 等. 玉米叶面积系数动态特征及其积温模型的建立[J]. 作物学报, 2011, 37(2): 321–330 Li X L, Zhao M, Li C F, et al. Dynamic characteristics of leaf area index in maize and its model establishment based on accumulated temperature[J]. Acta Agronomica Sinica, 2011, 37(2): 321–330
[10] Marcelis L F M, Gijzen H. A model for prediction of yield and quality of cucumber fruits[J]. Acta Horticulturae, 1998, 476: 237–242
[11] 汪涛, 黄文江, 董斌, 等. 基于夏玉米冠层内辐射分布的不同层叶面积指数模拟[J]. 农业工程学报, 2015, 31(z1): 221–229 Wang T, Huang W J, Dong B, et al. Leaf area index simulation of different layers in summer maize canopy based on photosynthetically active radiation vertical distribution model[J]. Transactions of the CSAE, 2015, 31(z1): 221–229
[12] 张文宇, 汤亮, 姚鑫锋, 等. 基于过程的小麦株型指标动态模拟[J]. 中国农业科学, 2012, 45(12): 2364–2374 Zhang W Y, Tang L, Yao X F, et al. Process-based simulation model for growth dynamics of plant type index in wheat[J]. Scientia Agricultura Sinica, 2012, 45(12): 2364–2374
[13] 张明达, 李蒙, 胡雪琼, 等. 基于辐热积法模拟烤烟叶面积与烟叶干物质产量[J]. 生态学报, 2013, 33(22): 7108–7115 Zhang M D, Li M, Hu X Q, et al. Simulation of leaf area and dry matter production of tobacco leaves based on product of thermal effectiveness and photosynthetically active radiation[J]. Acta Ecologica Sinica, 2013, 33(22): 7108–7115
[14] 苏永士, 苏富强, 王海波, 等. 不同施肥条件下烟草冠层光谱特征与叶绿素、叶面积指数的关系[J]. 中国烟草科学, 2013, 34(2): 23–27 Su Y S, Su F Q, Wang H B, et al. Relationship of spectral characteristics with chlorophyll and leaf area index in flue-cured tobacco leaf[J]. Chinese Tobacco Science, 2013, 34(2): 23–27
[15] Haboudane D, Miller J R, Pattey E, et al. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture[J]. Remote Sensing of Environment, 2004, 90(3): 337–352
[16] 刘国顺, 李向阳, 刘大双, 等. 利用冠层光谱估测烟草叶面积指数和地上生物量[J]. 生态学报, 2007, 27(5): 1763–1771 Liu G S, Li X Y, Liu D S, et al. Estimation of tobacco leaf area index and aboveground biomass using canopy spectra[J]. Acta Ecologica Sinica, 2007, 27(5): 1763–1771
[17] 张正杨, 马新明, 贾方方, 等. 烟草叶面积指数的高光谱估算模型[J]. 生态学报, 2012, 32(1): 168–175 Zhang Z Y, Ma X M, Jia F F, et al. Hyperspectral estimating models of tobacco leaf area index[J]. Acta Ecologica Sinica, 2012, 32(1): 168–175
[18] 刁明, 戴剑锋, 罗卫红, 等. 温室甜椒叶面积指数形成模拟模型[J]. 应用生态学报, 2008, 19(10): 2277–2283 Diao M, Dai J F, Luo W H, et al. Simulation model on the formation of greenhouse sweet pepper leaf area index[J]. Chinese Journal of Applied Ecology, 2008, 19(10): 2277–2283
[19] Marcelis L F M, Heuvelink E, Goudriaan J. Modeling biomass production and yield of horticultural crops: A review[J]. Scientia Horticulturae, 1998, 74(1/2): 83–111
[20] 张宾, 赵明, 董志强, 等. 作物高产群体LAI动态模拟模型的建立与检验[J]. 作物学报, 2007, 33(4): 612–619 Zhang B, Zhao M, Dong Z Q, et al. Establishment and test of LAI dynamic simulation model for high yield population[J]. Acta Agronomica Sinica, 2007, 33(4): 612–619
[21] 国家烟草专卖局. YC/T 142—2010 烟草农艺性状调查测量方法[S]. 北京: 中国标准出版社, 2010 State Tobacco Monopoly Administration. YC/T 142—2010 Investigating and measuring methods of agronomical character of tobacco[S]. Beijing: Standards Press of China, 2010
[22] 严定春, 朱艳, 曹卫星. 水稻栽培适宜品种选择的知识模型[J]. 南京农业大学学报, 2004, 27(4): 20–25 Yan D C, Zhu Y, Cao W X. A knowledge model for selection of suitable variety in rice production[J]. Journal of Nanjing Agricultural University, 2004, 27(4): 20–25
[23] Maddonni G A, Otegui M E, Cirilo A G. Plant population density, row spacing and hybrid effects on maize canopy architecture and light attenuation[J]. Field Crops Research, 2001, 71(3): 183–193
[24] 孙锐, 朱平, 王志敏, 等. 春玉米叶面积系数动态特征的密度效应[J]. 作物学报, 2009, 35(6): 1097–1105 Sun R, Zhu P, Wang Z M, et al. Effect of plant density on dynamic characteristics of leaf area index in development of spring maize[J]. Acta Agronomica Sinica, 2009, 35(6): 1097–1105
[25] 杨慧芹, 王莎莎, 金云峰, 等. 生长温度对不同生育期烟草多酚物质代谢的影响[J]. 基因组学与应用生物学, 2015, 34(9): 1957–1974 Yang H Q, Wang S S, Jin Y F, et al. Effects of different growth temperature on polyphenols metabolism in tobacco (L.) leaves at different growth stages[J]. Genomics and Applied Biology, 2015, 34(9): 1957–1974
[26] Zhou Y H, Huang L F, Zhang Y L, et al. Chill-induced decrease in capacity of RuBP carboxylation and associated H2O2accumulation in cucumber leaves are alleviated by grafting onto figleaf gourd[J]. Annals of Botany, 2007, 100(4): 839–848
[27] Zhang C J, de A Prado C H B, Zu Y G, et al. Effect of overnight temperature on leaf photosynthesis in seedlings ofKing[J]. Journal of Forestry Research, 2003, 14(2): 130–134
[28] 官宇, 吴昊, 胡建新, 等. 攀枝花烟区不同基因型烤烟的光合特性研究[J]. 西南农业学报, 2013, 26(6): 2558–2561 Guan Y, Wu H, Hu J X, et al. Study on photosynthetic characteristics of different tobacco genotypes in Panzhihua tobacco area[J]. Southwest China Journal of Agricultural Sciences, 2013, 26(6): 2558–2561
[29] 聂志刚, 李广. 光照和温度对旱地小麦叶面积指数的影响[J]. 干旱区研究, 2013, 30(5): 894–898 Nie Z G, Li G. Effects of light and temperature on leaf area index of wheat in dryland[J]. Arid Zone Research, 2013, 30(5): 894–898
[30] 吕新. 生态因素对玉米生长发育影响及气候生态模型与评价系统建立的研究[D]. 泰安: 山东农业大学, 2002: 20–25 Lü X. Studies on effects of ecological factors on growth of maize and establishment of climate ecology model and appraisement system[D]. Tai’an: Shandong Agricultural University, 2002: 20–25
[31] 黄丽芬, 董芙蓉, 霍中洋, 等. 氮素水平对不同氮效率基因型水稻的物质生产与分配的影响[J]. 核农学报, 2012, 26(9): 1290–1297 Huang L F, Dong F R, Huo Z Y, et al. Effects of nitrogen levels on dry matter accumulation and distribution in rice genotype with different nitrogen use efficiencies[J]. Acta Agriculturae Nucleatae Sinica, 2012, 26(9): 1290–1297
[32] 杜薇, 王昌全, 李冰, 等. 攀枝花烟区不同施氮量对云烟85生长和产质量的影响[J]. 西南农业学报, 2012, 25(1): 193–197 Du W, Wang C Q, Li B, et al. Effects of different nitrogen rate on Yunyan85’s growth, yield and quality in Panzhihua tobacco area[J]. Southwest China Journal of Agricultural Sciences, 2012, 25(1): 193–197
[33] 易迪, 易镇邪, 屠乃美. 施氮量与留叶数对烤烟光合特性的影响[J]. 中国农学通报, 2014, 30(10): 99–104 Yi D, Yi Z X, Tu N M. Effect of nitrogen application and remained leaf number on photosynthetic characteristics of flue-cured tobacco[J]. Chinese Agricultural Science Bulletin, 2014, 30(10): 99–104
[34] 胡国松, 郑伟, 王震东, 等. 烤烟营养原理[M]. 北京: 科学出版社, 2000: 57–61 Hu G S, Zheng W, Wang Z D, et al. Nutrition Principle of Flue Cured Tobacco[M]. Beijing: Science Press, 2000: 57–61
[35] 李向岭, 李从锋, 侯玉虹, 等. 不同播期夏玉米产量性能动态指标及其生态效应[J]. 中国农业科学, 2012, 45(6): 1074–1083 Li X L, Li C F, Hou Y H, et al. Dynamic characteristics of summer maize yield performance in different planting dates and its effect of ecological factors[J]. Scientia Agricultura Sinica, 2012, 45(6): 1074–1083
Dynamic simulation of leaf area index of tobacco based on nitrogen effect*
SHAO Huifang1, LIU Zhihong1, CUI Dengke2, MU Tong1, HUANG Wuxing1, XU Zicheng1
(1. College of Tobacco Science, Henan Agricultural University, Zhengzhou 450002, China; 2. Baofeng Tobacco Redrying Factory of Tianchang International Co. Ltd., Pingdingshan 467400, China)
This study was set up to clarify the dynamic characteristics of leaf area index of flue-cured tobacco under different nitrogen levels and the relationship with accumulated temperature. The leaf area index and dynamic characteristics of three flue-cured tobacco cultivars (‘YU12’, ‘QIN96’ and ‘YUN87’) were measured under different nitrogen levels (0 kg·hm-2, 30 kg·hm-2, 60 kg·hm-2and 90 kg·hm-2) to improve photosynthetic structure of flue-cured tobacco. Using the Curve Expert 1.40 software and the limit method, the normalized accumulated temperature model was established. The result indicated that: 1) A unimodal curve between leaf area index and accumulated temperature was observed after transplanting of flue-cured tobacco, with a skewness less than 0. Leaf area index of flue-cured tobacco increased with increasing nitrogen level, and the peak value of leaf area index was in the order of ‘QIN96’ > ‘YUN97’ > ‘YU12’ at the same nitrogen level. 2) The rational function model= (+) / (1++2) yielded the best simulation result of leaf area index change with relative active accumulated temperature with high precision level (2= 0.980 7**). The accuracy and precision of the normalized model were tested with data for 2015, and the simulation accuracy () was all above 0.958, the precision (2) all above 0.95 and RMSE below 6.04%. 3) There were significant differences among some varieties and nitrogen levels, showing that variety interaction with nitrogen level mainly changed the model values of,and. 4) The velocity of relative leaf area index change during the growing period showed trends in ‘N’ curve, reflecting the actual variability of leaf area index. 5) Secondary indexes were more sensitive to different nitrogen levels. The mean leaf area index and maximum leaf area index increased with increasing nitrogen level. These characteristic indexes of leaf area index could be served as a reference base for the regulation of plant nitrogen of flue-cured tobacco population. The model provided theoretical basis and decision support for monitoring growth dynamics and photosynthetic characteristics of flue-cured tobacco population.
Flue-cured tobacco; Leaf area index; Active accumulated temperature; Nitrogen application level; Rational function model
SHAO Huifang, E-mail: shf.email@163.com
Mar. 13, 2017; accepted May 2, 2017
S572
A
1671-3990(2017)09-1276-11
10.13930/j.cnki.cjea.170214
* 河南省烟草公司科技攻关项目(HYKJ201405)资助
2017-03-13
2017-05-02
邵惠芳, 主要研究方向为烟叶质量评价和数据挖掘。E-mail: shf.email@163.com
* This work was supported by the Scientific Program of Henan Provincial Tobacco Company (HYKJ 201405).
邵惠芳, 刘志宏, 崔登科, 穆童, 黄五星, 许自成. 基于氮素效应的烤烟叶面积指数动态模拟[J]. 中国生态农业学报, 2017, 25(9): 1276-1286
Shao H F, Liu Z H, Cui D K, Mu T, Huang W X, Xu Z C. Dynamic simulation of leaf area index of tobacco based on nitrogen effect[J]. Chinese Journal of Eco-Agriculture, 2017, 25(9): 1276-1286
猜你喜欢
中国农学通报(2022年5期)2022-03-02
中国土壤与肥料(2021年5期)2021-12-02
养殖与饲料(2021年4期)2021-04-17
现代农业科技(2020年4期)2020-04-08
上海农业学报(2017年4期)2017-04-10
中低纬山地气象(2016年6期)2016-07-31
农家顾问(2014年9期)2014-10-20
植物营养与肥料学报(2014年1期)2014-03-11
植物营养与肥料学报(2014年1期)2014-03-11
中低纬山地气象(2013年5期)2013-09-29
