
外源氮磷负荷比增加对刺苦草(Vallisneria spinu losa)生长的影响∗
2017-07-31 20:02:43郑建伟周茂飞殷春雨关保华
湖泊科学 2017年4期
张 雪,郑建伟,周茂飞,殷春雨,关保华∗∗
(1:南京信息工程大学应用气象学院,南京210044)(2:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京210008)
外源氮磷负荷比增加对刺苦草(Vallisneria spinu losa)生长的影响∗
张 雪1,2,郑建伟1,周茂飞1,2,殷春雨2,关保华2∗∗
(1:南京信息工程大学应用气象学院,南京210044)
(2:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京210008)
磷(P)是水生态系统生产力的限制因子,因此,在水体富营养化治理的过程中,许多地方采取了有效的措施控制流域P的排放;但氮(N)来源复杂,难以有效控制,导致湖泊外源的N∶P负荷比日益扩大.植物的生长需要合适的N∶P比,因此,外源N∶P负荷比的增加可能会对沉水植物的生长产生影响.采取控制实验,将刺苦草(Vallisneria spinulosa)栽种于塑料桶中,每隔3 d添加1次N、P营养盐,实验设置5个处理,P外源性负荷量固定为4mg/(m2·d),N外源性负荷量分别为0、40、80、120、160 mg/(m2·d).实验周期为80 d.结果显示:随着N∶P负荷比的增加,刺苦草的叶干重、植株总干重、根干重、叶N及总N含量、叶片数、叶P及总P含量等指标基本保持不变或略有增加,单株总根长、根茎长度、块茎干重和无性系小株数目则呈现下降趋势.结果表明:随外源N∶P负荷比的增加,刺苦草个体生长指标总体呈现不变的趋势,但与种群扩张潜力相关的指标如根茎长度、块茎干重和无性系小株数目等则呈现下降趋势;刺苦草体内累积的N随外源N∶P负荷比的增加而增加,但P的累积保持不变.说明N∶P负荷比的增加对刺苦草个体生长影响不大,但对种群扩张不利;刺苦草对P的吸收也并不随N∶P比的增加而改变.
沉水植物;氮磷负荷比;生长;形态;种群扩张潜力;刺苦草
Keywords:Submergedmacrophyte;N∶P ratio in loading;growth;morphology;population expansion potential;Vallisneria spinulosa
湖泊富营养化成为日益严峻的环境问题[1].人类活动导致的入湖营养盐增加是湖泊富营养化的主要诱因,大量含有氮(N)、磷(P)等营养物质的工业废水和生活污水排放到湖泊中,雨水冲刷和渗透使残留的化肥也进入湖泊水体,再加上江湖阻断使水流速度大幅减缓,水体滞留时间增长,更新缓慢,水体自净能力减弱[2].湖泊富富营养化的治理已经得到重视,外源污染控制是保障湖泊生态修复的前提[3].目前国内外都重视对外源P的控制,但N的来源比较复杂,难以有效控制,因此湖泊外源的N∶P负荷比日益扩大[4].如鄱阳湖外源负荷中N越来越多,导致N∶P负荷比值持续增加[5].太湖地区无论是来自降雨的湿沉降还是入湖河流带来的外源污染,N含量都远远超过P[6].
土壤中N∶P比的改变会影响植物体内生命物质的合成,从而影响植物的生长速率和死亡率[7-8].沉水植物除了通过根部从沉积物中吸收营养盐外,还能够通过叶片直接从上覆水中吸收营养盐[9].因此,沉水植物的生长不但会受底泥N、P含量的影响,还会受到水体中N、P浓度的影响[9-10].从生态化学计量学特征来看,沉水植物体内的N∶P比低于浮游植物[11],因此,水体中N∶P比增加将不利于沉水植物生长但有利于浮游植物生长[12].湖泊调查和研究发现,沉水植物能够在N、P浓度变化范围较大的湖泊出现,但N∶P比值增加会导致沉水植物生长受到抑制直至消失[13-15].
为了解N∶P负荷比升高对沉水植物生长的影响,本实验参考太湖入湖河流负荷情况,通过每隔3 d添加1次外源营养液,设置P负荷稳定在4 mg/(m2·d),而N负荷设置0、40、80、120和160 mg/(m2·d)梯度,测定刺苦草(Vallisneria spinulosa)的生长和种群扩张与维持潜力相关指标,揭示外源N∶P负荷比增加对沉水植被的影响,为沉水植物保护和水生态修复提供理论支撑.
1 材料与方法
1.1 实验设计及处理
实验于2008年5月17日-8月6日在中国科学院南京地理与湖泊研究所太湖“863”实验基地(31°28′19.19″N,120°13′28.14″E)开展.将20只塑料桶统一放置在一块土壤平台上,桶底端埋入10 cm土,以保持其稳定.塑料桶上内径41 cm、下内径35 cm(下底面积约0.1m2)、深56 cm.每只塑料桶中铺设6 cm沉积物;沉积物于5月17日前采自太湖梅梁湾的芦苇带,拣出沉积物中的螺蛳、杂物和植物残体,混匀后铺入塑料桶中.每只桶中加入40 cm深的水(约40 L)(水来自太湖,泵入湖边的水泥池中澄清1周后使用).沉积物和水放入实验桶中之后,盖上桶盖,遮光静置24 h,避免附着和浮游藻类生长.实验桶放置在梅梁湾岸边的一处开阔平台上,处于自然光照和温度条件.
刺苦草是水鳖科苦草属沉水草本植物,在长江中下游湖泊中广泛分布[16].随着长江中下游湖泊富营养化,刺苦草分布面积逐渐缩小[17].实验开始时,在每只桶中种植10株来源相同、大小一致的刺苦草(干物质含量为0.0605±0.0080 g/株,叶片末端至根末端的长度为16.74±1.90 cm;叶片数8±1),刺苦草为块茎萌发的幼苗.10株刺苦草均匀分配空间.取刺苦草源苗测定初始总氮(TN)、总磷(TP)含量.实验处理如表1所示,每组4个重复.
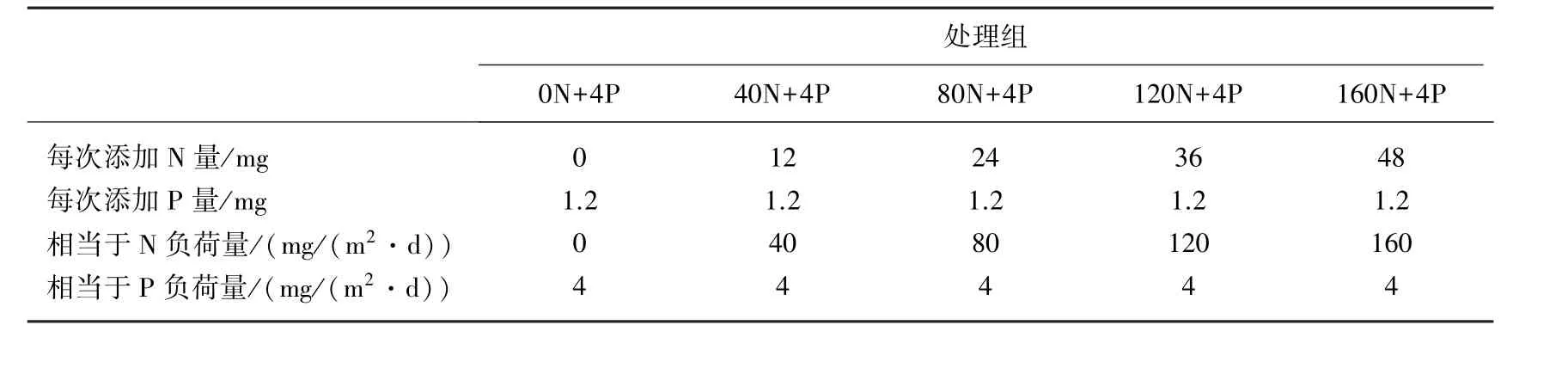
表1 实验处理Tab.1 Experimental treatment
本研究根据太湖等湖泊的外源负荷特征设置入湖N、P负荷量[23-24].实验系统上覆水的氮本底浓度为0.94mg/L,磷本底浓度为0.11mg/L.每3 d添加1次营养盐(NH4Cl与KH2PO4),用N、P添加量除以实验桶底面积(0.1m2)和天数(3 d),获得N、P入水负荷量.实验分5个处理(表1).在添加营养盐之前定时测定水柱pH值和水温.实验过程中每天刷洗桶壁,避免附着生物生长;定期添加蒸馏水补充蒸发掉的水分;下雨时用雨布遮盖,防止过多雨水进入引起溢水.用塑料绳标记每桶中种入的10株刺苦草,以便在实验后期与无性系分株区别开来.
1.2 样品分析
实验结束时,将用塑料绳标记的刺苦草植物完整取出,冲洗干净,数叶片数目;每桶随机取一棵整株,另外随机取一株将根、叶以及根茎和块茎等部分分开,将刺苦草的须根全部拔下,等分成4份,随机取一份测量每根的长度,相加后乘以4,计算每株刺苦草的总根长.整株和各部分分开后的样品放入60℃烘箱中烘干至恒重,测植物整株和各部分干重以及叶片和整株植物的TN、TP含量.收获每桶中所有的刺苦草,测量所有根茎的长度,相加后除以10,获得每桶中单株刺苦草的根茎长.同样的方法获得每桶中单株刺苦草的块茎干重和无性系小株数目.
1.3 数据处理
实验数据运用SPSS 19.0统计软件进行方差分析,采用单因素方差分析,P<0.05为差异性显著.用平均值±标准误在Excel软件中作图.

图1 刺苦草生物量随N∶P负荷比升高的变化Fig.1 Changes of V.spinulosa biomasswith increasing nitrogen and phosphorus ratio loading
2 结果
2.1 刺苦草生物量随N∶P负荷比升高的变化
叶干重、总干重和根干重各处理组间无显著差异(P>0.05).随着N∶P负荷比的增加,叶干重和总干重在变动中略呈增加趋势,根干重的增加趋势比较明显(图1).
2.2 刺苦草形态随N∶P负荷比升高的变化
叶片数各处理间无显著性差异(P>0.05).随着N∶P负荷比的增加,叶片数基本保持不变.单株总根长0N+4P和40N+4P显著高于其他处理(P<0.05),单株总根长随着N∶P负荷比的升高呈现下降趋势(图2).
2.3 刺苦草氮磷含量随N∶P负荷比升高的变化
叶氮含量160N+4P处理组显著高于0N+4P处理组(P<0.05),随着N∶P负荷比的升高,叶氮含量呈现上升趋势;总氮含量160N+4P处理组显著高于其他处理组(P<0.05),总体也呈现上升趋势.叶磷含量和总磷含量各处理间无显著性差异(P>0.05),总体保持不变趋势(图3).
2.4 刺苦草种群扩张潜力随N∶P负荷比升高的变化
根茎长度、块茎干重和无性系小株数各处理间均无显著性差异(P>0.05),随着N∶P负荷比的升高均呈下降趋势(图4).
3 讨论
随着流域P排放措施的实施,汇入湖泊的P含量逐步得到控制,N含量相对较高,外源N∶P负荷比的升高导致许多湖泊的富营养化趋势越来越表现出N∶P比升高的趋势[4-6].如天目湖N∶P比由2001-2002年的18变为2006年的25[18].鄱阳湖、巢湖和太湖水体入湖N∶P负荷比近年也呈现出升高趋势[5,19-20].水生植物体内的N∶P比介于4∶1~13∶1之间,而浮游植物体内的N∶P比值高达16∶1,因此,水体N∶P比提高将会利于浮游植物生长[11-12].在本实验中,经过90 d持续不断的营养盐输入,0N+4P和40N+4P处理组的实验水体N∶P比低于13∶1,而其他外源N∶P负荷比增加的处理组,最终上覆水中N∶P比都超过13∶1(图5).吴红飞等[21]研究发现,N∶P比升高会导致浮游和附着植物生物量增加,但对沉水植物个体生物量没有显著影响.这与本研究得到的结论相似.随外源N∶P负荷比升高,刺苦草的叶干重、叶片数和植株总干重(主要由叶干重组成)等个体指标保持不变甚至略呈增加趋势.
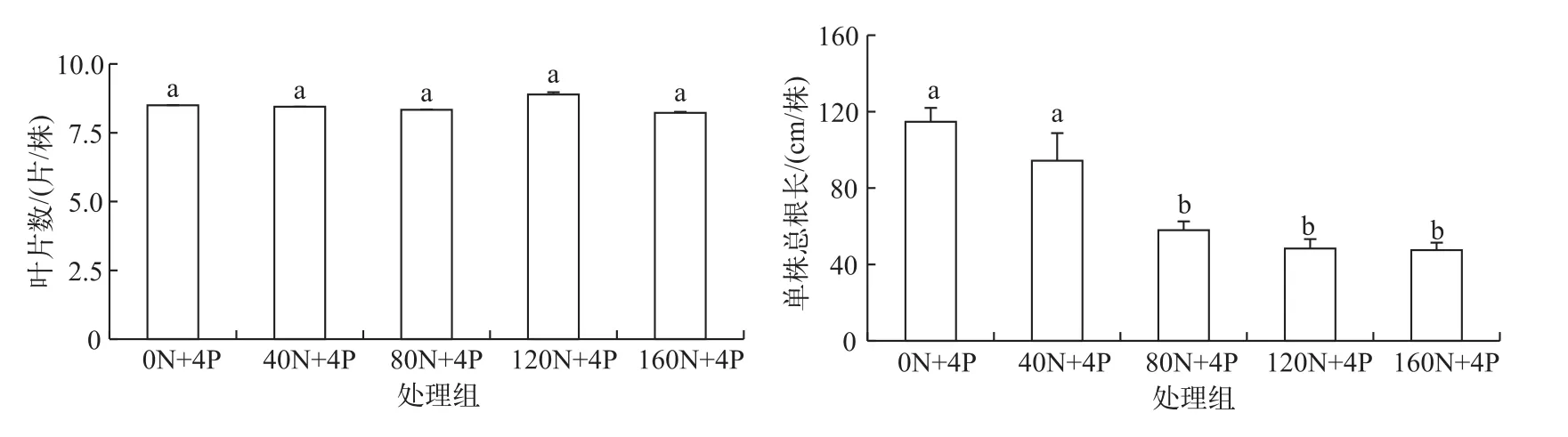
图2 刺苦草形态随N∶P负荷比升高的变化Fig.2 Changes of V.spinulosa morphology with increasing nitrogen and phosphorus ratio loading
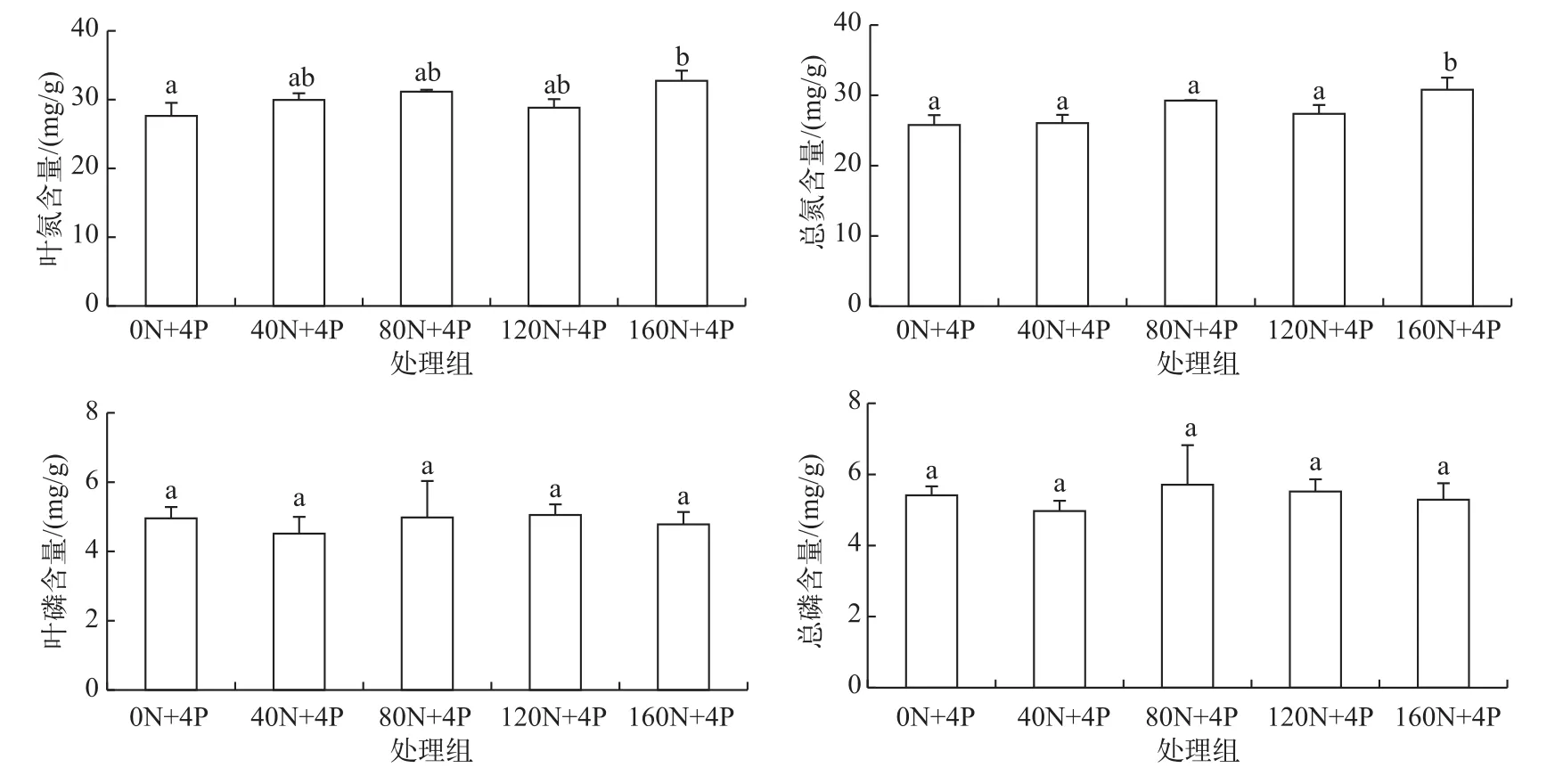
图3 刺苦草氮磷含量随N∶P负荷比升高的变化Fig.3 Changes of nitrogen and phosphorus contents in V.spinulosa with increasing nitrogen and phosphorus ratio loading
但以往的研究对沉水植物繁殖方面的关注较少,因此把水体富营养化导致的沉水植物消失,归因于浮游植物大量生长后对水生植物生长的制约[12].实际上,由于沉水植物体内的N∶P比不超过13∶1,因此,水体N∶P比超过此值可能会不利于沉水植物生长.沉水植物以无性繁殖为主,其无性繁殖器官主要由茎及其衍生物形成[22].一般通过克隆生长,不断增加分枝或分株数目来扩大分布面积,维持种群数量;在生长末期,则形成休眠越冬繁殖体,等生长条件合适时发芽生长[22-23].因此,当沉水植物分枝或分株生长受到制约时,种群扩张能力受到抑制[24];而当休眠体生长受到制约时,未来的种群维持能力受到抑制[25].刺苦草叶基生,无直立茎,生长季节集中在春、夏季,通过根状匍匐茎上分生多数无性系分株拓展分布区域;秋季开花结实,但种子发芽率不高,主要靠越冬块茎在来年发芽维持种群.随着水体N∶P比升高,刺苦草无性系分株数量下降,地下块茎数量和质量(干重)都呈现下降趋势,导致其在长江中下游湖泊的分布面积大量萎缩[16-17].
此外,随着外源N∶P负荷比的升高,刺苦草干物质中P含量不变,但N含量提高,即刺苦草干物质中N∶P比升高.N∶P比提高会导致植物出现P限制,植物倾向于分配较多的碳水化合物到根部去获取充足的P,导致叶片中光合效能相对不足,叶片光合作用受到限制,影响植物的初级生产力,影响植物的种间关系和优势度,导致植物竞争力下降[7-8,26].

图4 刺苦草种群扩张潜力随N∶P负荷比升高的变化Fig.4 Changes of population expansion potential of V.spinulosa with increasing nitrogen and phosphorus ratio loading

图5 各处理组上覆水N∶P比Fig.5 Nitrogen and phosphorus ratio in overlying water in different treatments
沉水植物是维持浅水湖泊清水稳态的关键生物种类,沉水植物覆盖度降低或消失会导致水体从清水稳态向浊水稳态转换[27].因此,在富营养化水体生态修复的过程中,大面积种植沉水植物是一个重要的手段[28].刺苦草是长江中下游水生态修复中常常使用的工具种[27].外源N∶P负荷比的提高会导致湖泊水体N∶P比升高,促进浮游植物生长,抑制沉水植物的种群扩张潜力,不利于沉水植物建群和扩张分布面积;同时也促使沉水植物体内N∶P比提高,影响植物光合作用和碳水化合物流向.因此,控制N负荷、降低N∶P负荷比应该在湖泊保护和生态修复中引起关注.
[1] Qin Boqiang,Yang Liuyan,Chen Feizhou et al.Themechanism and control technology of lake eutrophication and its application.Chinese Science Bulletin,2006,16:1857-1866.[秦伯强,杨柳燕,陈非洲等.湖泊富营养化发生机制与控制技术及其应用.科学通报,2006,16:1857-1866.]
[2] Qin Boqiang,Gao Guang,Zhu Guangwei et al.Lake eutrophication and its ecosystem response.Chinese Science Bulletin,2013,10:855-864.[秦伯强,高光,朱广伟等.湖泊富营养化及其生态系统响应.科学通报,2013,10:855-864.]
[3] Conley DJ,Paerl HW,Howarth RW et al.Controlling eutrophication:Nitrogen and phosphorus.Science,2009,323(5917):1014-1015.
[4] Wu Yali,Xu Hai,Yang Guijun et al.Progress in nitrogen pollution research in Lake Taihu.J Lake Sci,2014,26(1):19-28.DOI:10.18307/2014.0103.[吴雅丽,许海,杨桂军等.太湖水体氮素污染状况研究进展.湖泊科学,2014,26(1):19-28.]
[5] Hu Chunhua.The water environmental characteristic and its evolutionary trends of Poyang Lake[Dissertation].Nanchang:Nanchang University,2010.[胡春华.鄱阳湖水环境特征及演化趋势研究[学位论文].南昌:南昌大学,2010.]
[6] Wang Xiaozhi,Yin Weiqin,Chan Yuhua et al.Nitrogen and phosphorus input from wet deposition in Taihu Lake region:A case study in Changshu ecological Experimental station.Journal ofApplied Ecology,2009,20(10):2487-2492.[王小治,尹微琴,单玉华等.太湖地区湿沉降中氮磷输入量——以常熟生态站为例.应用生态学报,2009,20(10):2487-2492.]
[7] Venterink H O,Güsewell S.Competitive interactions between twomeadow grasses under nitrogen and phosphorus limitation.Functional Ecology,2010,24(4):877-886.
[8] Güsewell S.N∶P ratios in terrestrial plants:Variation and functional significance.New phytologist,2004,164(2):243-266.
[9] Denny P.Sites of nutrient absorption in aquatic macrophytes.The Journal ofEcology,1972,60(3):819-829.
[10] Wang Bin,LiWei.Physiological reactionsof Potamogetonmalaianus to different N and P concentrations in the growthmedium.Journal of Ecology,2002,10:1616-1621.[王斌,李伟.不同N、P浓度条件下竹叶眼子菜的生理反应.生态学报,2002,10:1616-1621.]
[11] Xing Wei,Liu Han,Liu Guihua.Ecological stoichiometry in aquatic ecosystems:Studies and applications.Journal of Plant Science,2015,(5):608-619.[邢伟,刘寒,刘贵华.生态化学计量学在水生态系统中的研究与应用.植物科学学报,2015,(5):608-619.]
[12] Sagrario G,María A,Jeppesen E et al.Does high nitrogen loading prevent clear-water conditions in shallow lakesatmoderately high phosphorus concentrations?Freshwater Biology,2005,50(1):27-41.
[13] Hu Mianhao,Yuan Juhong,Xiang Lücheng et al.Influence of different nitrogen:phosphorus on growth characteristic of perennial aquatic plant.Journal of Environmental Engineering,2011,5(11):2487-2493.[胡绵好,袁菊红,向律成等.不同氮磷比对多年生水生植物生长特性影响的研究.环境工程学报,2011,5(11):2487-2493.]
[14] James C,Fisher J,Moss B.Nitrogen driven lakes:The Shropshire and Cheshire Meres?Archiv für Hydrobiologie,2003,158(2):249-266.
[15] Jensen JP,Kristensen P,Jeppesen E.Relationships between nitrogen loading and in-lake concentrations in shallow Danish lakes.International Association of Theoretical and Applied Limnology,Proceedings,1990.
[16] Chen Lei.Population dynamics and persistence of Vallisneria spinulosa in lakes of themiddle-lower reaches of the Yangtze River revealed by molecular markers[Dissertation].Wuhan:Wuhan Botanical Garden,Chinese Academy of Sciences,2007.[陈磊.基于分子标记的刺苦草在长江中下游湖泊的种群动态与维持研究[学位论文].武汉:中国科学院武汉植物园,2007.]
[17] Meng Fanjuan.Effects of eutrofication and changing water level on biomass of submerged macrophytes[Dissertation]. Hefei:China University of Science and Technology,2011.[孟凡娟.湖泊富营养化和水位变化对沉水植被生物量的影响[学位论文].合肥:中国科学技术大学,2011.]
[18] Gao Yongxia,Zhu Guangwei,He Ranran et al.Variation ofwater quality and trophic state of Lake Tianmu.EnvironmentalScience,2009,(3):673-679.[高永霞,朱广伟,贺冉冉等.天目湖水质演变及富营养化状况研究.环境科学,2009,(3):673-679.]
[19] Wang Shunli,Zhang Hongcai,Wei Hefen et al.Eutrophication current situation and research of the east-half of Chaohu Lake.Journal of Biology,2016,(2):13-17.[汪顺丽,张宏才,魏荷芬等.巢湖东半湖富营养化现状的研究.生物学杂志,2016,(2):13-17.
[20] Zhu Guangwei.Eutrophic statusand causing factors for a large,shallow and subtropical Lake Taihu.JLake Sci,2008,20(1):21-26.DOI:10.18307/2008.0103.[朱广伟.太湖富营养化现状及原因分析.湖泊科学,2008,20(1):21-26.]
[21] Wu Hongfei,Guan Baohua,Yao Sipeng etal.Influence ofN:P ratio on primary producersofshallow lakes.Ecological Science,2014,33(6):1041-1046.[吴红飞,关保华,姚思鹏等.氮磷负荷比对浅水湖泊初级生产者的影响.生态科学,2014,33(6):1041-1046.]
[22] Yuan Longyi,Xue Xinghua,Jing Bohan.Advances in the growth pattern and reproduction characteristics of aquatic plants. Northern Gardening,2013,24:191-195.[袁龙义,薛兴华,经博翰.水生植物生长格局和繁殖特点研究进展.北方园艺,2013,24:191-195.]
[23] Grace JB.The adaptive significance of clonal reproduction in angiosperms:An aquatic perspective.Aquatic Botany,1993,44(2/3):159-180.
[24] Wang Wei.Characteristics of reproduction and spread for Aconitum leucostomum Worosch population in Nalati Mountain Meadow[Dissertation].Urumchi:Xinjiang Agricultural University,2012.[王卫.那拉提山地草甸草地白喉乌头种群繁殖及蔓延特性研究[学位论文].乌鲁木齐:新疆农业大学,2012.]
[25] Jiao Dezhi,Me Lu,Huang Zhaoyue et al.Bud population dynamics of Phragmites australis in heterogeneous habitats of Northeast grassland.Journal ofApplied Ecology,2015,26(2):404-410.[焦德志,么璐,黄曌月等.东北草地异质生境芦苇芽种群动态.应用生态学报,2015,26(2):404-410.]
[26] Wang Qi,Xu Chengyang.Affects of nitrogen and phosphorus on plant leaf photosynthesis and carbon partitioning.Journal of Shandong Forestry Science and Technology,2005,(5):59-62.[王琪,徐程扬.氮磷对植物光合作用及碳分配的影响.山东林业科技,2005,(5):59-62.]
[27] Liu Zhengwen.Ecosystem recovery and water quality improvement of lakes.China Water Conservancy,2006,17:30-33.[刘正文.湖泊生态系统恢复与水质改善.中国水利,2006,17:30-33.]
[28] Liu Congyu,Liu Pingping,Liu Zhengwen et al.Study on the functions of submerged macrophytes in ecological restoration and water quality improvement.Anhui Agricultural Sciences,2008,36(7):2908-2910.[刘从玉,刘平平,刘正文等.沉水植物在生态修复和水质改善中的作用——以惠州南湖生态系统的修复与构建(中试)工程为例.安徽农业科学,2008,36(7):2908-2910.]
Effect of increasing nitrogen/phosphorus ratio in loading on the grow th of Vallisneria spinulosa
ZHANG Xue1,2,ZHENG Jianwei1,ZHOU Maofei1,2,YIN Chunyu2&GUAN Baohua2∗∗
(1:College of Applied Meteorology,Nanjing University of Information Science&Technology,Nanjing 210044,P.R.China)
(2:State Key Laboratory of Lake Science and Environment,Nanjing Institute ofGeography and Limnology,Chinese Academy ofSciences,Nanjing 210008,P.R.China)
Phosphorus(P)is a limiting factor for productivity in aquatic ecosystems.In order to control lake eutrophication,many effectivemeasures are taken to control the P effluent in the basin,but nitrogen(N)is difficult to control due to its complex sources.Thus,the situation of low P but yethigh N effluent results in a high ratio of N∶P in loading inmany lakes.Plantsneed a suitable N∶P ratio to sustain their growth,therefore,the increasing of the N∶P ratio in effluentsmight affect the growth of submerged macrophytes.To acquire an insight into the influence of increasing N∶P ratios on the submerged plants,we carried out an outdoor control experiment.Vallisneria spinulosa was planted in plastic bucketswith N and P added to the bucketsevery 3 days.We set up five treatments with an unchanging P loading dose of 4 mg/(m2·d),with a range of nitrogen loading dose including 0,40,80,120 and 160 mg/(m2·d).The experiment lasted for 80 days.Results showed thatwith an increase of the N∶P ratio in the loading,some indicatorswere nearly stable,such as leaf dry weight,total plant dry weight,leaf N and total N content,leaf number,leaf P and total P content,while some other indicators,such as plant total root length,cloning stem length,stem dry weight and ramet numbers decreaed.The results above revealed thatwith an increasing N∶P ratio in loading,individual growth indicators of V.spinulosa would remain stable or exhibit a minor increase,but that the indicators of population expansion ability,such as cloning stem length,tuber dry weightand rametnumbers,would decline,while the P absorption would not changewith increasing N∶P ratio in loading.
DOI 10.18307/2017.0411
©2017 by Journal of Lake Sciences
∗国家自然科学基金项目(31270409)资助.2016-05-18收稿;2016-10-07收修改稿.张雪(1992~),女,硕士研究生;E-mail:evonneshary@163.com.
∗∗通信作者;E-mail:bhguan@niglas.ac.cn.
猜你喜欢
海洋通报(2022年4期)2022-10-10 07:41:48
舰船科学技术(2022年11期)2022-07-15 07:51:56
皮革制作与环保科技(2020年14期)2020-03-17 07:16:04
花卉(2020年12期)2020-01-09 07:12:52
西藏农业科技(2019年3期)2019-11-04 00:35:10
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21 07:43:18
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
环境保护与循环经济(2017年10期)2017-03-16 03:16:20
大科技(2016年1期)2016-07-17 02:42:07
