
UGT1A10基因SNP rs10187694对吗替麦考酚酯代谢的影响*
2017-07-18 11:50肖超孙红成邓贵龙宋国贺王宇鹏陈健孙星钟林彭志海王晓亮
中国现代医学杂志 2017年12期
肖超,孙红成,邓贵龙,宋国贺,王宇鹏,陈健,孙星,钟林,彭志海,王晓亮
(上海交通大学附属第一人民医院 普外科,上海 200080)
UGT1A10基因SNP rs10187694对吗替麦考酚酯代谢的影响*
肖超,孙红成,邓贵龙,宋国贺,王宇鹏,陈健,孙星,钟林,彭志海,王晓亮
(上海交通大学附属第一人民医院 普外科,上海 200080)
目的 体外研究尿苷二磷酸葡萄糖醛酸转移酶(UGT1A10)基因SNP rs10187694多态位点G/A变异对吗替麦考酚酯(MMF)代谢的影响,明确该位点变异是否与MMF个体代谢差异有关。方法采用基因重组、定点突变技术,构建UGT1A10基因SNP rs10187694位点含有不同等位基因的重组过表达载体,POLO 3000转染法将重组过表达载体转染入HEK293细胞,采用液相色谱-质谱联用仪(LC/MS/MS)系统,检测霉酚酸(MPA)代谢产物7-O-葡醛酸苷(MPAG)的生成量,以评估转染不同等位基因HEK293细胞中酶的活性。结果成功构建pIRES2-EGFP-prom(G)和pIRES2-EGFP-prom(A)重组过表达载体,并转染HEK293细胞。LC/MS/MS系统检测结果显示,携带A等位基因的突变型UGT1A10代谢MMF产生MPAG的能力降低,其24 h MPAG的生成量为(226.00±14.57)nmol/L,而野生型的生成量为(269.00±14.07)nmol/L,UGT1A10突变型 MPAG 的生成量是野生型的84.00%,两者比较,差异有统计学意义(P<0.05)。结论UGT1A10基因SNP rs10187694位点G/A突变可影响MMF代谢产物MPAG的生成,可能是造成不同个体MMF代谢差异的重要原因之一。
尿苷二磷酸葡萄糖醛酸转移酶;霉酚酸酯;药物代谢
吗替麦考酚酯(mycophenolate mofetil,MMF)是临床广泛应用的抗增殖免疫抑制剂之一,能够预防肝、肾等大脏器移植术后的急、慢性移植排斥反应[1]。MMF作为霉酚酸(mycophenolic acid,MPA)的2-乙基酯类衍生物,是一种前体药物,本身不具有活性,经口服或静脉给药后,需在体内迅速水解生成活性产物MPA,才能发挥药理作用[2]。MPA能非竞争性、选择性及可逆性地抑制次黄嘌呤核苷酸酶(inosine-5'-monophosphate dehydrogenase,IMPDH),从而抑制T、B淋巴细胞增殖,被广泛用于器官移植术后和自身免疫病的治疗。但是MPA的个体化药代动力学差异明显,如果免疫抑制不足就会发生器官移植后排斥反应,免疫抑制过度则会产生许多副作用,比如腹泻、贫血、白细胞减少和感染等[3]。尿苷二磷酸葡萄糖醛酸转移酶(UDP-glucuronosyltransferases,UGT)是参与MPA代谢的限速酶,UGT及其单核苷酸多态性(single nucleotide polymorphism,SNP)会影响MPA的药物代谢,并最终造成不同人群、不同个体免疫抑制的疗效和副作用出现差异[4]。
UGT是一个超家族酶,其中最重要的是UGT1和UGT2家族。有研究表明,UGT1亚家族中的UGT1A10参与MMF代谢,UGT1A10基因多态性可以影响MMF代谢产物7-O-葡醛酸苷(mycophenolic acid glucuronide,MPAG)的生成水平[5]。UGT1A10 基因SNP rs10187694是第415位碱基处的G碱基突变为A碱基。该位点突变后,对MMF代谢影响尚不明确。因此,本研究通过体外构建UGT1A10基因该位点的野生型和突变型过表达载体,评估转染不同等位基因的细胞中MMF代谢产物MPAG的24 h生成量,从而明确该位点突变对MMF代谢的影响,将有助于预测用药时可能产生的副作用,并为临床个体化用药提供指导意义。
1 材料与方法
1.1 材料与试剂
MPA、MPAG购自美国Roche Bioscience公司,HEK293细胞(美国ATCC公司),DH5α感受态细胞(北京全式金公司),BglⅡ酶、SalⅠ酶购自美国MBI公司,DNA连接试剂盒(日本TaKaRa公司),小量质粒抽提试剂盒、聚合酶链反应(polymerase chain reaction,PCR)产物纯化试剂盒购自美国Axgene公司,胎牛血清、改良伊格尔培养基(dulbecco's modified eagle medium,DMEM)基础培养基购自美国Invtrogen公司,POLO 3000(上海锐赛生物技术有限公司),质粒pIRES2-EGFP、引物设计及目的基因测序为美国Invitrogen公司。
1.2 仪器与设备
PCR扩增仪(美国ABI公司),CEQ8000测序仪(美国Beckman&Coulter公司)。液相色谱-质谱联用仪(LC/MS/MS)系统包括:Agilent 1100(美国安捷伦科技有限公司)液相色谱系统、LEAP CTC HTS PAL自动进样系统(瑞士CTC分析仪器股份公司)、API4000三重四级杆检测器(美国应用生物系统有限公司),操作软件为Analyst 1.5.1。
1.3 方法
1.3.1 UGT1A10基因CDS区全长扩增和过表达载体的构建 以含目的基因UGT1A10的质粒作为模板,PCR扩增UGT1A10基因CDS区,在目的基因的5'-端添加BglⅡ酶切位点,3'-端添加SalⅠ酶切位点,引物设计信息见表1。反应条件:95℃预变性5 min,95℃变性 30 s,61℃退火 30 s,72℃延伸 60 s,共35个循环,72℃继续延伸10 min。预期PCR扩增产物全长1 593 bp,经琼脂糖凝胶电泳分离PCR产物,观察目的DNA条带并拍照。目的基因PCR产物和目的载体pIRES2-EGFP用BglⅡ、SalⅠ分别进行酶切后,将目的基因连接至表达载体。将10μl连接产物转化DH5α感受态细胞,涂布于含卡那抗性的溶菌肉汤(luria bertani,LB)培养皿,37℃培养过夜。次日挑取单菌落进行菌落PCR,将PCR鉴定呈阳性的克隆接种到LB液体培养基中,摇菌过夜,次日抽提质粒。将菌落PCR呈阳性的质粒进行酶切鉴定,预期DNA产物应为1 593 bp和5.3 kbp 2个条带,将菌落PCR和酶切鉴定均呈阳性的克隆进行测序和序列比对。
1.3.2 UGT1A10基因相应SNP位点定点突变和过表达载体的构建 按照突变PCR程序,进行分段扩增并电泳切胶回收2个片段。将上述2个片段进行PCR拼接,进行突变体全长扩增。用BglⅡ/SalⅠ酶对突变体PCR产物和目的载体pIRES2-EGFP进行双酶切,并将酶切回收的突变PCR产物与pIRES2-EGFP连接。按1.3.1的方法,培养挑选菌落进行PCR和酶切鉴定,预期酶切鉴定DNA产物应为1 590和2 300 bp 2个条带,将鉴定均呈阳性的克隆测序与序列比对。见表1。
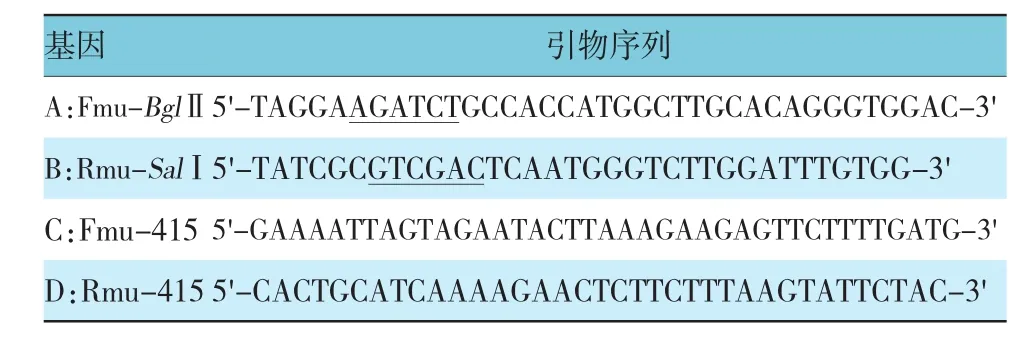
表1 扩增与突变引物序列
1.3.3 UGT1A10目的基因转染HEK293细胞 采用POLO 3000转染法,将携带绿色荧光蛋白的目的基因真核过表达载体pIRES-EGFP-UGT1A10转染HEK293细胞。于转染前24 h,将HEK293细胞用胰蛋白酶充分消化,接入6孔板,然后将各孔细胞分别置于1 ml(DMEM+10%胎牛血清+1%青霉素和链霉素+1%Glutamax)培养基,37℃、95%湿润空气、5%二氧化碳CO2培养箱中,使转染前细胞融合度达60%~80%。转染当天,将6孔板各孔分别换上新鲜800 μl DMEM+10%胎牛血清的培养基(不含抗生素),37℃孵育。配制转染所用的试剂混合物。A管取目的基因真核表达载体、阳性对照质粒、阴性对照质粒各 1.2 μg,分别与 50 μl Opti-MEM 混匀;B 管取19.2 μl Lipo 2000与300 μl Opti-MEM混匀(6孔用量)。室温静置5 min,分别取50 μl B管溶液加入A组各管中,混匀,室温孵育20 min。取100 μl上述DNA-Lipo混合液,均匀滴加到6孔板的相应HEK293细胞中,轻轻摇匀,37℃培养箱过夜。8 h后,换新鲜的完全培养基继续培养。转染48 h后,荧光显微镜观察细胞的转染效率并拍照。
1.3.4 转染不同过表达载体HEK293细胞的MPAG产量检测 MMF的有效成分是MPA,MPA的主要终产物是MPAG,故可以将MPA作为底物加入HEK293细胞培养皿,通过评价最终代谢产物MPAG的水平来评估不同基因型细胞对MMF的代谢能力。用二甲基亚砜(dimethyl sulfoxide,DMSO)将MPA配制为浓度为600 mmol/L的溶液,用DMEM Media稀释DMSO终浓度为0.1%,MPA的终浓度为600 μmol/L。将稀释好的MPA加入转染UGT1A10野生型、突变型及空白质粒的HEK293细胞中,在37℃、5%CO2条件下培养2、4和24 h后,加入 4倍含内标的预冷甲醇,终止反应。离心取上清液,LC/MS/MS系统检测代谢终产物MPAG的生成量。最终结果以突变型UGT1A10的相对活性表示:相对活性(%)=突变型亚酶活性/野生型亚酶活性×100%。
1.4 统计学方法
数据分析采用SPSS 13.0统计软件,计量资料以均数±标准差(±s)表示,用 t检验,P<0.05为差异有统计学意义。
2 结果
2.1 UGT1A10基因CDS区全长扩增和过表达载体的构建结果
PCR扩增产物经过琼脂糖凝胶电泳,得到与预期1 593 bp大小一致的目的条带(见图1)。样品3~10在1 593 bp处均见与预期一致的目的条带(见图2)。随机挑选的样品,均在1 593 bp和5.3 kbp处出现与预期一致的2条目的条带(见图3)。
2.2 UGT1A10基因定点突变和过表达载体的构建结果
样品4~9均在15 90 bp处出现目的条带(见图4)。样品1、2、3均在1 590和2 300 bp处出现与预期一致的2条目的条带。见图5。

图1 UGT1A10目的基因的PCR扩增结果
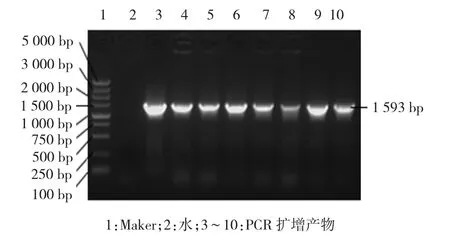
图2 重组克隆菌落的PCR扩增结果
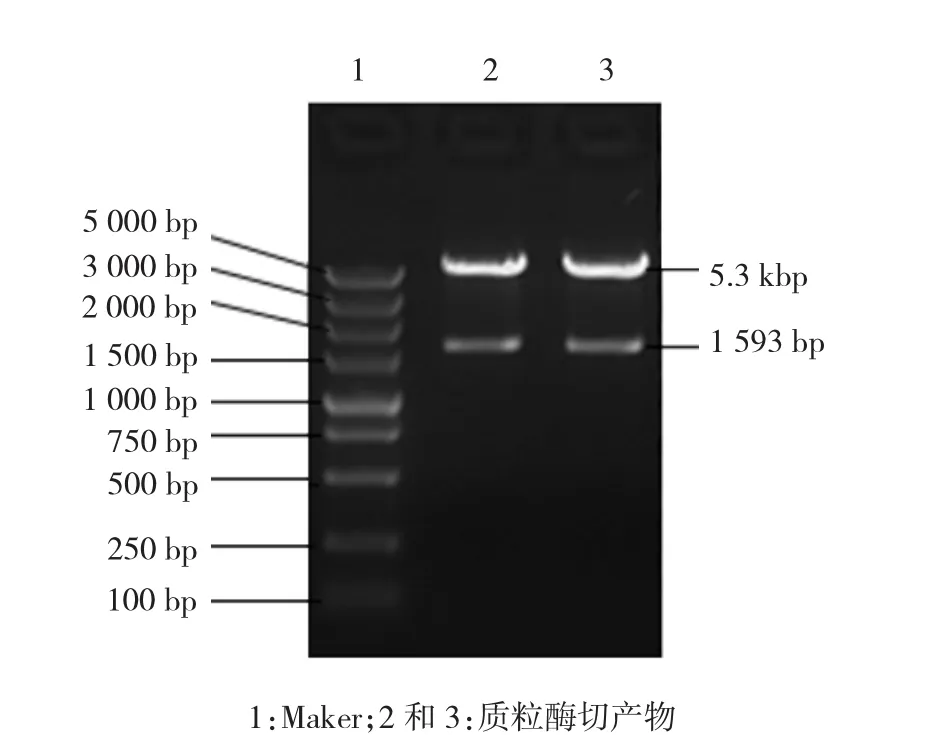
图3 重组UGT1A10质粒酶切产物经琼脂糖凝胶电泳图
2.3 目的基因定点突变前后测序结果
连接成功的样品进行基因测序证明,目的基因的相应位点(UGT1A10基因的第415位碱基)的G碱基已经突变为A碱基,并且其他位置的基因序列与基因库中该基因序列无差异。见图6。
2.4 目的基因过表达载体转染细胞结果
分别将空白质粒、携带突变型位点及野生型位点的UGT1A10过表达载体(表达绿色荧光蛋白)转染HEK293细胞。48 h后,普通光学显微镜和荧光显微镜下转染细胞结果。见图7。
2.5 转染不同过表达载体HEK293细胞的MPAG生成量比较
分别转染突变型、野生型及空白质粒的3种HEK293细胞,每种细胞设置6个复孔,共3个6孔板,以检测终产物MPAG的生成量,最终每种细胞MPAG的生成量为该种细胞各复孔结果的平均值。突变型与野生型复孔生成量比较,经t检验,差异有统计学意义(t=5.199,P=0.000)。将相同浓度的底物MPA加入上述细胞24 h后,分别检测转染突变型、野生型期空白质粒的HEK293细胞所代谢产生的MPAG生成量,结果显示,在转染空白质粒的HEK293细胞中,6个复孔均未检测到MPAG;在转染突变型UGT1A10(AA)的HEK293细胞中,检测到的MPAG生成量为转染野生型UGT1A10(GG)的84.00%。结果表明,该位点的基因突变会影响MPA代谢产物MPAG的生成。2种UGT1A10类型各复孔MPAG的生成量见表2。

图4 重组克隆菌落的PCR扩增结果

图5 重组UGT1A10质粒酶切产物经琼脂凝胶电泳图

图6 目的基因定点突变前后测序结果
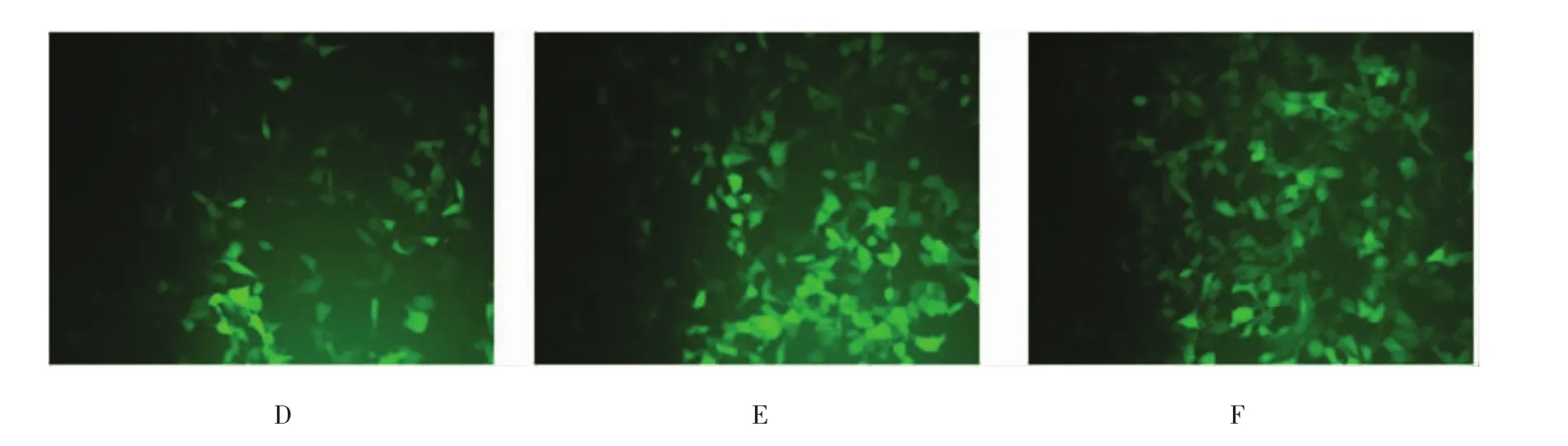
图7 重组质粒转染细胞结果 (×40)
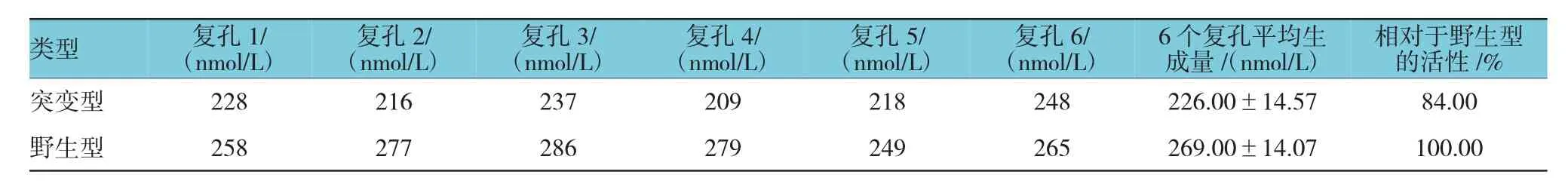
表2 2种UGT1A10类型各复孔MPAG的生成量比较
3 讨论
目前,MMF被广泛应用于预防器官移植术后的急、慢性排斥反应和自身免疫疾病的治疗[6]。MMF口服完全吸收,在体内可迅速且完全(95%)地转化成活性成分MPA,进而发挥免疫抑制作用。然而,MMF的治疗窗较窄,许多研究报道其药效和毒性反应,且存在很大个体差异[7-8]。有研究表明,体外孵育实验时,相同浓度MPA对T、B淋巴细胞产生的抑制效果差异不大。然而,口服相同剂量MMF时,不同个体的MPA暴露水平差异>10倍[9]。临床许多因素,如性别、年龄、吸烟、肝肾功能及白蛋白浓度等,均可影响MPA的药物代谢。UGT作为MPA代谢的限速酶,其SNP也被证明是MPA在不同个体之间产生药物代谢差异的重要原因之一。
UGT是酶的超家族,含有117种酶,根据其核苷酸序列的相似性,分为4大家族:UGT1、UGT2、UGT3及UGT8。其中,最重要的是UGT1和UGT2家族。UGT1又包括 9个亚型:UGT1A(1、3~10),UGT2 包括亚型:UGT2A(1~3)和 UGT2B(4、7、10、11、15、17、28)[10]。人体内 40%~70%的药物需要通过Ⅱ相代谢反应清除,而由UGT催化的葡萄糖醛酸化反应占人体所有Ⅱ相反应>35%[11]。MMF的活性成分是MPA,MPA也需要在肝脏、肠道、肾脏的UGT的作用下,经过葡萄糖醛酸化反应,最终生成主产物MPAG,并经肾脏排出体外[3]。在胃肠道中,UGT1A10对MPA葡萄苷酸化具有重要作用。已有研究表明,UGT1A10基因的多态性影响多种药物的代谢,如某些抗肿瘤药、免疫抑制类药物,如麦考酚酸、类固醇类和酚类物质等。基因多态性方面,SAEKI等[12]发现,UGT1A10的 3 个单核苷酸多态性:A177G(M59I)、T605C(T202I)和 T693C(无氨基酸改变)。其中,T202I的突变可能会对口服药物的肠肝循环和葡萄苷酸化产生影响[13]。MARTINEAU等[14]报道UGT1A10的I211T突变会导致该酶对麦考酚酸的代谢能力丧失。ELAHI等[15]发现UGT1A10的E139K、T240M和L244I突变会导致氨基酸的改变,E139K突变即UGT1A10基因SNP rs10187694位点G/A突变。
如上所述,许多研究表明,UGT1A10对MPA的代谢产生影响。为此,本研究采用体外实验,构建UGT1A10基因SNP rs10187694位点野生型和突变型过表达载体,通过评估转染该位点不同等位基因细胞中MMF代谢产物MPAG的24h生成量,明确该位点突变对MMF代谢产生的影响。数据分析显示,携带A等位基因的变异型UGT1A10酶的活性降低,代谢产物MPAG的24 h生成量为携带G等位基因野生型的84.00%。结果表明,UGT1A10基因SNP rs10187694位点突变可影响MMF代谢产物MPAG的24 h生成量,推测该位点的多态性可能是造成不同个体MMF药物代谢差异的重要原因之一。
在本研究中,笔者揭示UGT1A10基因第415位点突变与MPAG生成量的关系,表明UGT1A10基因多态性可能是造成不同个体MMF药物代谢差异的重要原因之一。当然,关于UGT1A10还有许多方面需要深入探索,如监测临床患者在服用MMF后的血药浓度变化、进一步明确该酶参与体内药物转化的机制和过程,以及研究我国不同地区、人群中该基因的变异率等。
总之,UGT1A10基因多态性是影响MMF代谢的重要因素之一,本研究为临床MMF的个体化用药提供理论支持。如果能够根据患者的基因遗传特征,预测血药浓度,合理设计个体化的给药方案,相信会取得更好的治疗效果。
[1]WAGNER M,EARLEY A K,WEBSTER A C,et al.Mycophenolic acid versus azathioprine as primary immunosuppression for kidney transplantrecipients[J].Cochrane Database SystRev,2015,12:DOI:10.1002/14651858.CD007746.
[2]SOKUMBI O,EL-AZHARY R A,LANGMAN L J.Therapeutic dose monitoring of mycophenolate mofetil in dermatologic diseases[J].J Am Acad Dermatol,2013,68(1):36-40.
[3]STAATZ C E,TETT S E.Pharmacology and toxicology of mycophenolate in organ transplant recipients:an update[J].Arch Toxicol,2014,88(7):1351-1389.
[4]DUPUIS R,YUEN A,INNOCENTI F.The influence of UGT polymorphisms as biomarkers in solid organ transplantation[J].Clinica Chimica Acta,2012,413(17-18):1318-1325.
[5]THERVET E,ANGLICHEAU D,LEGENDRE C.Pharmacology of mycophenolate mofetil:recent data and clinical consequences[J].Nephrologie,2001,22(7):331-337.
[6]BATKO B,KRAWIEC P,OSIELENIEC J,et al.Mycophenolate mofetil in the treatment of selected connective tissue diseases[J].Przegl Lek,2013,70(9):724-729.
[7]SAINT-MARCOUX F,VANDIERDONCK S,PREMAUD A,et al.Large scale analysis of routine dose adjustments of mycophenolate mofetil based on global exposure in renal transplant patients[J].Ther Drug Monit,2011,33(3):285-294.
[8]KIM H,YI N J,LEE J,et al.Safety of reduced dose of mycophenolate mofetil combined with tacrolimus in living-donor liver transplantation[J].Clin Mol Hepatol,2014,20(3):291-299.
[9]SHAW L M,KORECKA M,VENKATARAMANAN R,et al.Mycophenolic acid pharmacodynamics and pharmacokinetics provide a basis for rational monitoring strategies[J].Am J Transplant,2003,3(5):534-542.
[10]MACKENZIE P I,BOCK K W,BURCHELL B,et al.Nomenclature update for the mammalian UDP glycosyltransferase(UGT)gene superfamily[J].Pharmacogenet Genomics,2005,15(10):677-685.
[11]GUILLEMETTE C,LEVESQUE E,ROULEAU M.Pharmacogenomics of human uridine diphospho-glucuronosyltransferases and clinical implications[J].Clin Pharmacol Ther,2014,96(3):324-339.
[12]SAEKI M,OZAWA S,SAITO Y,et al.Three novel single nucleotide polymorphisms in UGT1A10[J].Drug Metab Pharmacokinet,2002,17(5):488-490.
[13]JINNO H,SAEKI M,TANAKA-KAGAWA T,et al.Functional characterization of wild-type and variant(T202I and M59I)human UDP-glucuronosyltransferase 1A10[J].Drug Metab Dispos,2003,31(5):528-532.
[14]MARTINEAU I,TCHERNOF A,BELANGER A.Amino acid residue ILE211 is essential for the enzymatic activity of human UDP-glucuronosyltransferase 1A10(UGT1A10)[J].Drug Metabolism And Disposition,2004,32(4):455-459.
[15]ELAHIA,BENDALY J,ZHENG Z,etal.Detectionof UGT1A10 polymorphisms and their association with orolaryngeal carcinoma risk[J].Cancer,2003,98(4):872-880.
(童颖丹 编辑)
Effect of SNP rs10187694 ofUGT1A10gene on metabolism of Mycophenolate mofetil*
Chao Xiao,Hong-cheng Sun,Gui-long Deng,Guo-he Song,Yu-peng Wang,Jian Chen,Xing Sun,Lin Zhong,Zhi-hai Peng,Xiao-liang Wang
(Department of General Surgery,Shanghai General Hospital,Shanghai 200080,China)
ObjectiveTo study the effect of G/A mutation in SNP rs10187694 site of uridine diphosphate glucuronosyl transferase (UGT1A10)on Mycophenolate mofetil metabolism.MethodsThe recombinant overexpression vectors ofUGT1A10gene were constructed by gene recombination technique and the strategy of site directed mutagenesis.Then,the recombinant plasmid was transfected into HEK293 cells by POLO3000 method.By detecting the yields of mycophenolic acid(MPA)metabolite 7-o-glucuronide(MPAG),the activity of enzyme in the HEK293 cells transfected with different alleles were evaluated.ResultsThe overexperession vectors pIRES2-EGFP-prom(G)and pIRES2-EGFP-prom(A)were successfully constructed and transfected into HEK293 cells.The results analyzed by LC/MS/MS showed that the cells containing mutant type ofUGT1A10gene with A allele produced less MPAG than those with G allele,the concentration of MPAG produced by mutant type in 24 hours was(226.00±14.57)nmol/L,however,the concentration in wild type was(269.00±14.07)nmol/L.The generation amount of MPAG in mutant type was 84.00%of that in wild type in 24 hours(P<0.05).ConclusionsUGT1A10gene SNP rs10187694 can significantly affect the production of MPAG,which may be one of the reasons causing the difference in Mycophenolate mofetil metabolism.
UGT1A10;Mycophenolate mofetil;drug metabolism
R969.1
A
10.3969/j.issn.1005-8982.2017.12.002
1005-8982(2017)12-0009-06
2016-03-29
国家自然科学基金(No:81270557,81110108010)
王晓亮,E-mail:xiaoliangwang2009@163.com
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
世界科学技术-中医药现代化(2022年3期)2022-08-22
成都医学院学报(2022年4期)2022-08-19
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
江西农业学报(2021年4期)2021-04-20
中国产前诊断杂志(电子版)(2020年4期)2021-01-11
中国医学物理学杂志(2020年11期)2020-12-12
三农资讯半月报(2020年11期)2020-06-21
