
小麦-滨麦2D/ 2Ns异代换系DM2411的分子细胞遗传学鉴定
2017-06-22 14:47赵继新谈宏斌王秀娟李家创梁邦平刁慧珊白宇皓陈新宏
麦类作物学报 2017年5期
刘 洋,赵继新,谈宏斌,王秀娟,李家创,梁邦平,刁慧珊,白宇皓,武 军,陈新宏
(1.西北农林科技大学农学院,陕西杨陵 712100; 2.陕西省种业集团有限责任公司,陕西杨凌 712100)
小麦-滨麦2D/ 2Ns异代换系DM2411的分子细胞遗传学鉴定
刘 洋1,赵继新1,谈宏斌2,王秀娟1,李家创1,梁邦平1,刁慧珊1,白宇皓1,武 军1,陈新宏1
(1.西北农林科技大学农学院,陕西杨陵 712100; 2.陕西省种业集团有限责任公司,陕西杨凌 712100)
为进一步挖掘利用滨麦优异基因,并丰富小麦遗传种质资源,利用形态学、细胞遗传学、基因组原位杂交(Genomicinsituhybridization,GISH)、EST-STS分子标记、SSR分子标记等技术,对从八倍体小滨麦M842-16和硬粒小麦D4286杂交F7代材料中筛选出的1个遗传稳定的小滨麦异代换系DM2411进行了鉴定。细胞遗传学观察表明,DM2411的染色体主要构型为2n=42=21Ⅱ,遗传稳定。根尖体细胞和花粉母细胞的原位杂交研究表明,DM2411含有1对滨麦Ns基因组。SSR分析表明,DM2411可能缺失了小麦2D染色体。EST分析表明,DM2411可能含有滨麦2Ns染色体。形态学调查表明,DM2411的株高极显著降低。
小麦;滨麦;代换系;GISH;EST-STS;SSR
小麦野生亲缘物种具有许多优良性状,通过远缘杂交可以将小麦亲缘物种的有益基因导入到普通小麦中,从而扩大小麦遗传变异,达到改良栽培小麦的目的。滨麦(Leymusmollis(Trin.)Hara,2n=28,NsNsXmXm),是禾本科(Poaceae)小麦族(Triticeae)大麦亚族(Hordinae)赖草属(LeymusHochst.)的一个多年生异花授粉植物[1],具有根系发达、茎秆粗壮、大穗多花、抗旱耐盐碱、抗多种细菌和真菌病害等优良特性[2-3]。滨麦基因组为NsNsXmXm,其中Ns基因组来自于新麦草属(PsathyrostachysKeng)[4-5],而Xm基因组的来源目前尚未确定。早在20世纪50-70年代,苏联科学家通过胚拯救技术获得了四倍体、六倍体小麦与大赖草、滨麦和沙生赖草的杂种,以及杂种的双二倍体和不完全双二倍体[6];Wang等[7]对6种类型的八倍体小滨麦的染色体组构成进行分子细胞遗传学分析,确定八倍体小滨麦M842-4、M842-8、M842-12和M842-16的染色体组构成为AABBDDNsNs,而M842-10的染色体组构成为AABBDDNsNs+2Ta-2Lm(Ns),M842-13 的染色体组构成确定为AABBDDJJ;Li等[8-9]分析了八倍体小滨麦小孢子的发生、配子体的形成及滨麦染色体的遗传行为,并利用八倍体小黑麦与八倍体小滨麦杂交,获得了一个包含6条黑麦染色体和6条滨麦染色体的小麦-黑麦-滨麦三属杂种。Wang等[10]根据细胞学和种子贮藏蛋白试验结果,确定八倍体小滨麦M842-1的染色体组成为AABBDDNsNs,抗病鉴定结果显示其对条锈病免疫,高抗白粉病。
本课题组自20世纪80年代开始进行普通小麦7182与滨麦的杂交研究,陈潄阳等[11]、傅 杰等[12-13]通过胚拯救和秋水仙素加倍技术获得普通小麦和滨麦的杂交种,并最终获得了具有3种穗型的八倍体小滨麦M842(AABBDDJJ或AABBDDNsNs);利用M842与普通小麦和缺体小麦杂交,培育出了3个异附加系和2个异代换系。在此基础上,赵继新等[14-15]利用八倍体小滨麦与硬粒小麦品种Trs-372和D4286进行杂交,对杂种F1进行了细胞遗传学分析;利用八倍体小滨麦M842-12与硬粒小麦品种Trs-372杂交,选育出1个含有6条滨麦染色体的小滨麦多重异代换系05DM6;利用小滨麦多重异代换系05DM6和普通小麦7182杂交得到了在千粒重和麦谷蛋白方面优于其亲本普通小麦7182的二体代换系DM45。
本研究在D4286×M842-16的后代中,筛选出了1个染色体数目为2n=42、含有2条滨麦染色体的株系DM2411。本研究拟采用形态学、细胞遗传学、基因组原位杂交、EST-STS分子标记和SSR分子标记等技术对该株系进行研究,以期了解株系DM2411的形态特征、遗传行为、染色体构成,及所含滨麦染色体的同源群归属等,为进一步挖掘利用滨麦优异基因、丰富小麦遗传资源及最终的小麦品种改良提供新的种质资源。
1 材料与方法
1.1 供试材料
供试材料包括滨麦(Leymusmollis(Trin.) Hara,2n=28,NsNsXmXm)、华山新麦草(Psathyrostachyshuashanica,2n=14,NsNs)、普通小麦7182(TriticumaestivumL.,2n=42,AABBDD)、硬粒小麦D4286 (Triticumdurum,2n=28,AABB)、八倍体小滨麦M842-16 (基因组AABBDDNsNs,2n=56)及M842-16与硬粒小麦D4286的F7代材料DM2411,均保存于西北农林科技大学农学院陕西省植物遗传工程育种重点实验室。其中,八倍体小滨麦M842-16是由普通小麦(TriticumaestivumL.)品种7182和滨麦经胚拯救、秋水仙素染色体加倍而获得的不完全二倍体。
1.2 试验方法
1.2.1 细胞遗传学分析
根尖体细胞染色体数目鉴定:于室内将数粒DM2411种子浸泡,待种子露白后移到垫有双层滤纸的培养皿内发芽(25 ℃);当根长至1~2 cm时用镊子夹取根尖放入冰水中处理24 h,然后用卡诺氏固定液I (乙醇∶乙酸=3∶1) 固定,于-20 ℃冰箱保存。待一周后,可将根尖转入70%酒精中,45%醋酸洋红染色制片,Olympus BX60显微镜观察并照相。
花粉母细胞减数分裂中期I (PMC MI)染色体构型观察:田间选取适龄幼穗,用卡诺氏固定液Ⅱ(乙醇∶氯仿∶冰乙酸=6∶3∶1)固定24 h即刻转入70%酒精,45%醋酸洋红染色制片,Olympus BX60显微镜观察并照相。
1.2.2 基因组原位杂交(GISH)
DNA提取及探针标记:采用CTAB法[16 ]提取供试材料全基因组DNA。依照Dig-Nick-Translation Mix试剂盒(Roche,Germany)使用说明书对华山新麦草的全基因组DNA进行标记。
染色体制片:将DM2411的根尖体细胞和花粉母细胞用45%醋酸压片,液氮冷冻揭片,气干备用。
GISH参照Han等[17]的方法进行。每张制片加20 μL杂交液,包含20×SSC 2 μL、ssDNA(鲑鱼精DNA,5 μg·μL-1) 0.5 μL、10% SDS(十二烷基磺酸钠) 0.5 μL、50%(W/V)DS(硫酸葡聚糖) 4 μL、去离子甲酰胺10 μL、探针DNA 100 ng。80 ℃变性8~10 min,然后转至恒温箱37 ℃杂交过夜(16~24 h)。探针杂交信号用荧光素(FITC)检测,洗脱后用PI复染,加抗褪色剂H-1300后盖上盖玻片,用Olympus BX60荧光显微镜检测,在蓝光下,外源染色体呈现黄绿色,小麦染色体呈现红棕色,最后用Olympus BX60荧光显微镜观察,Pixera Penguin 150CL显微数码相机照相。
1.2.3 SSR和EST-STS标记分析
SSR标记来源于Röder等[18]和Pestsoval等[19]。EST-STS标记来源于http://wheat.pw.usda.gov/SNP/new/pcr-primers.shtm.网站。PCR反应体系为20 μL,包含1×buffer (100 mmol·L-1Tris-HCl,pH 8.3;1.5 mmol·L-1MgCl2)、0.2 mol·L-1dNTPs、50 ng引物、1 UTaqDNA聚合酶、50~100 ng模板DNA。PCR反应在PTC-200 (Bio-Rad,USA)上进行,扩增程序:94 ℃预变性3 min;94 ℃变性1 min,50/55/60 ℃退火1 min,72 ℃延伸1 min,32~35个循环;72 ℃延伸10 min,4 ℃或者15 ℃结束。扩增产物经8%非变性聚丙烯酰胺凝胶进行电泳。点样量为4 μL,150~200 V恒压条件下电泳2~3 h。硝酸银染色10 min,显影液显色后立即用数码相机照相,保存结果。
1.2.4 农艺性状调查
随机选取八倍体小滨麦M842-16、硬粒小麦品种D4286、DM2411以及普通小麦品种7182的10个单株,调查统计每个单株的株高、穗长、小穗数、穗粒数、结实率等农艺性状,并利用SPASS Statistic 20对各性状进行差异显著性分析。
2 结果与分析
2.1 DM2411的选育及其细胞遗传学分析
在八倍体小滨麦M842-16与硬粒小麦品种D4286杂交F7代材料中,通过根尖体细胞镜检,筛选到1株2n=42的植株DM2411。以华山新麦草全基因组DNA为探针进行GISH鉴定,结果显示,DM2411含有2条完整的黄绿色染色体信号,推断其应含有2条来源于滨麦的Ns染色体。随机选取DM2411的14粒种子在室内发芽,将根尖固定后播种于大田,经镜检发现14粒种子的根尖体细胞的染色体数目都为2n=42,GISH鉴定表明所有种子都含有2条Ns染色体(图1)。
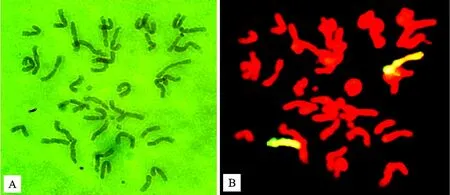
图1 DM2411根尖体细胞有丝分裂中期染色体(A)及其原位杂交鉴定结果(B)
田间选取DM2411的适龄幼穗,对56个花粉母细胞减数分裂中期Ⅰ染色体配对情况进行观察。结果(图2A、表1)显示,DM2411有48个花粉母细胞可以正常分裂,并形成2n=21Ⅱ的染色体构型,6个花粉母细胞具有20个二价体和2个单价体,2个花粉母细胞具有19个二价体和4个单价体。以华山新麦草全基因组DNA作为探针,对其花粉母细胞进行GISH鉴定,结果显示,DM2411花粉母细胞在减数分裂中期Ⅰ都有1个完整的二价体,呈现黄绿色信号,表明DM2411中的2条Ns染色体可以正常配对形成1个二价体(图2B)。由此说明,DM2411是一个遗传稳定的、包含40条小麦染色体和1对滨麦Ns染色体的小麦-滨麦二体异代换系。

图2 DM2411花粉母细胞减数分裂中期I染色体(A)及其原位杂交鉴定结果(B)

观察的细胞数Observedcells染色体数Chromosomenumber染色体构型Chromosomeconfiguration21Ⅱ20Ⅱ+2Ⅰ19Ⅱ+4Ⅰ染色体配对ChromosomeparingⅠⅡ环状Ring棒状Rod总数Total564248620.36(0~4)17.76(16~21)3.06(0~5)20.82(19~21)
2.2 DM2411的SSR分析
利用Röder等[18]和Pestsova等[19]开发的分布于小麦1D~7D染色体上的SSR标记对DM2411进行了分析,结果(图3)显示,小麦1D、3D、4D、5D、6D和7D染色体上的SSR引物在DM2411和普通小麦品种7182中都扩增出了预期的目标条带,而2D染色体上的SSR引物,虽然在小麦品种7182中扩增出了预期的目标条带,但却不能在DM2411中扩增出预期的目标条带,说明DM2411应该含有小麦1D、3D、4D、5D、6D和7D染色体,而缺失了小麦2D染色体。
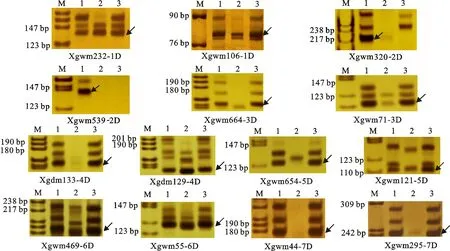
M:DNA ladder;1:普通小麦7182;2:硬粒小麦D4286;3:异代换系DM2411;箭头指示目标条带。
M:DNA ladder; 1:Triticumaestivumcv. 7182; 2:Triticumdurumcv. D4286; 3:Alien substitution line DM2411;The arrows show the target bands.
图3 DM2411的SSR分子标记分析结果
Fig.3 The results of SSR analysis on alien substitution line DM2411
2.3 DM2411的EST-STS分析
利用源于http://wheat.pw.usda.gov/SNP/new/pcr-primers.shtm.网站的分布于小麦1D~7D染色体上的EST-STS标记对小滨麦二体异代换系DM2411所含外源染色体的同源群归属进行了分析,结果(图4)显示,第1、3、4、5、6、7同源群的特异EST-STS引物在普通小麦7182、硬粒小麦D4286和异代换系DM2411中均扩增出了预期的目标条带,但却不能在滨麦和华山新麦草中扩增出特异条带;第2同源群上的3对EST-STS引物BE404332、BQ159450和BQ159532在异代换系DM2411中既扩增出了小麦基因组的预期目标条带,同时也扩增出了滨麦和华山新麦草Ns基因组的特异条带。由此推断该二体异代换系DM2411应该含有滨麦的2Ns染色体,说明DM2411为小麦-滨麦2Ns(2D)异代换系。
2.4 DM2411及其亲本的形态学特征
对DM2411及其亲本普通小麦品种7182、硬粒小麦品种D4286和八倍体小滨麦M842-16的形态学特征进行了调查,结果(表2、图5)表明,DM2411的株高较亲本明显变矮,差异达极显著水平;穗长显著长于亲本7182和D4286;分蘖数显著低于亲本;穗粒数和结实率显著低于亲本7182和D4286;小穗数显著大于亲本7182和D4286;穗形与亲本7182较为接近,均为方形穗。

M:DNA ladder;1:普通小麦7182;2:硬粒小麦D4286;3:异代换系DM2411;4:滨麦;5:华山新麦草;箭头指示特异目标条带。
M:DNA ladder; 1:Triticumaestivumcv.7182; 2:Triticumdurumcv. D4286; 3:Alien substitution line DM2411; 4:Leymusmollis; 5:Psathyrostachyshuashanica; The arrows show the target bands.
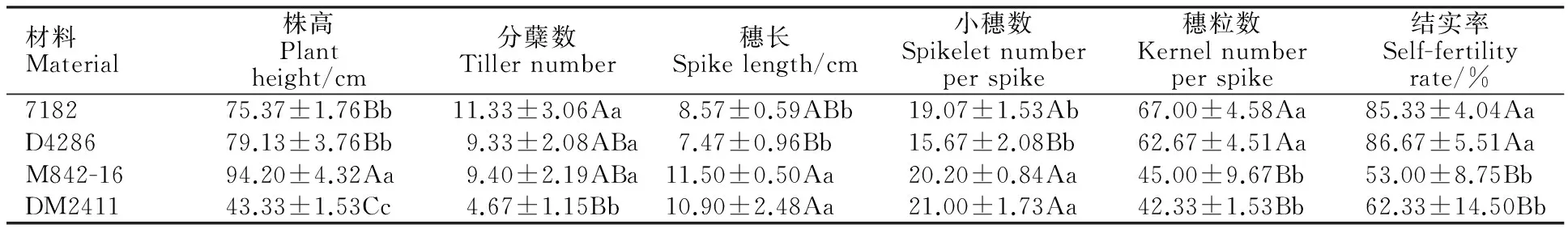
图4 DM2411的EST-STS分子标记分析结果
数据后的大、小写字母不同表示不同材料间的差异在0.01和0.05水平上显著。
Different capital and small letters indicate significant differences at 0.01 and 0.05 levels between the substitution line DM2411 and its parents,respectively.

1:普通小麦7182;2:异代换系DM2411。
1:Triticumaestivumcv. 7182; 2:Alien substitution line DM2411.
图5 异代换系DM2411及其亲本普通小麦7182的植株和穗子
Fig.5 Plant and spike of alien substitution line DM2411 and its parentTriticumaestivumcv.7182
3 讨 论
本研究利用八倍体小滨麦M842-16与硬粒小麦品种D4286杂交,经过7代自交,在其后代中筛选出1个染色体数目为2n=42,且包含2条滨麦Ns基因组的小麦-滨麦异代换系DM2411。细胞学观察表明,DM2411的染色体主要构型为2n=21Ⅱ,遗传稳定。根尖体细胞和花粉母细胞的原位杂交研究表明,DM2411含有1对滨麦Ns染色体。本研究中八倍体小滨麦的A和B基因组与来自硬粒小麦的A和B基因组,理论上可以正常配对且能完整遗传,而来自八倍体小滨麦的D和Ns基因组,在杂交后代自交过程中会出现不配对而随机分离的遗传现象。SSR是鉴定小麦遗传背景中外源染色体的有力工具。为了确定DM2411所缺失的小麦染色体,本研究采用了Röder等[18]和Pestsova等[19]开发的来自于粗山羊草D基因组的SSR标记对小滨麦异代换系DM2411进行了分析,结果显示,小滨麦异代换系DM2411可能缺失了小麦2D染色体。EST和STS数据库中大量定位于不同染色体区段的EST-STS位点为小麦不同基因组间部分同源关系的分析提供了简单高效的方法。本研究采用了在小麦A、B、D染色体组同源群之间有共显性的EST-STS标记,结果显示,异代换系DM2411应含有与滨麦第2同源群染色体相同的遗传物质,即含有滨麦的2Ns染色体。
株高作为小麦的一个重要农艺性状,我国早在20世纪70年代就开始了矮化育种。刘兆晔等[20]认为小麦品种适当矮化不仅可以防止倒伏,更重要的是削减了部分茎、叶鞘的生长,有利于籽粒生长,从而提高经济产量。Du等[21-22]利用GISH结合EST-STS和SSR分子标记从小麦和华山新麦草的杂交后代中鉴定出一个2Ns附加系和一个2Ns(2D)二体异代换系,两个材料的株高较亲本在0.05水平上都显著降低,穗长都显著增长,千粒重显著增加。本研究中DM2411中含有的Ns基因组来源于滨麦,而滨麦中的Ns基因组来源于新麦草属。DM2411的株高显著降低,平均株高为43.33 cm,极显著地低于亲本,与Du等[21-22]的试验结果相吻合,可以推断2Ns染色体具有降秆作用,可能含有矮秆基因,或是与小麦的其他染色体发生基因的互作而使植株变矮;但该材料结实率较亲本有所降低,这与之前华山新麦草的2Ns基因组可以降低株高的同时也增加了千粒重有所不同,产生这种现象的原因可能是滨麦的2Ns基因组与华山新麦草的2Ns基因组在普通小麦背景中的遗传效应有所不同。
滨麦具有多种有益基因,对抗病性和改善农艺性状都具有潜在的利用价值。该小滨麦异代换系的育成,可作为一个蕴含矮秆基因的中间材料,为进一步创制新的小麦矮秆种质、研究滨麦染色体的形态结构特征及遗传机理、挖掘利用滨麦优异基因、丰富小麦遗传种质资源具有重要价值。
[1]WANG R R,HSIAO C.Morphology and cytology of interspecific hybrids ofLeymusmollis[J].JournalofHeredity,1984,75(6):488.
[2]FORSSTRM P O,MERKER A.Sources of wheat powdery mildew resistance from wheat-rye and wheat-Leymushybrids [J].Hereditas,2001,134(2):115.
[3]KISHII M,WANG R R,TSUJIMOTO H.Characteristics and behaviour of the chromosomes ofLeymusmollisandL.racemosus(Triticeae,Poaceae) during mitosis and meiosis [J].ChromosomeResearch,2003,11(8):741.
[4]DEWEY D R.Genome relations among diploidElymusjunceusand certain tetraploid and octoploidElymusspecies [J].AmericanJournalofBotany,1970(6):633.
[5]DEWEY D R.The genome constitution and phylogeny ofElymusambiguous[J].AmericanJournalofBotany,1976,63(5):626.
[6]ANAMTHAWAT-JNSSON K.Wide-hybrids between wheat and lymegrass:breeding and agricultural potential [J].IcelandicAgriculturalSciences,1995,9:101.
[7]WANG X P,FU J,ZHANG X Q,etal.Molecular cytogenetic study on genome constitutions of octoploidTritileymus[J].ActaBotanicaSinica,2000,42(6):582.
[8]LI X F,LIU S B,GAO J R,etal.Abnormal pollen development of bread wheat-Leymusmollispartial amphiploid [J].Euphytica,2005,144(3):247.
[9]LI X F,SONG Z Q,LIU S B,etal.Cytogenetic study of a trigeneric (triticale×trileymus) hybrid [J].Euphytica,2006,150(1):117.
[10]WANG J,CHEN X H,DU W L,etal.Morphological and molecular cytogenetic characterization of partial octoploidTritileymus[J].GeneticResourcesandCropEvolution,2013,60(4):1453.
[11] 陈漱阳,付 杰,高立贞.普通小麦与滨麦的杂交[J].西北植物学报,1985(4):260.
CHEN S Y,FU J,GAO L Z.The hybridization betweenTriticumaestivumandLeymusmollis[J].ActaGeneticaSinica,1985(4):260.
[12] 傅 杰,陈漱阳,张安静.八倍体小滨麦的形成及细胞遗传学研究[J].遗传学报,1993,20(4):317.
FU J,CHEN S Y,ZHANG A J.Study of the formation and cytogenetic of octoploidTritileymus[J].ActaGeneticaSinica,1993,20(4):317.
[13] 傅 杰,陈漱阳,张安静,等.八倍体小滨麦与普通小麦杂交后代的细胞遗传学研究[J].遗传学报, 1996,23(1):24.
FU J,CHEN S Y,ZHANG A J,etal.Cytogenetic studies on the cross progenies between octoploidTritileymusandTriticumaestivum[J].ActaGeneticaSinica,1996,23(1):24.
[14] 赵继新,陈新宏,王小利,等.普通小麦-华山新麦草异代换系的分子细胞遗传学研究[J].西北植物学报,2004,24(12):2277.
ZHAO J X,CHEN X H,WANG X L,etal.Cytogenetic and molecular identification of a wheat-P.huashanicaalien substitution line [J].ActaBotanicaBoreali-OccidentaliaSinica,2004,24(12):2277.
[15]ZHAO J,WANG X,PANG Y,etal.Molecular cytogenetic and morphological identification of a wheat-L.mollis,1Ns(1D) substitution line,DM45 [J].PlantMolecularBiologyReporter,2016,34(6):1.
[16]UBAYASENA K.Ready-to-use DNA extracted with a CTAB method adapted for herbarium specimens and mucilaginous plant tissue [J].PlantMolecularBiologyReporter,2006,24:161.
[17]HAN F,BAO L,FEDAK G,etal.Genomic constitution and variation in five partial amphiploids of wheat-Thinopyrumintermedium,as revealed by GISH,multicolor GISH and seed storage protein analysis [J].TheoreticalandAppliedGenetics,2004,109(5):1070.
[18]RÖDER M S,KORZUN V,WENDEHAKE K,etal.A microsatellite map of wheat [J].Genetics,1998,149(4):2007.
[19]PESTSOVA E,GANAL M W,RÖDER M S.Isolation and mapping of microsatellite markers specific for the D genome of bread wheat [J].Genome,2000,43(4):689.
[20] 刘兆晔,于经川,辛庆国.小麦株高问题的探讨[J].山东农业科学,2014(3):130.
LIU Z H,YU J C,XIN Q G.Study of wheat height [J].ShandongAgricultureScience,2014(3):130.
[21]DU W,WANG J,WANG L,etal.Molecular characterization of a wheat-Psathyrostachyshuashanica,Keng 2Ns disomic addition line with resistance to stripe rust [J].MolecularGeneticsandGenomics,2014,57(5):37.
[22]DU W,ZHAO J,WANG J,etal.Cytogenetic and molecular marker-based characterization of a wheat-Psathyrostachyshuashanica,Keng 2Ns(2D) substitution line [J].PlantMolecularBiologyReporter,2015,33(3):414.
Molecular Cytogenetic and Morphological Identification of a Wheat-Leymusmollis2Ns (2D) Substitution Line DM2411
LIU Yang1,ZHAO Jixin1,TAN Hongbin2,WANG Xiujuan1,LI Jiachuang1,LIANG Bangping1,DIAO Huishan1,BAI Yuhao1,WU Jun1,CHEN Xinhong1
(1.College of Agronomy,Northwest A&F University,Yangling,Shaanxi 712100,China;2.Shaanxi Seed Group Co.,Ltd.,Yangling,Shaanxi 712100,China)
A wheat-L.mollisalien substitution line DM2411 was identified from the F7progenies of octoploidTritileymusM842-16 andT.durumD4286 by using morphology,cytogenetics,genomicinsituhybridization (GISH),expressed sequence tag-sequence tagged site(EST-STS) and simple sequence repeat (SSR). Cytological studies demonstrated that DM2411 had a chromosome karyotype of 2n=42=21Ⅱ.GISH analysis indicated that DM2411 had a pair of Ns chromosomes fromL.mollis. Analysis with SSR and EST-STS markers showed that the wheat 2Ds chromosomes were substituted by a pair of 2Ns chromosomes fromL.mollisin DM2411. Agronomic trait evaluations showed that the plant height of DM2411 was dramatically lowered. Therefore,the substitution line DM2411 was a dwarf material,which could be exploited as an intermediate material in wheat breeding programs and genetic resources.
Wheat;L.mollis; Substitution line; GISH; EST-STS; SSR
时间:2017-05-12
2017-01-13
2017-04-13
陕西省自然科学基金项目(2015JM3095);西北农林科技大学唐仲英育种基金项目
E-mail:yyangliu@outlook.com
陈新宏(E-mail:cxh2089@126.com)
S512.1;S330
A
1009-1041(2017)05-0571-07
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170512.1955.002.html
猜你喜欢
作物学报(2022年2期)2022-11-06
西南农业学报(2022年5期)2022-06-06
中国糖料(2021年3期)2021-07-13
华北农学报(2021年1期)2021-03-26
快乐语文(2020年14期)2020-07-04
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
西南农业学报(2018年9期)2018-10-19
西部(2016年6期)2016-05-14
今日农药(2016年5期)2016-05-14
