
红景天苷对急性缺氧性神经细胞损伤保护作用及机制探讨
2017-06-28 16:06:44蒲亚芸
陕西医学杂志 2017年6期
王 军,曹 艳,常 江,蒲亚芸
武警陕西省总队医院麻醉科(西安710054)
红景天苷对急性缺氧性神经细胞损伤保护作用及机制探讨
王 军,曹 艳,常 江,蒲亚芸
武警陕西省总队医院麻醉科(西安710054)
目的:探讨红景天苷对急性缺氧性神经细胞损伤的保护作用及机制。方法:新生小鼠处死后取大脑皮层神经元进行原代细胞培养,神经元细胞经红景天苷处理后测定胞内ATP水平、己糖激酶等活性,提取mRNA通过RT-PCR检测细胞内PI3K、己糖激酶、葡萄糖转运体-1 mRNA表达水平。同时以1%低氧处理原代神经元细胞建立急性缺氧模型,红景天苷处理后,通过MTT法观察红景天苷对缺氧神经元细胞的保护作用,PI3K抑制剂LY294002处理缺氧神经元细胞后探讨红景天苷的分子保护机制。结果:红景天苷组处理细胞的PI3K、己糖激酶(HK)、葡萄糖转运体-1(GLUT-1)等表达水平显著升高, HK活性增强,胞内ATP水平显著提高。PI3K抑制后红景天苷的上述作用明显减弱。缺氧处理促进神经细胞死亡,红景天苷减少缺氧性神经细胞死亡,PI3K抑制后红景天苷的上述作用明显减弱。结论:红景天苷处理可诱导神经细胞中PI3K的表达,增加葡萄糖代谢,提高胞内ATP水平,抑制缺氧性神经细胞损伤。
红景天具有抗高原缺氧作用,其主要成分红景天苷对心脏和大脑等器官的缺血再灌注损伤可产生一定的保护作用[1]。文献报道,红景天苷可增强培养的人神经母细胞瘤细胞对抗缺氧性死亡的能力,起到细胞保护作用[2-3]。红景天苷还可促进神经干细胞向神经元细胞分化,并促进分化的神经元细胞生长[4]。可见,红景天苷对脑神经细胞具有良好的保护作用,为其临床应用提供了依据。不过,目前有关红景天苷神经保护作用的分子机制研究并不完全清楚。多数研究表明,缺氧诱导因子(HIF-1)介导红景天苷的多种生物学效应[5]。另外,JAK2-STAT3信号途径可介导红景天苷抗结肠癌的作用[6]。磷脂酰肌醇3激酶(PI3K)信号分子激活是红景天苷减弱心肌缺血再灌注损伤的细胞内主要分子机制[7]。已知,糖代谢和细胞内ATP生成变化对于细胞活性至关重要。本研究观察了红景天苷保护神经元细胞抗损伤过程中糖代谢和能量变化情况,进一步探讨红景天苷保护神经元细胞的机制。
材料和方法
1 材 料 Balb/c小鼠由第四军医大学实验动物中心提供。四氮唑盐(MTT)购自于碧云天生物科技有限公司。SYBR green荧光定量PCR试剂盒由Takara公司提供。己糖激酶检测试剂盒购自上海索宝生物科技有限公司。细胞ATP定量检测试剂盒和葡萄糖转运体-1(GLUT-1)检测试剂盒分别购自Sigma公司及武汉伊莱瑞特生物科技有限公司。
2 实验方法
2.1 细胞培养:出生5 d的雄性Balb/c小鼠处死后快速于冰冷条件下取大脑皮质,预冷的PBS冲洗后加入0.25%胰酶于37℃消化10 min,终止消化后,吹打组织成匀浆,离心去上清,加入培养液(DMEM/F12+20%FBS)接种,6 h后取出未贴壁神经元细胞,按实验要求种植于培养板中,次日加入阿糖胞苷(5g/ml)培养,获得原代神经元细胞。
2.2 原代神经元细胞处理:原代神经元细胞培养至对数生长期后进行实验处理,10 mg/ml红景天苷处理神经元细胞48 h观察其对神经元细胞的保护机制;之后红景天苷处理细胞48 h,再以1%低氧处理6 h建立缺氧模型,检测红景天苷对缺氧性神经元保护作用。以往的文献报道红景天苷的保护机制可能是通过PI3K通路实现[7],为了明确红景天苷对缺氧性神经细胞保护作用的分子机制,上述三组先以1mol/L PI3K抑制剂LY294002预处理48 h,再分别以红景天苷和(或)缺氧处理;同时以PI3K抑制剂LY294002单独处理及正常未处理细胞作为阴性对照或空白对照。
2.3 定量RT-PCR:神经元细胞经过处理后,去除上清,加入裂解液,用RNA提取试剂盒提供的试剂和步骤提取RNA进行反转录,随后应用定量PCR方法进行检测。所用引物的序列如下:己糖激酶(HK)的上游引物5’-CAT GAG GAA GAT GCT TGC CG-3’,下游引物5’-CTA CCA GCA CTC TGC TTG CC-3’。GLUT-1的上游引物5’- GCC CCC AGA AGG TTA TTG A -3’,下游引物5’- CGT GGT GAG TGT GGT GGA T -3’。PI3K的上游引物5’- ACA GAT TGA ATC GCA CAC GC -3’,下游引物5’- GAG GAA TGG ATG AGG GCG TC -3’。内对照18S的上游引物5’-CGG CTA CCA CAT CCA AGG AA-3’,下游引物5’-GCT GGA ATT ACC GCG GCT-3’。采用Delta-delta Ct法对基因表达水平差异进行分析。
2.4 HK活性测定:细胞种植于6孔板,分组处理后,去除培养液,用PBS洗细胞两次,加入匀浆缓冲液刮细胞至微量离心管内超声破碎细胞离心细胞上清液用于酶活性测定。己糖激酶活性测定采取6-磷酸葡萄糖脱氢酶偶联比色法,反应体系3 ml,充分混匀后,30℃水浴 5 min,再加入细胞上清液50 μl进行反应,在340 nm处读取吸光度的变化,每30 s记录1次,连续记录3 min,测出每分钟光密度增加值。与标准曲线对比计算己糖激酶活性。
2.5 细胞内ATP水平测定:根据荧光素酶化学发光法测定ATP水平。细胞种植于6孔板分组处理后去除培养液,每孔加入200l裂解液,裂解后12 000×g离心5~10 min,取上清用于ATP水平测定。加100l ATP检测工作液到检测孔或检测管内。室温放置3~5 min,随后在检测孔或检测管内加上100l样品或标准品,迅速混匀,立即用化学发光测定仪进行化学发光强度测定。各个样本的化学发光测定值与标准曲线对比计算ATP水平。
2.6 GLUT-1表达水平检测:细胞种植于96孔板,分组处理后,取细胞培养上清于1000 r/min离心20 min,除去杂质及细胞碎片。取上清在450 nm处检测。以标准品的浓度为横坐标,OD值为纵坐标,绘出标准曲线。通过标准曲线计算样品中GLUT-1的表达水平。
2.7 细胞活性测定:小鼠神经元细胞均匀种植于96孔板中,在培养2 d后,加药处理后,在收取细胞前4 h加入MTT(5 mg/ml),37℃孵育4 h,随后去除细胞上清,加入二甲基亚砜(DMSO)(100l/孔)溶解MTT结晶,脱色摇床混匀后在酶标仪上检测560 nm处的吸光度。根据吸光度反映细胞活性。
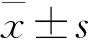
结 果
1 红景天苷诱导PI3K、HK、GLUT-1的基因表达 见图1。红景天苷(10 mg/ml)处理2 d后,小鼠神经元细胞内HK、GLUT-1基因表达水平较对照组显著升高(P<0.05)。同时,代谢相关调节基因PI3K表达也显著升高(P<0.05)。
2 红景天苷增加神经元细胞HK活性和细胞内ATP水平 红景天苷(10 mg/ml)处理2 d后,小鼠神经元细胞内HK活性较对照组显著升高(P<0.05),图2A。小鼠神经元细胞的胞内ATP水平也较对照组显著升高(P<0.05),图2B。
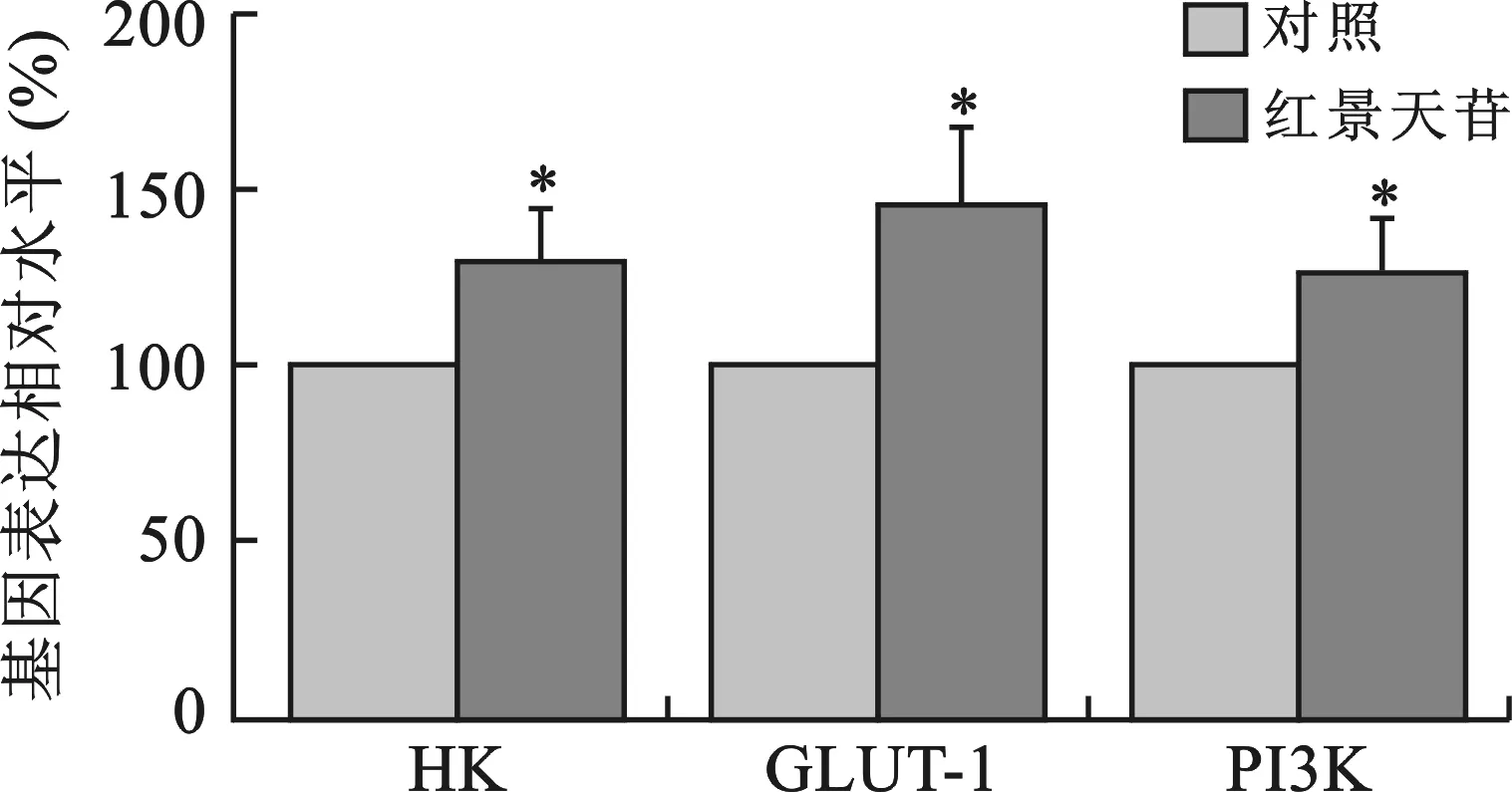
与对照组比较,*P<0.05
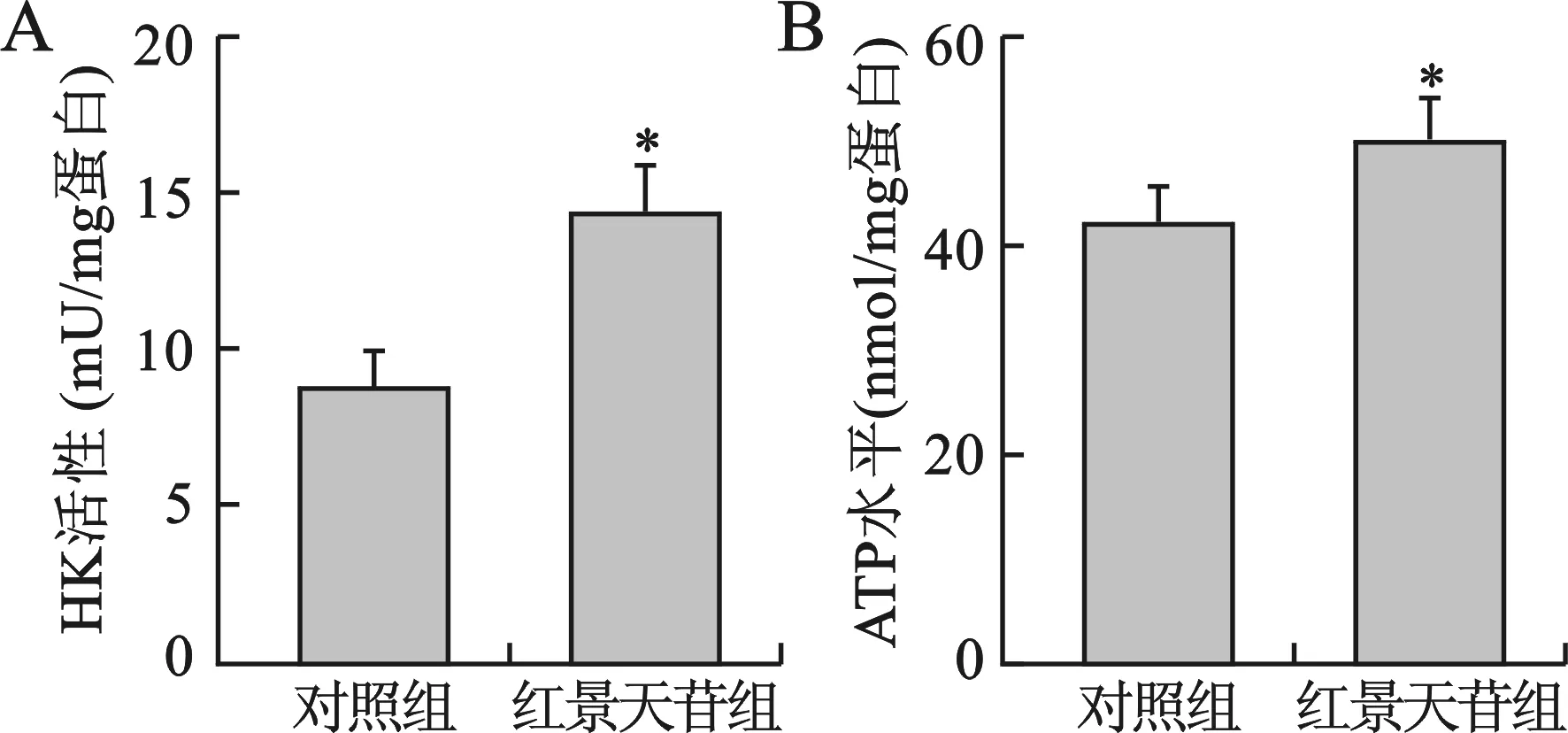
与对照组比较,*P<0.05
3 红景天苷对缺氧性神经元细胞损伤的保护作用 见图3。急性缺氧可导致小鼠神经元细胞出现明显的死亡,MTT检测缺氧组(在1%低氧状态下处理6 h)、缺氧+红景天苷组(红景天苷10 mg/ml,预先处理2 d后再缺氧处理6 h)细胞活性检测均低于对照组(P<0.01)。缺氧组细胞死亡率达到80%,缺氧+红景天苷组神经元细胞的死亡率降低到65%(P<0.01)。

与对照组比较,*P<0.01
4 PI3K抑制可消除红景天苷对神经元细胞的保护作用 见图4。LY294002+红景天苷组,红景天苷(10 mg/ml)处理较LY294002组不能够升高神经元细胞内HK、GLUT-1表达,同时细胞内ATP水平也无明显升高(P>0.05),结果见图4A~C。在LY294002+缺氧组,缺氧所致神经元细胞的死亡率达到88%,LY294002+红景天苷+缺氧组在红景天苷(10 mg/ml)预处理2 d死亡率也高达85%,与LY294002+缺氧组无显著区别(P>0.05),红景天苷不再显著降低缺氧所致神经元细胞的死亡率,结果见图4D。
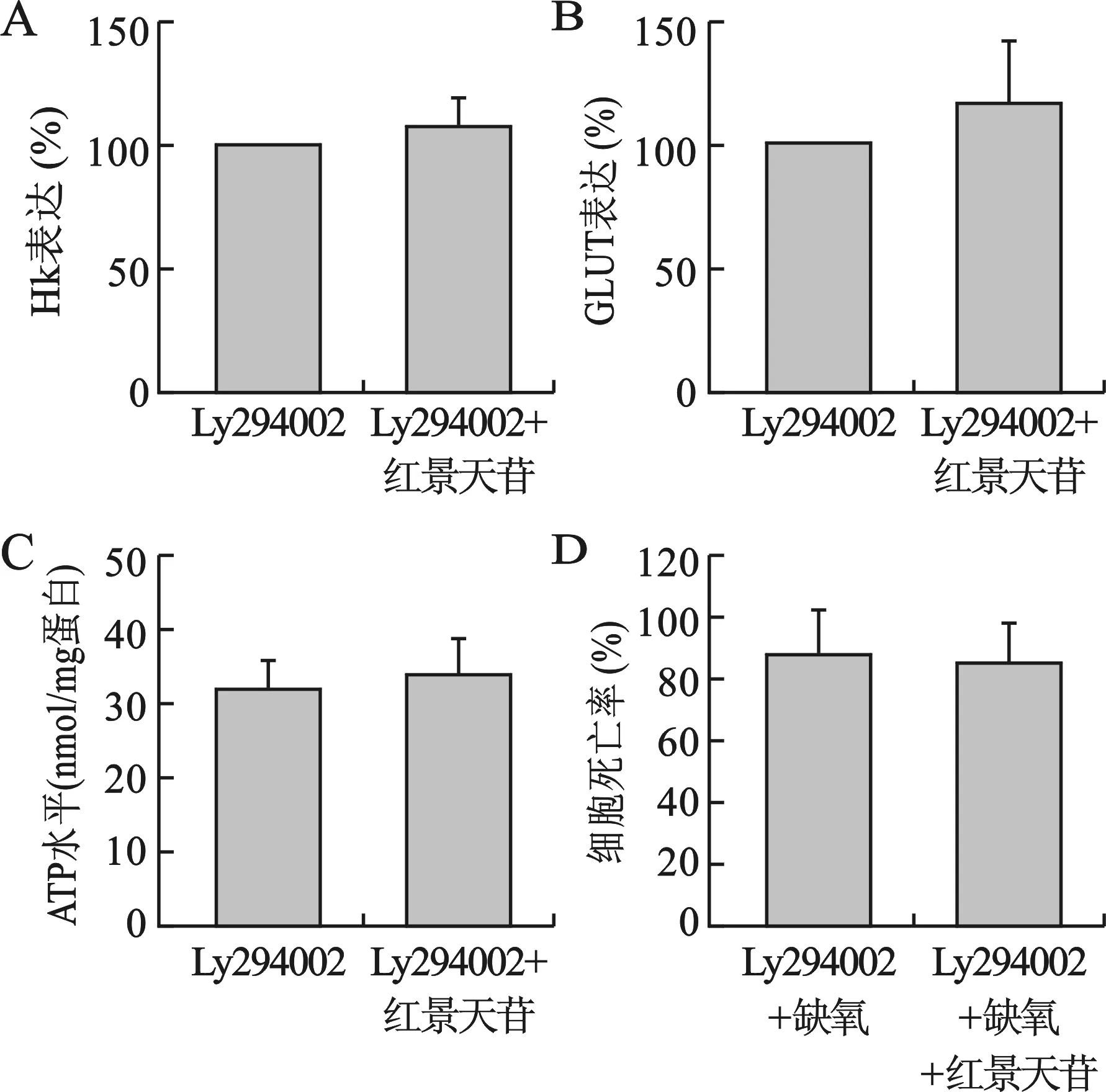
图4 PI3K抑制对红景天苷神经元保护作用的影响
讨 论
本研究首先明确了红景天苷具有明确的神经保护作用。以往多项研究从在体水平明确了红景天苷具有减少大脑缺血再灌注损伤的作用,在离体细胞水平也有一些研究表明红景天苷可减少缺氧等刺激对神经细胞的损伤。本研究在小鼠原代培养的神经元细胞发现红景天苷可减少急性缺氧导致的神经元细胞死亡,这进一步证实了红景天苷的保护作用,表明红景天苷是一个神经保护剂,具有良好的应用前景。
虽然红景天苷的作用十分明确,但是保护作用的机制还有待深入研究。现已明确能量代谢异常是细胞死亡的一个主要原因。葡萄糖作为神经元细胞的主要能源物质,经葡萄糖转运体(GLUT)进入细胞后被HK活化为6-磷酸葡萄糖,进而进入糖代谢的一系列过程中,最终在线粒体氧化磷酸化生成ATP。糖代谢异常和随之ATP生成减少成为许多种细胞经受不同损伤后发生死亡的一个共同表现[8]。本研究观察了红景天苷对神经元细胞代谢的调节作用,发现红景天苷上调GLUT-1、HK等表达,并提高HK活性和细胞内ATP水平。胞内ATP水平升高可对抗缺氧所致的代谢下降,起到一定的细胞保护作用,本研究认为这是红景天苷具有神经保护作用的一个重要原因。
本研究进一步发现,介导红景天苷代谢调节作用的一个关键信号分子是PI3K。PI3K是磷脂酰肌醇家族中一个重要成员,可催化产生磷酸化的磷脂酰肌醇,进而激活下游分子,调节细胞的增殖分化、黏附迁移、代谢活性等[9]。目前已知,PI3K与细胞的物质代谢尤其是糖代谢关系密切[9]。本研究发现,原代培养神经元细胞经红景天苷处理可高表达PI3K。PI3K高表达伴随着糖代谢关键蛋白HK和GLUT-1高表达。PI3K表达被抑制后,红景天苷增加HK和GLUT-1高表达的作用也被削弱,其对神经元保护作用也相应减弱。由此可见,PI3K是神经元糖代谢中一个关键分子,并通过促进糖代谢增加细胞内ATP水平而产生抗损伤作用。
总之,本研究在细胞水平进一步证实红景天苷具有神经保护作用,并且表明,PI3K介导的糖代谢增强是红景天苷保护作用的一个重要原因。
[1] 赖文芳,张小琴,洪海棉,等.红景天苷对大鼠局灶性脑缺血/再灌注损伤的神经保护作用[J].中国药理学通报,2015,31(6):775-780.
[2] 张文生,朱陵群,牛福玲,等.红景天苷对缺氧/缺糖损伤神经细胞的保护作用[J].中国中药杂志,2004,29(5):79-82.
[3] 祁存芳,张军峰,陈新林,等.红景天苷通过抑制凋亡相关蛋白的表达保护缺氧对培养神经干细胞的损伤[J].南方医科大学学报,2013,33(7):962-966.
[4] 李海龙,吴守振,李 洺,等.红景天苷对百草枯诱发PC12细胞损伤的保护作用[J].陕西中医,2015,36(7):925-928.
[5] 张金平,陈建宗,刘安恒,等.红景天苷通过HIF-1途径对缺氧诱导心肌细胞凋亡的抑制作用[J].第四军医大学学报,2008,29(12):1057-1060.
[6] Sun KX, Xia HW, Xia RL. Anticancer effect of salidroside on colon cancer through inhibiting JAK2/STAT3 signaling pathway [J]. Int J Clin Exp Pathol,2015,8(1):615-621.
[7] 张金平,陈建宗,刘安恒,等.红景天苷通过PI(3)K/Akt激活HIF-1α表达抑制缺氧诱导的心肌细胞凋亡[J].细胞与分子免疫学杂志,2008,24(4):406-408.
[8] Seppet E, Gruno M, Peetsalu A,etal. Mitochondria and energetic depression in cell pathophysiology[J]. International Journal of Molecular Sciences,2009, 10(5): 2252-2303.
[9] 迟毓婧,李 晶,管又飞,等.PI3K-Akt信号传导通路对糖代谢的调控作用[J].中国生物化学与分子生物学报,2010,26(10):879-885.
(收稿:2016-11-22)
红景天苷 脑损伤 缺氧,脑 神经保护
R651.15
A
10.3969/j.issn.1000-7377.2017.06.008
猜你喜欢
今日农业(2022年14期)2022-09-15 01:45:16
中成药(2021年5期)2021-07-21 08:39:04
中成药(2018年7期)2018-08-04 06:03:58
知识经济·中国直销(2017年12期)2018-01-03 08:21:20
哈尔滨医药(2016年3期)2016-12-01 03:58:41
中国卫生标准管理(2015年16期)2016-01-20 09:26:30
国际心血管病杂志(2015年5期)2015-02-27 12:11:37
发明与创新(2015年37期)2015-02-27 10:40:25
西南军医(2015年1期)2015-01-22 09:08:31
中国药业(2014年24期)2014-05-26 09:00:11
