
基于结构方程模型分析荒漠啮齿动物优势种对不同放牧干扰的响应
2017-08-28 15:14:37付和平武晓东甘红军岳秀贤
生态学报 2017年14期
袁 帅,付和平,武晓东,*,兴 安,甘红军,岳秀贤
1 内蒙古农业大学草原与资源环境学院, 呼和浩特 0100112 草业与草地资源教育部重点实验室, 呼和浩特 0100113 内蒙古阿拉善盟草原站, 巴彦浩特 7503064 内蒙古自治区林业监测规划院, 呼和浩特 010020
基于结构方程模型分析荒漠啮齿动物优势种对不同放牧干扰的响应
袁 帅1,2,付和平1, 2,武晓东1,2,*,兴 安1, 2,甘红军3,岳秀贤4
1 内蒙古农业大学草原与资源环境学院, 呼和浩特 0100112 草业与草地资源教育部重点实验室, 呼和浩特 0100113 内蒙古阿拉善盟草原站, 巴彦浩特 7503064 内蒙古自治区林业监测规划院, 呼和浩特 010020
放牧被认为是草地生态系统的主要干扰之一。许多研究表明,放牧影响了啮齿动物的个体条件、繁殖、种群动态、群落组成及群落多样性等多个方面。由于受传统统计学方法的限制,这些研究只是单独的对放牧作用进行假设检验,而没有将多个假设放在同一个框架内进行验证。2006—2011年,采用标志重捕法对内蒙古阿拉善荒漠区不同放牧生境中啮齿动物优势种的数量进行研究并使用结构方程建模的方法将可能影响荒漠啮齿动物优势种的环境因子(气候条件、土壤硬度、植物隐蔽、植物生物量等19个指标)与啮齿动物优势种种群动态结合后进行多假设检验,旨在找出不同放牧干扰下制约优势啮齿动物环境因子的发生途径。结果表明:气候条件对五趾跳鼠和三趾跳鼠有直接不利影响,隐蔽对于两足活动的五趾跳鼠有负效应,而对四足活动的子午沙鼠有正效应。植物生物量对五趾跳鼠有正效应,但对子午沙鼠有负效应。土壤硬度的增加间接的有利于五趾跳鼠,对子午沙鼠有直接的正效应。因此荒漠啮齿动物优势种种群动态是多个过程共同作用的结果,气候条件是驱动啮齿动物种群动态的根本原因,而放牧会因为改变植物隐蔽而影响啮齿动物在群落中的组成。
啮齿动物, 种群动态, 放牧干扰, 结构方程
放牧对干旱和半干旱区的草原影响远大于其他任何一种土地利用方式[1]。过度放牧使草地生态系统在组成、结构和功能上产生了较高生态代价[2]。啮齿动物是草地生态系统的重要组成部分[3],有关放牧对啮齿动物的影响国内外已经集中了大量研究[4- 6]。现有的研究表明,放牧影响了啮齿动物的个体体质[4]、种群动态[6]、繁殖[7]、群落组成[8]及群落多样性等多个方面[9]。虽然这些研究大多认为放牧干扰对啮齿动物作用途径并不单一,但是受传统统计学方法的限制,这些研究只是单独的对放牧作用进行假设检验,而没有将多个假设放在同一个框架内进行验证。而使用结构方程建模(Structural Equation Modeling,SEM)可以很好的解决这些问题[10]。
SEM是通过使用变量的协方差矩阵来分析变量间关系的一种多变量统计方法[11]。SEM模型可以评估有关生态系统的复杂多变量假说。打破了传统单变量假说测试框架对研究相互制约系统的限制[10]。本文通过对阿拉善荒漠区啮齿动物优势种及其环境因子关系的研究,旨在找出不同放牧干扰下制约优势啮齿动物环境因子的发生途径,以期为荒漠生态系统小哺乳动物多样性的维持以及荒漠区鼠害生态控制提供一些科学依据。
1 研究区概况及研究方法
1.1 研究区概况
研究区(104°10′—105°30′E, 37°24′—38°25′N)位于内蒙古自治区阿拉善左旗南部,腾格里沙漠东缘。气候为典型高原大陆性气候。极端最低温和极端最高温分别为-36 ℃和42 ℃。年降雨量45—215 mm,约70%的降雨集中在6—9月。无霜期156 d。地形起伏不平,丘陵、沙丘与平滩相间。土壤为灰漠土和灰棕土。草地类型为沙质荒漠亚类属于温性荒漠类型。植物建群种以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygophyllaceae)为主,其次为蔷薇科(Rosaceae)、柽柳科(Tamaricaceae)。在此本底生境的基础上,按照土地利用方式的不同,选择禁牧区、轮牧区和过牧区3种不同的生境作为取样区(表1)。
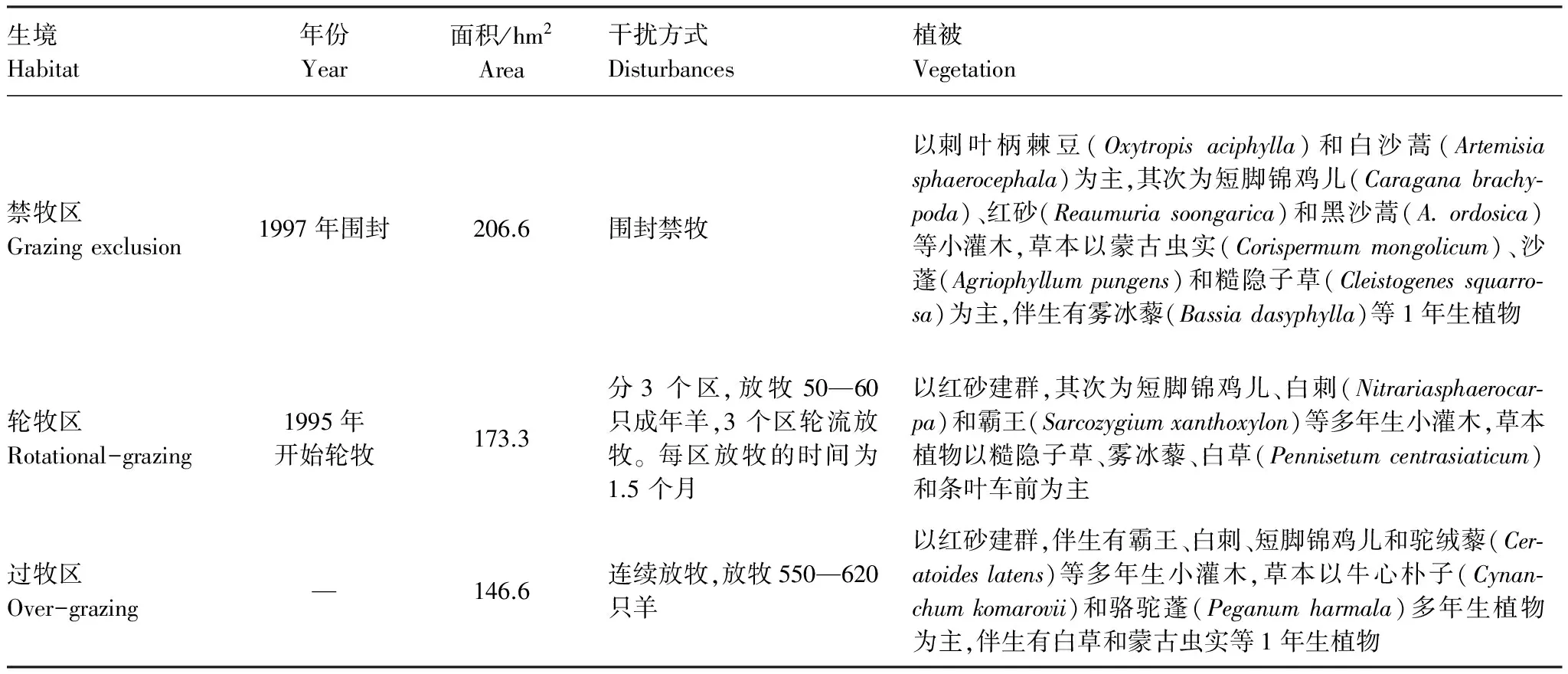
表1 研究区各放牧干扰生境的基本情况
1.2 研究方法
1.2.1 数据采集方法
(1)啮齿动物调查
在每种生境中各选择2个取样区,在各取样区中心建立1个0.96 hm2固定的标志重捕样地。在每个样地中按7 × 8的方格布笼,每个网格点上放置1个活捕笼,行距及笼距均为15 m。使用新鲜花生作诱饵。2006—2011年每年的4—10月每月进行标记重捕,每次连捕4 d。记录捕获个体的种名、性别、繁殖状况和捕获位置,称量捕获个体体重,并对每个捕获个体进行标记,除跳鼠科啮齿动物用脚环标记外,其它啮齿动物均使用剪趾法标记[9]。
(2)植物调查
2006—2011年每年4—10月进行植物数据采集,每月月初进行植物取样。在每个动物取样样区内随机选取3个10 m×10 m的样方进行灌木取样,在每个灌木样方中随机选取3个1 m×1 m的样方作为草本样方。在这些样方中测量植物的“四度一量”。
(3)土壤数据采集
土壤取样在每个灌木样方内测量0—5、5—10、10—15 cm和15—20 cm土壤的硬度。土壤硬度使用土壤硬度仪(1Psi=6.89KP)测定。
(4)气象资料收集
气象资料均来自“阿拉善盟孪井滩气象站”。气象资料包括2006—2011年4—10月降雨量、气温和相对湿度等指标。
1.2.2 数据分析
(1)种群数量计算
①捕获量比例(Pi)
第i个物种的捕获数量与其所在群落中动物总捕获数量的比值。
②最小存活法
使用最小已知存活数(Minimum number known alive,MNA)[12]来表示各优势种群大小,最小已知存活数包括两个部分:a.在t时刻所捕获的啮齿动物数量;b.在t时刻之前标记,在t时之后出现,但不出现在t时刻的啮齿动物数量。这两部分之和就是MNA。
将同一干扰下各鼠种的捕获量比例在混合效应模型中进行了差异性检验。所有分析用SAS 9.2软件完成,显著水平为α=0.05,应用Tukey检验进行均值比较。
(2)响应机制验证
X=Λxξ+δ
Y=Λyη+ε
η=Bη+Γξ+ζ
式中,Λx和Λy表示指标在潜变量上的因子载荷矩阵;η为内生潜变量;ξ表示外源潜变量;B表示内生潜变量间的关系;Γ表示外源潜变量对内生潜变量的影响;δ为外源指标的误差项;ε表示内生指标的误差项;ζ为结构方程的残差项。
本研究使用两阶段策略(Two-stage modeling)构建全模型。第一阶段为验证性因子分析(Confirmatory Factor Analysis,CFA)。第二阶段为结构模型拟合[13]。使用极大似然法(Maximum likelihood,ML)进行参数估计[11]。本研究选用卡方自由度比、近似误差均方根(Root Mean Square Error of Approximation, RMSEA)、非范拟合指数(Non-Normed Fit Index, NNFI)和比较拟合参数(Comparative Fit Index,CFI)进行拟合优度检验。其中卡方自由度比小于2为拟合良好,2—5之间为可接受模型。RMSEA小于0.1为好的拟合,且越小效果越好。NNFI和CFI越接近1表示拟合越好。潜变量与测量指标之间的关系用因素载荷来表示;潜变量之间的关系用回归系数表示;所有路径系数均使用完全标准化解。当第一阶段的CFA模型拟合良好后,才可进行第二阶段的全模型构建。所有结构方程模型的分析均使用Lisrel8.7完成。
2 结果与分析
2.1 优势种的确定
2006—2011年共布放活捕笼56448笼日, 捕获啮齿动物3科7属9种。其中跳鼠科(Dipodidae)3种,包括:三趾跳鼠(Dipussagitta)、五趾跳鼠(Allactagasibirica)和蒙古羽尾跳鼠(Stylodipusandrewsi);仓鼠科(Cricetidae)5种,包括:子午沙鼠(Merionesmeridianus)、长爪沙鼠(M.unguiculatus)、小毛足鼠(Phodopusroborovskii)、黑线仓鼠(Cricetulusbarabensis)以及短尾仓鼠(C.eversmanni);松鼠科(Sciuridae)1种为:阿拉善黄鼠(Spermophilusalaschanicus)。
本研究依据啮齿动物捕获量比例大小确定优势种。对相同干扰下不同鼠种6a捕获数据的方差分析表明禁牧区优势鼠种为三趾跳鼠,次优势鼠种为子午沙鼠和小毛足鼠;轮牧区优势鼠种为三趾跳鼠,次优势鼠种为五趾跳鼠、小毛足鼠和子午沙鼠;过牧区优势鼠种为五趾跳鼠,次优势种为三趾跳鼠(表2)。
2.2 优势种种群动态
本研究使用MNA对啮齿动物优势种的种群数量动态进行了分析。子午沙鼠数量高峰在禁牧、轮牧和过牧区均出现在2008年。其中禁牧区的子午沙鼠种群数量高于轮牧区和过牧区,但是3个区子午沙鼠种群数量的变化趋势基本相同。3个干扰区内三趾跳鼠种群动态总体趋势类似,禁牧区和过牧区三趾跳鼠种群数量高峰均出现在2008年。禁牧与轮牧干扰下的三趾跳鼠种群数量高于过牧区。过牧区的五趾跳鼠种群数量在2008—2010年保持着数量高峰,轮牧区高峰出现在2010年。禁牧区2009年后种群数量出现上升(图1)。
The influence study of complex consolidation on stability for high slag slope WANG Xiu-feng LIU Wen-lian LIAO Shao-zhong et al.(113)
2.3 不同干扰对优势鼠种作用机理
本研究使用结构方程模型(SEM)来检验不同干扰对啮齿动物优势种的作用途径。由于本底生境相似,所以过牧、轮牧和禁牧合并建模。
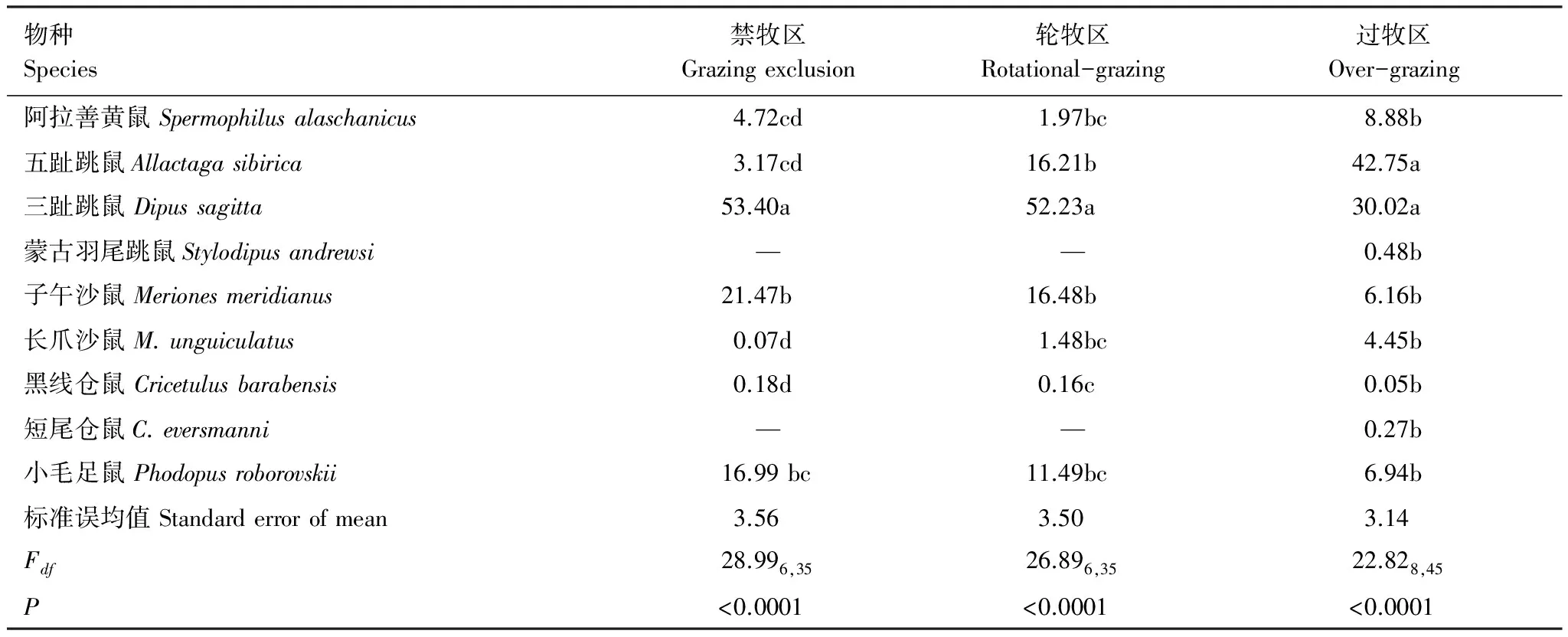
表2 不同干扰下各鼠种6年总捕获量比例%

图1 不同干扰下优势种群数量年动态Fig.1 Annual dynamic of dominant species population subjected to different disturbances
2.3.1 概念模型构建
之前的研究已经表明啮齿动物种群可能受到来自气候、土壤硬度和植物等方面的影响[14-16],因此选择衡量动物优势种种群数量、气候条件、土壤硬度、植物隐蔽和植物生物量的19个惯用指标进行结构方程模型的构建。测量指标与描述统计量如表3所示。
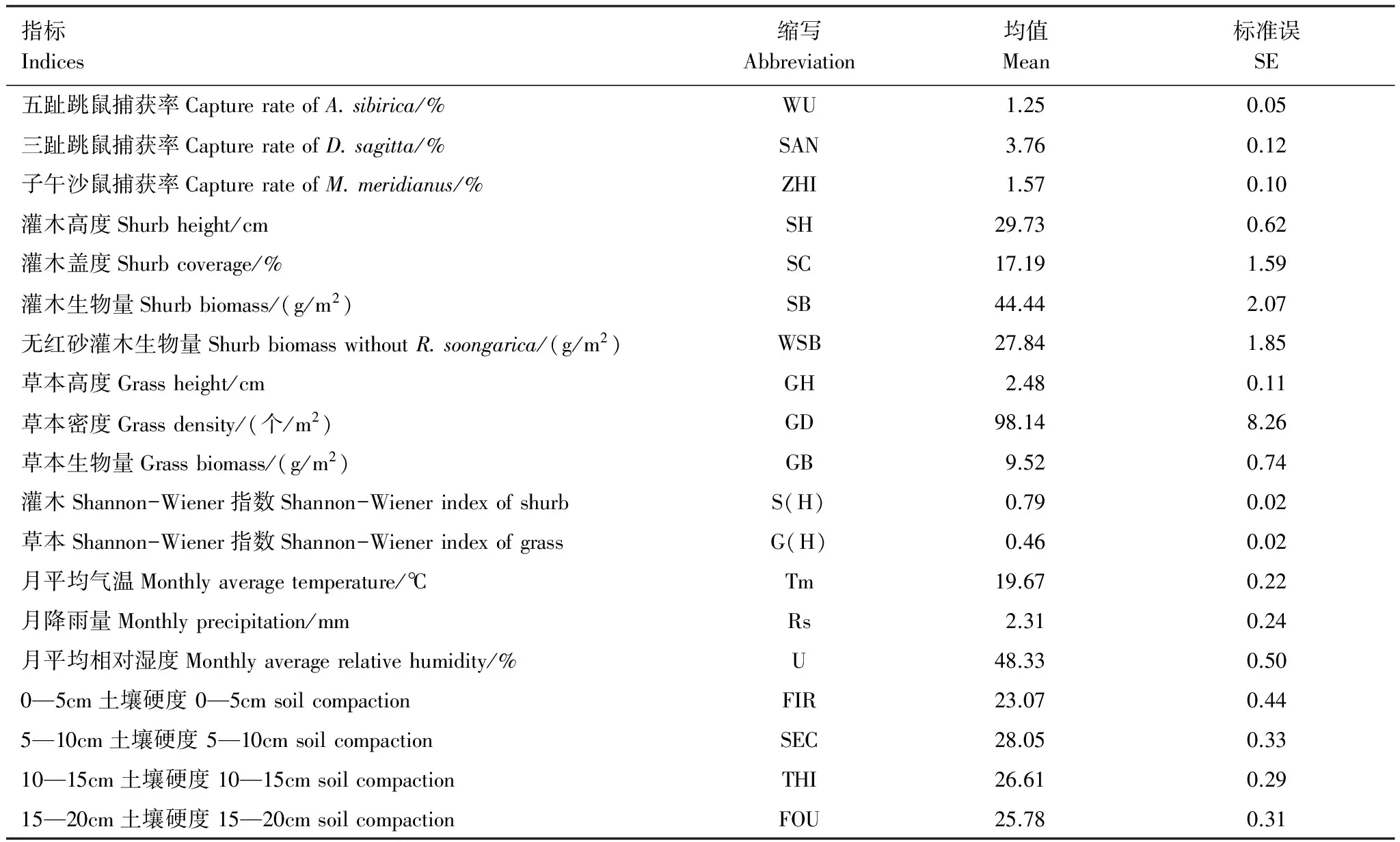
表3 啮齿动物优势种结构方程模型(SEM)测量指标
SE为标准误
假设五趾跳鼠(WUZHI)、三趾跳鼠(SANZHI)、子午沙鼠(ZIWU)、植物隐蔽(SHIELD)和植物生物量(PBIO)作为内生潜变量,气候条件(CLI)和土壤硬度(SOIL)作为外源潜变量。外源潜变量气候条件由月平均气温(Tmean)、月降雨量(Rsum)、月平均湿度(U)说明;土壤硬度由0—5cm(FIR)、5—10cm(SEC)、10—15cm(THI)和15—20cm(FOU)的土层硬度来说明。内生潜变量五趾跳鼠、三趾跳鼠和子午沙鼠分别由单一指标五趾跳鼠捕获率(WU)、三趾跳鼠捕获率(SAN)和子午沙鼠捕获率(ZHI)来说明。内生潜变量植物隐蔽由指标灌木高度(SH)、灌木盖度(SC)、灌木Shannon-Wiener指数(S(H))、草本Shannon-Wiener指数(G(H))、草本高度(GH)、草本密度(GD)来说明。内生潜变量植物生物量由灌木生物量(SB)、无红砂灌木生物量(WSB)和草本生物量(GB)表示。根据上述假设及所提及的各变量的测量指标构建概念模型,该模型共有19个测量变量,样本量为424(图2)。
2.3.2 全模型构建
经CFA拟合修正后[WLS=481.04(df=124,P<0.01),卡方自由度比为3.88,RMSEA=0.083,NNFI=0.87,CFI=0.91],因素载荷良好,验证模型拟合基本理想进行全模型构建。全模型最终拟合结果表明,WLS卡方为480.79(df=123,P<0.01),卡方自由度比为3.91,小于5为可接受模型。RMSEA为0.083,小于0.1。NNFI(0.87)和CFI(0.90)略小,但较接近1,以上指标表明模型拟合基本理想(图3)。各指标对潜变量的载荷和残差表明,除灌木盖度在植物隐蔽上的载荷和灌木高度在植物生物量上的载荷较小外,其它测量指标载荷均相对较高。所有测量指标载荷均达到统计显著(表4,图3)。其中灌木高度在植物隐蔽上的载荷最大(0.77);在植物生物量上载荷最大的为草本生物量(0.83);在气候条件上载荷最大的为(0.98);土壤的后三层硬度对土壤硬度的载荷较大,均在0.70以上(表4)。

图2 过牧、禁牧和轮牧区优势鼠种对干扰响应的概念模型Fig.2 Conceptual model representing hypothesized relationships between biotic and abiotic factors and dominant species in treatment areasWU:五趾跳鼠数量;SAN:三趾跳鼠数量;ZHI:子午沙鼠数量; SH:灌木高度; SC:灌木盖度; S(H):灌木Shannon-Wiener指数; G(H):草本Shannon-Wiener指数;GH:草本高度; GD:草本密度; SB:灌木生物量; WSB:除去红砂后灌木生物量; GB:草本生物量; Tmean:月平均温度; Rsum:月降雨量; U:月平均湿度; FIR:0—5cm土壤硬度; SEC:5—10cm土壤硬度; THI:10—15cm土壤硬度; FOU:15—20cm土壤硬度;WUZHI:潜变量五趾跳鼠;SANZHI:潜变量三趾跳鼠;ZIWU:潜变量子午沙鼠;CLI:潜变量气候条件;SOIL:潜变量土壤硬度;SHIELD:潜变量隐蔽; PBIO:潜变量植物生物量;方框:测量指标;圆圈:潜变量;单箭头:单向影响;双向箭头:相关

表4 过牧、禁牧和轮牧区优势鼠种SEM模型各测量指标因素载荷
*:t值大于1.96,P<0.05;**:t值大于2.58,P<0.01;***:t值大于3.29,P<0.001
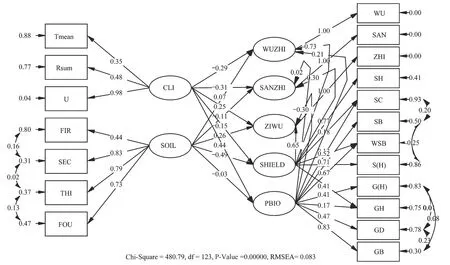
图3 过牧、禁牧和轮牧区优势鼠种全模型最终解路径图Fig.3 Final results of full model for relationships between biotic and abiotic factors and dominant species in treatment areas图中所有路径的系数均为完全标准化系数,“-”表示负效应;其它同图2
潜变量的关系:植物隐蔽对五趾跳鼠和子午沙鼠的捕获率直接影响最大,分别达到了-0.73(t=-7.95,P<0.001)和0.65(t=6.33,P<0.001,表5)。植物生物量对三趾跳鼠的捕获率影响较弱,且统计不显著(t=0.26,P>0.05)。土壤对植物生物量的影响也不明显(t=-0.03,P>0.05)。气候条件与土壤硬度存在显著的负相关(由于外源潜变量之间不能用单箭头连接,所以二者之间的关系为相关);植物隐蔽与植物生物量之间显著正相关;五趾跳鼠与三趾跳鼠和子午沙鼠捕获率之间无显著相关;但三趾跳鼠与子午沙鼠捕获率显著正相关(图4)。
各潜变量之间的效应分析表明:对于五趾跳鼠种群数量,对其整体影响最大的是植物隐蔽;气候条件对其有显著的直接(t=-4.11,P﹤0.001)和间接效应(t=3.09,P﹤0.01),但整体效应并不显著(t=-0.09,P>0.05);土壤硬度对其有显著的间接效应(t=5.66,P﹤0.001);植物隐蔽(t=-7.95,P﹤0.001)和生物量(t=2.44,P﹤0.01)对其均有显著的直接效应。对于三趾跳鼠种群数量,对其整体影响最大的是气候条件,其次为植物隐蔽;气候条件(t=-4.24,P﹤0.001)和植物隐蔽(t=3.53,P﹤0.001)对其有显著直接效应;土壤硬度(t=-3.41,P﹤0.001)对其有间接负效应。对于子午沙鼠种群数量,对其整体影响最大的为植物隐蔽;气候条件对其有显著的直接(t=3.13,P﹤0.01)和间接(t=-3.32,P﹤0.001)效应,但整体效应并不显著(t=0.34,P>0.05);土壤硬度对子午沙鼠也具有显著的直接(t=3.26,P﹤0.01)和间接(t=-4.70,P﹤0.001)效应,但无显著的整体效应(t=-0.05,P>0.05);植物隐蔽(t=6.33,P﹤0.001)和植物生物量(t=-3.14,P﹤0.01)对子午沙鼠种群数量有显著直接效应(表5)。
过牧、禁牧和轮牧区:对于五趾跳鼠种群数量而言,对其最大的直接影响来自植物隐蔽;对其最大的间接效应来自土壤硬度,对其整体影响最大的是植物隐蔽。对于三趾跳鼠种群数量而言,对其最大的直接正效应来自气候条件和植物隐蔽;对其最大的间接负效应来自土壤硬度;对其影响最大的整体效应是气候条件,其次为植物隐蔽。对于子午沙鼠种群数量而言,最大的直接正效应和负效应分别来自植物隐蔽和植物生物量;最大的间接负效应来自土壤硬度;但对其整体影响最大的是植物隐蔽。
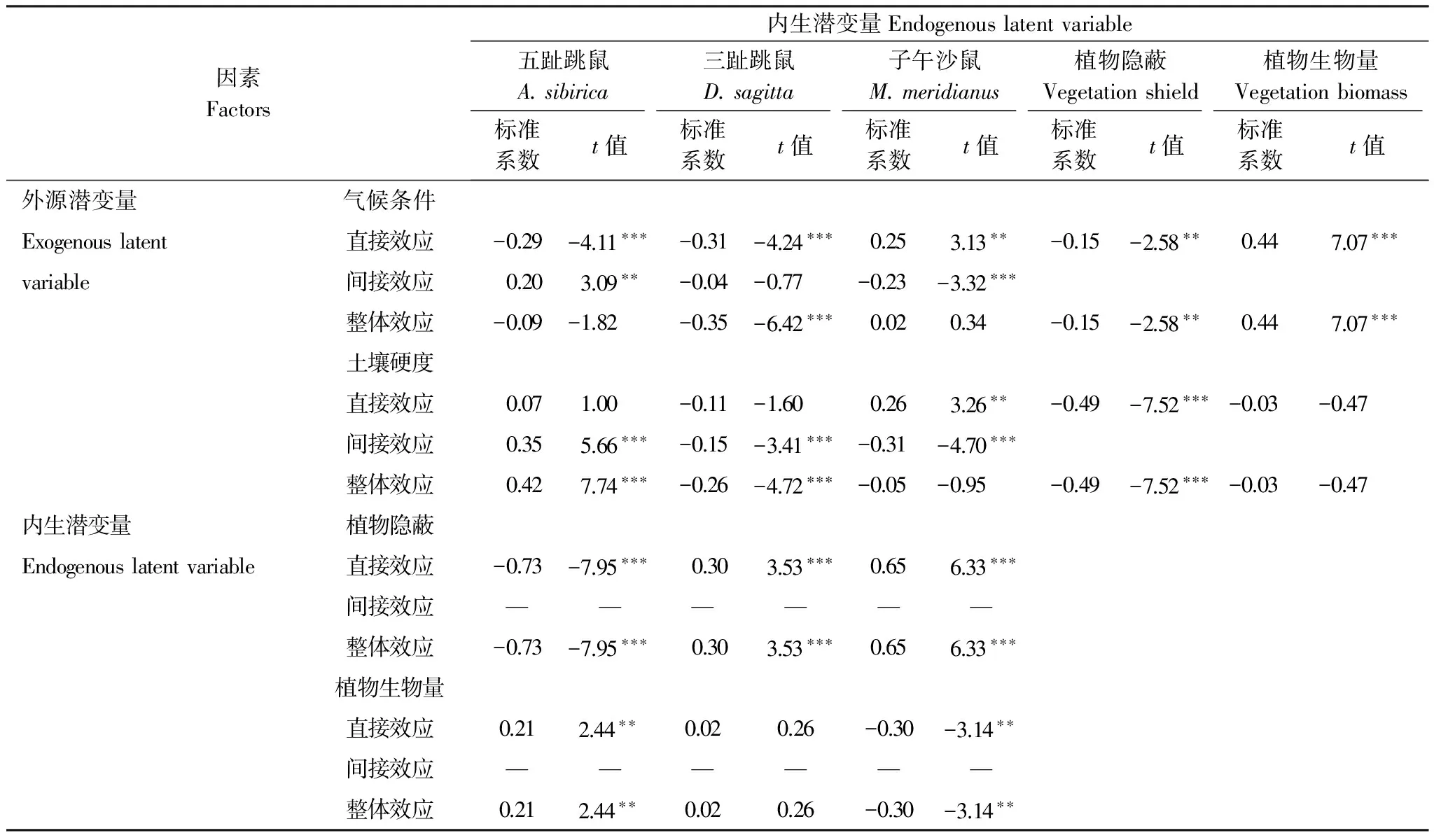
表5 过牧、禁牧和轮牧区优势鼠种SEM全模型潜变量效应分析
*t值大于1.96,P<0.05;**t值大于2.58,P<0.01;***t值大于3.29,P<0.001
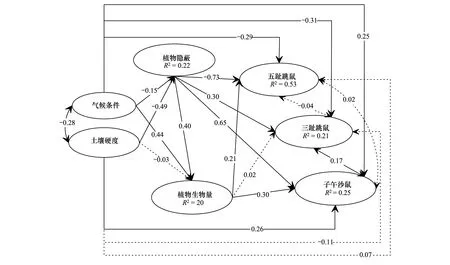
图4 过牧、禁牧和轮牧区优势鼠种全模型潜变量关系路径图Fig.4 Relationship between latent variables of full model for dominant species in treatment areas圆圈:潜变量;单箭头:单向效应;双箭头:相关;实体黑线:统计显著;虚线:统计不显著;图中所有路径的系数均为完全标准化系数,“-”:负效应
3 讨论与结论
3.1 气候与种群动态
气候变化影响了动物的地理分布、物候、个体行为以及种群动态等[17- 19]。阿拉善荒漠区5—9月的气候条件对五趾跳鼠和三趾跳鼠有直接不利影响。荒漠啮齿动物夜行性的目的之一是为了躲避高温,这是长期适应荒漠气候的结果[20]。五趾跳鼠和三趾跳鼠主要在夜间活动,白天则入洞休息[21]。但由于两种跳鼠在植物生长季所居洞穴较浅且结构简单[22],荒漠地区白天较高的气温会使距地表较浅的洞穴内部温度升高,而降雨量的增加也可能会直接入渗到跳鼠的洞中,对跳鼠产生不利影响。虽然子午沙鼠也为夜行性鼠种,但是其洞穴与三趾跳鼠和五趾跳鼠相比距地面较深,并且更加复杂[23-24]。这种情况可以更好的规避气候条件直接带来的不利影响。
3.2 植物与种群动态
浓密的植物隐蔽不利于两足活动的啮齿动物取食和躲避敌害,但有利于四足活动的啮齿动物生存[25- 27]。SEM的结果映证了这一结论,隐蔽对于两足活动的五趾跳鼠有负效应,而对四足活动的子午沙鼠有正效应。一些文献常提及三趾跳鼠喜欢开阔生境[22],但是对于隐蔽与三趾跳鼠的关系多是描述性的,并没直接的数据支持。本研究SEM的结果表明,隐蔽对三趾跳鼠表现出正效应,因此在一定的范围内二者关系为正相关,但三趾跳鼠与隐蔽之间可能是一种非线性的关系。
植物生物量对五趾跳鼠有正效应,但对子午沙鼠有负效应。这种关系可能是由于两种啮齿动物对食物的可用性要求不同导致的。Ernest等[28]的研究发现,一些啮齿动物的丰富度与初级生产力有关。而Arsenault和Smith[29]认为小型啮齿动物需要较高质量的食物而不是高数量的食物。二者之间可能并不矛盾,只是动物本身的生物学和进化史特征决定了对食物可用性的需求不同而已。李俊生[30]的研究结果表明体型较大的跳鼠具有相对较长的盲肠长度,这种特点使其可以从低质量的牧草中摄取营养物[31-32],因而降低了其对食物的选择性,从而适应低质量的生境。对食物的低选择性,一方面可补偿食谱食物减少带来的影响,另一方通过增加对食物的非选择性,可以降低天敌对啮齿动物的捕食风险[33]。
3.3 土壤硬度与种群动态
土壤硬度的增加不利于植物隐蔽的增加。一方面,土壤硬度的增加可能对植物本身存在不利影响,包括影响幼苗的建立、植物的生根等,另一方面较高的土壤硬度也意味着较高的放牧压力。这两方面都可能导致植物隐蔽的简化,从而间接的有利于喜欢开阔生境的五趾跳鼠,而不利于喜欢高隐蔽的子午沙鼠。对于子午沙鼠来说,土壤硬度的增加虽然会增加其挖掘时的体力消耗,但是硬度增加会减少洞道的坍塌[34],这就为对洞道结构要求较高的子午沙鼠提供了有利条件,因而土壤硬度对子午沙鼠有直接的正效应。
3.4 SEM与啮齿动物种群动态
物种的种群数量通常受到生物和非生物因素的直接和间接影响。而理解物种出现的驱动因素是基础生态学和应用生态学的根本目标,但很多统计处理一般无法区分直接和间接效应[35]。结构方程分析的出现为解决这些问题提供了有利途径[10]。与传统的统计方法相比,SEM模型具有多个优点如[11]:a.可同时处理多个因变量;b.允许自变量和因变量含测量误差;c.可同时评估因子结构和因子关系;d.可对整个模型的拟合效果给出多种拟合优度评估。此外,SEM还可通过路径图表示诸如群落构建复杂过程中的直接和间接效应[36],通过明确的描述多变量间的因果关系,变量间的机制关系变得更加清晰,同时要比二元统计方法更具预测能力[37]。因此,SEM模型允许生态学家评估有关生态系统的复杂多变量假说。这种优点打破了传统单变量假说测试框架对研究相互制约系统的限制[10]。SEM提供了阐明、表达和评估生态学假说的框架,这一框架优于传统的相关统计方法[35]。啮齿动物种群不仅受到气候、土壤、植物隐蔽、食物可用性、捕食风险甚至人为干扰造成的大尺度上的景观异质性的影响,而且物种的生物学特征、生活史策略以及种间关系也对其产生很大的作用。所以啮齿动物种群生态学研究是一个围绕动物种群的多变量研究。在这样的研究中,挖掘潜藏在动物种群数量波动表面现象之下的运行机制是人们最为关心的问题,也是对于生产实践指导最具实际意义的问题。
综上所述,不同干扰下啮齿动物优势种的种群数量受到多个过程的共同作用。从整体效应来看,植物隐蔽在过牧、禁牧和轮牧区的优势鼠种种群数量的调节中占主导地位。由不同放牧程度导致的植物隐蔽情况变化,使五趾跳鼠在隐蔽较差的过牧区成为优势种,而三趾跳鼠和子午沙鼠在隐蔽较好的轮牧和禁牧区成为优势种。荒漠区气候条件是发生在较大尺度上的自然干扰事件,其直接和间接的影响啮齿动物种群数量,而啮齿动物通过长期进化来适应这些影响。因而形成了荒漠啮齿动物自身的特有的生物学和生活史特征(如夜行性,跳跃活动,听泡增大等)[20]。这些特征对生境适宜性提出了不同的要求,而人为干扰又在时空尺度上创造或扰动了生境的多样性和异质性。因此啮齿动物的自身特征和外部干扰共同决定了它们在不同生境中的停留、续存或灭绝,最终导致了不同生境中优势种的出现。这些因素决定了啮齿动物优势种的种群数量是多个因子间复杂作用的结果。因此,使用SEM研究荒漠区啮齿动物优势种及其环境因子关系,了解制约啮齿动物优势种种群动态的环境因子发生途径,对于减少鼠害防治中的无差别防治,维持荒漠生态系统小哺乳动物及上层肉食动物多样性及精准控制荒漠区鼠害具有一定的实践意义。
致谢:内蒙古农业大学张福顺、鄂晋、吉晟男、韩艳静等帮助收集野外数据,野外工作还得到内蒙古阿拉善盟草原站的帮助,特此致谢。
[1] Frank A S K, Dickman C R, Wardle G M, Greenville A C. Interactions of grazing history, cattle removal and time since rain drive divergent short-term responses by desert biota. PLoS One, 2013, 8(7): e68466.
[2] Fleischner T L. Ecological costs of livestock grazing in western North America. Conservation Biology, 1994, 8(3): 629- 644.
[3] Davidson A D, Detling J K, Brown J H. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world′s grasslands. Frontiers in Ecology and the Environment, 2012, 10(9): 477- 486.
[4] Keesing F. Impacts of ungulates on the demography and diversity of small mammals in central Kenya. Oecologia, 1998, 116(3): 381- 389.
[5] Yoshihara Y, Okuro T, Buuveibaatar B, Undarmaa J, Takeuchi K. Complementary effects of disturbance by livestock and marmots on the spatial heterogeneity of vegetation and soil in a Mongolian steppe ecosystem. Agriculture, Ecosystems & Environment, 2010, 135(1/2): 155- 159.
[6] Li G L, Yin B F, Wan X R, Wei W H, Wang G M, Krebs C J, Zhang Z B. Successive sheep grazing reduces population density of Brandt′s voles in steppe grassland by altering food resources: a large manipulative experiment. Oecologia, 2016, 180(1): 149- 159.
[7] Seldal T, Andersen K J, Högstedt G. Grazing-induced proteinase inhibitors: a possible cause for lemming population cycles. Oikos, 1994, 70(1): 3- 11.
[8] Lagesse J V, Thondhlana G. The effect of land-use on small mammal diversity inside and outside the great fish river nature reserve, Eastern Cape, South Africa. Journal of Arid Environments, 2016, 130: 76- 83.
[9] 袁帅, 武晓东, 付和平, 杨泽龙, 张福顺, 张晓东. 不同干扰下荒漠啮齿动物群落多样性的多尺度分析. 生态学报, 2011, 31(7): 1982- 1992.
[10] Grace J B, Bollen K A. Representing general theoretical concepts in structural equation models: the role of composite variables. Environmental and Ecological Statistics, 2008, 15(2): 191- 213.
[11] 侯杰泰, 温忠麟, 成子娟. 结构方程模型及其应用. 北京: 教育科学出版社, 2004.
[12] Krebs C J. Demographic changes in fluctuating populations of microtus californicus. Ecological Monographs, 1966, 36(3): 239- 273.
[13] 邱皓政, 林碧芳. 结构方程模型的原理与应用. 北京: 中国轻工业出版社, 2009.
[14] Dickman C R, Greenville A C, Tamayo B, Wardle G M, Blake B H. Spatial dynamics of small mammals in central Australian desert habitats: the role of drought refugia. Journal of Mammalogy, 2011, 92(6): 1193- 1209.
[15] Kutt A S, Thurgate N Y, Hannah D S. Distribution and habitat of the desert mouse (Pseudomysdesertor) in Queensland. Wildlife Research, 2004, 31(2): 129- 142.
[16] Morilhat C, Bernard N, Bournais C, Meyer C, Lamboley C, Giraudoux P. Responses ofArvicolaterrestrisschermanpopulations to agricultural practices, and toTalpaeuropaeaabundance in eastern France. Agriculture, Ecosystems & Environment, 2007, 122(3): 392- 398.
[17] Lutermann J S H, Alagaili A N, Mohammed O B, Bennett N C. Seasonal reproduction in the Arabian spiny mouse,Acomysdimidiatus(Rodentia: Muridae) from Saudi Arabia: the role of rainfall and temperature. Journal of Arid Environments, 2016, 124: 352- 359.
[18] Jiang G S, Liu J, Xu L, Yan C, He H L, Zhang Z B. Intra- and interspecific interactions and environmental factors determine spatial-temporal species assemblages of rodents in arid grasslands. Landscape Ecology, 2014, 30(9): 1643- 1655.
[19] 武晓东, 袁帅, 付和平, 张晓东, 张福顺, 高全荣. 不同干扰下阿拉善荒漠啮齿动物优势种对气候变化的响应. 生态学报, 2016, 36(6): 1765- 1773.
[20] Ojeda R A, Tabeni S. The mammals of the Monte Desert revisited. Journal of Arid Environments, 2009, 73(2): 173- 181.
[21] 武晓东, 付和平, 杨泽龙. 中国典型半荒漠与荒漠区啮齿动物研究. 北京: 科学出版社, 2009.
[22] 赵肯堂. 三趾跳鼠(DipussagittaPallas)的生态研究. 动物学杂志, 1964, (2): 59- 62.
[23] 甘肃农业大学. 草原保护学-第一分册: 草原啮齿动物学(第二版). 北京: 中国农业出版社, 1997.
[24] 宋恺, 刘荣堂. 子午沙鼠(MerionesmeridianusPallas)的生态研究. 兽类学报, 1984, 4(4): 291- 300.
[25] Eccard J A, Walther R B, Milton S J. How livestock grazing affects vegetation structures and small mammal distribution in the semi-arid Karoo. Journal of Arid Environments, 2000, 46(2): 103- 106.
[26] Jones Z F, Bock C E, Bock J H. Rodent communities in a grazed and ungrazed Arizona grassland, and a model of habitat relationships among rodents in southwestern grass/shrublands. The American Midland Naturalist, 2003, 149(2): 384- 394.
[27] Brown J H, Whitham T G, Ernest S K M, Gehring C A. Complex species interactions and the dynamics of ecological systems: long-term experiments. Science, 2001, 293(5530): 643- 650.
[28] Ernest S K M, Brown J H, Parmenter R R. Rodents, plants, and precipitation: spatial and temporal dynamics of consumers and resources. Oikos, 2000, 88(3): 470- 482.
[29] Arsenault R, Owen-Smith N. Facilitation versus competition in grazing herbivore assemblages. Oikos, 2002, 97(3): 313- 318.
[30] 李俊生, 宋延龄, 曾治高. 7种荒漠啮齿动物食物组成与消化道长度的比较. 动物学报, 2003, 49(2): 171- 178.
[31] Gordon I J, Illius A W, Milne J D. Sources of variation in the foraging efficiency of grazing ruminants. Functional Ecology, 1996, 10(2): 219- 226.
[32] Murray M G, Brown D. Niche separation of grazing ungulates in the Serengeti: an experimental test. Journal of Animal Ecology, 1993, 62(2): 380- 389.
[33] Rosi M I, Puig S, Cona M I, Videla F, Méndez E, Roig V G. Diet of a fossorial rodent (Octodontidae), above-ground food availability, and changes related to cattle grazing in the Central Monte (Argentina). Journal of Arid Environments, 2009, 73(3): 273- 279.
[34] 周宗汉, 刘昌威, 罗远才. 河西荒漠子午沙鼠的生态学初步观察. 四川动物, 1985, (1): 10- 13.
[35] Joseph M B, Preston D L, Johnson P T J. Integrating occupancy models and structural equation models to understand species occurrence. Ecology, 2016, 97(3): 765- 775.
[36] Clough Y. A generalized approach to modeling and estimating indirect effects in ecology. Ecology, 2012, 93(8): 1809- 1815.
[37] Grace J B, Adler P B, Harpole S W, Borer E T, Seabloom E W. Causal networks clarify productivity-richness interrelations, bivariate plots do not. Functional Ecology, 2014, 28(4): 787- 798.
Response of dominant desert rodent species to grazing disturbances: a structural equation modeling analysis
YUAN Shuai1,2, FU Heping1,2, WU Xiaodong1,2,*, Hinggan1,2, GAN Hongjun3,YUE Xiuxian4
1CollegeofGrassland,ResourcesandEnvironment,InnerMongoliaAgriculturalUniversity,Hohhot010011,China2KeyLaboratoryofPratacultureGrasslandResources,MinistryofEducation,Hohhot010011,China3TheGrasslandResearchStationofAlashan,Bayehot750306,China4TheInstituteofForestryMonitoringandPlanningofInnerMongoliaAutonomousRegion,Hohhot010020,China
Livestock grazing is one of the primary causes of disturbances to grassland ecosystems. Ecosystems pay dramatic ecological costs in terms of their composition, structure, and function because of overgrazing. These costs included decreases in plant biomass, reductions in species richness and the number of high quality forage species, and increases in the numbers of toxic species and the percentage of bare ground, as well as changes to the physical and chemical properties of their soil. Livestock grazing has an effect not only on plants and soil, but also on small mammals. Many studies have suggested that grazing affects rodents in terms of their individual conditions, breeding, population dynamics, community structure, and species diversity. These effects could be further complicated by a background of climate change. However, because of the limits of traditional statistical methods, most studies only test individual hypotheses for grazing impacts on rodents, and so few studies test multi-hypothesis frameworks. A study was conducted from 2006 to 2011 on sites experiencing grazing exclusion,rotational-grazing, and over-grazing to investigate the effects of grazing on the population densities of dominant rodent species in Alashan, Inner Mongolia, China. Rodent species were identified and monitored using a live-trapping method.We trapped nine species includingDipussagitta,Stylodipusandrewsi,Allactagasibirica,Phodopusroborovskii,Cricetulusbarabensis,Allocricetuluseversmanni,Merionesmeridianus,M.unguiculatus, andSpermophilusalaschanicus.D.sagitta,A.sibirica, andM.meridianuswere found to be dominant in their local rodent communities. In addition, climate data and soil compaction, as well as the coverage, height, density, and biomass of vegetation were measured in the study areas. In this study, data on dominant rodent species population densities were combined with habitat factors to perform a multi-hypothesis test using structural equation modeling. We expected to find which habitat factors had the greatest effects on the rodent population. The results indicated that climate conditions had direct negative effects on the population densities ofD.sagittaandA.sibirica. The vegetation shield had negative effects on the population density ofA.sibirica, but positive effects on that ofM.meridianus. Increasing soil compaction could indirectly promote population density ofA.sibirica, but directly increased the population density ofM.meridianus. The vegetation shield had the greatest direct effect on dominant rodents, but soil compaction had the most indirect effects. These results suggest that the population dynamics of dominance in desert rodent species have been driven by many processes, and that climate has been a key factor in these processes. However, the population dynamics of dominance in rodent communities has been primarily regulated by changes in grazing due to changing vegetation cover.
rodent; population dynamic; grazing disturbance; structural equation modeling
国家自然科学基金项目(31160096,30560028,30760044);公益性行业(农业)科研专项经费项目(201203041)
2016- 04- 09; 网络出版日期:2017- 03- 02
10.5846/stxb201604090650
*通讯作者Corresponding author.E-mail: wuxiaodong_hgb@163.com
袁帅,付和平,武晓东,兴安,甘红军,岳秀贤.基于结构方程模型分析荒漠啮齿动物优势种对不同放牧干扰的响应.生态学报,2017,37(14):4795- 4806.
Yuan S, Fu H P, Wu X D, Hinggan, Gan H J,Yue X X.Response of dominant desert rodent species to grazing disturbances: a structural equation modeling analysis.Acta Ecologica Sinica,2017,37(14):4795- 4806.
猜你喜欢
疯狂英语·新读写(2024年2期)2024-03-20 01:41:16
海外星云(2021年6期)2021-10-14 07:20:46
意林·少年版(2021年13期)2021-08-30 16:26:41
今日农业(2020年19期)2020-12-14 14:16:52
动漫星空(兴趣百科)(2020年5期)2020-06-10 03:12:02
浙江医学(2018年16期)2018-09-08 05:58:00
消费导刊(2016年7期)2017-08-12 13:23:32
中国科技信息(2015年2期)2015-11-16 08:18:32
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
实验动物与比较医学(2014年5期)2014-02-28 14:53:08
