
木麻黄青枯病菌的分离及强致病菌株的筛选
2017-06-19 18:29:46许秀玉甘先华仲崇禄张华新
林业科学研究 2017年3期
许秀玉,徐 斌,甘先华,仲崇禄,张华新
(1. 中国林业科学研究院国家林业局盐碱地研究中心, 北京 100091;2. 广东省森林培育与保护利用重点实验室,广东省林业科学研究院,广东 广州 510520;3. 中国林业科学研究院热带林业研究所, 广东 广州 510520)
木麻黄青枯病菌的分离及强致病菌株的筛选
许秀玉1,2,徐 斌2,甘先华2,仲崇禄3,张华新1*
(1. 中国林业科学研究院国家林业局盐碱地研究中心, 北京 100091;2. 广东省森林培育与保护利用重点实验室,广东省林业科学研究院,广东 广州 510520;3. 中国林业科学研究院热带林业研究所, 广东 广州 510520)
[目的]筛选出强致病菌株用于木麻黄抗病育种研究工作。[方法] 在广东沿海木麻黄青枯病发病区采集病根,开展病原菌两种不同分离方法的比较研究,对分离出的31个病株进行16s rRNA测序鉴定及致病性测定。[结果] 采用稀释分离法及根系溢出法在TTC培养基上共分离出了31个病原菌株,根系溢出法操作简便,杂菌含量低,分离率在60%左右,可作为常规稀释分离法的补充。31个菌株进行分子鉴定,只有22个菌株扩增出了特异性条带,经测序比对确定这22个菌株为青枯菌。青枯菌株致病性测定结果显示菌株致病性在无性系间、菌株间及菌株与无性系间的交互作用均具有极显著差异(P<0.01),不同接种方法间菌株致病性相关系数值较小,介于0.496 6 ~ 0.731 0之间,即室内水培接种与小苗盆栽接种不存在密切的直线相关关系。[结论] 综合选择在不同无性系及不同接种方法中均具有较强致病性的GL-2、H、M、TC-1、F、Q菌株作为下一步木麻黄种质资源抗性鉴定及抗病育种研究试验菌株。
木麻黄;青枯菌;分离;分子鉴定;菌株筛选
植物细菌性青枯病由青枯劳尔氏菌(Ralstoniasolanacearum)引起的毁灭性土传病害,是世界范围内传播广泛、危害严重、最难防治的细菌性重大病害之一[1]。该病原菌菌系复杂,寄主范围广,可危害农作物、果树、林木、花卉、药材、牧草、杂草等54个科的450余种植物[2]。
近二十年来,木麻黄已成为广东沿海地区不可替代的当家树种,生产上利用木麻黄A13及A8无性系大面积造林,使得林分遗传多样性单一,防护林带退化,林木生长衰退,木麻黄青枯病迅速扩展蔓延,严重威胁整个沿海防护林体系。抗病育种目前被认为是防治木麻黄青枯病的根本途径[3]。 我国木麻黄青枯病抗病育种研究基础薄弱,在抗源、抗性遗传规律、杂交育种、分子标记及基因工程等方面研究技术几乎都是空白,相关研究工作集中在上世纪末,如接种方法及接种条件的研究[4-5],木麻黄不同树种或种源的青枯病抗病能力评价[6-8],抗病优良品系的筛选[4,9-11]等。由于青枯菌能与植物、环境相互作用,协同进化产生变异[12],原来选育出的A8、A13、501等一批抗病无性系随着时间推移抗病性大大降低,强致病性菌株也逐渐失去其致病性。因此,目前要开展木麻黄抗病育种研究工作,首先要分离、获得有效的强致病性菌株。
长期以来,木麻黄青枯菌株的鉴定及毒性鉴别采用的是简单的TTC培养基法[4-5,8-9,13],即利用TTC培养基特异性区分青枯菌和其他细菌,并根据菌落生长形态来判断其致病性的强弱[14]。研究表明,青枯菌致病性严重分化,菌株间存在着显著的致病性差异,且在木麻黄—青枯菌的病理系统中,同时存在水平抗性与垂直抗性[13,15],因此利用TTC培养基传统肉眼观察判断毒性菌株的方法可靠性较差,在进行实验时有必要对其致病性进行检测。目前16S rRNA 基因检测技术已成为病原细菌检测和鉴定的重要方法[16-20],但在木麻黄青枯菌检测鉴定中尚未见类似报道。本研究将16S rRNA 基因检测技术应用于木麻黄青枯菌株的筛选,将根系溢出法用于木麻黄青枯菌的分离,利用水培接种及盆栽接种对分离出的菌株进行致病性测定,筛选出有效的强致病菌株用于进一步木麻黄选育试验,为相关领域科研人员筛选木麻黄强致病青枯菌株提供参考。
1 材料与方法
1.1 病害标本采集
选择广东电白、吴川、湛江南三岛、湛江东海岛及徐闻等有代表性的木麻黄新发病区林分,采样时选择田间自然发病的新发病植株,挖取木质部呈水渍状或半透明状根系2~3段,每段10 ~ 20 cm,封口袋封装,记录采集时间、地点,带回试验室分离病菌。具体病害标本采集地点见表1。

表1 菌株编号与来源
1.2 病原菌不同分离方法的比较
采用两种分离方法。稀释分离法参考朱圣杰[21]并加以改进:取3cm长病根,洗净,75%的酒精浸泡30 s,无菌水冲洗3次,在超净工作台上剥去外皮,切除两端,最后将其横切成3~5份,放入含有5~10 mL无菌水的培养皿内30 min,用移植环蘸取菌液划线培养于TTC培养基上。根系溢出法:取3~6 cm长的木麻黄病根,洗净,两端切成平整的新鲜断面,然后把其中一端浸泡于无菌水中,12 ~ 24 h后,在病根的另一端断面会流出乳白色的菌脓。用接种针挑取菌脓划线培养于TTC培养基上。
设计双因素试验,先将病根按无性系分类,再同一无性系不同地点的病根随机混合并分别采用2种方法分离病菌,试验重复3次,每段病根做不少于3个平板,于30℃下恒温培养 48 h。分离率(%)=分离出典型菌落的病根数/参试总病根数×100。
1.3 病原菌16s rRNA测序鉴定
青枯菌在TTC培养基上的典型特征为菌落呈不规则圆形,略隆起,中心部位粉红色,周围有白色晕圈,不透明,具有流动性的。选择具有典型菌落特征的菌株进行16s rRNA测序鉴定。
利用Ezup柱式细菌基因组DNA抽取试剂盒进行待测病菌基因组DNA的提取,试剂盒购买自生工生物工程(上海)股份有限公司。根据 Seal 等[22]结果设计引物,以 16s rRNA 为靶基因的青枯菌特异性引物序列 OL I1:(5’GGGGGTAGCTTGCTACCTG-CC3’)和 Y2:(5’CCCACTGCTGCCTCCCGTAGGA-GT3’)进行扩增。引物由上海生工生物技术有限公司合成。PCR反应体系和反应条件参考王胜坤等[23]的方法。以DL Marker2000为标准分子量,扩增产物经2.0%琼脂糖凝胶电泳,在Bio-Rad凝胶成像系统下观察并记录结果。将扩增产物送至上海生工生物技术有限公司进行测序,将获得的序列经BLAST与GenBank的核酸序列库中已知青枯菌16S rRNA序列进行同源性比较,以判断所分离得到的菌株是否为青枯菌。
1.4 青枯病菌致病性测定
1.4.1 参试无性系及菌株培养 选择抗性不同的木麻黄K18、A14无性系木质化褐梗枝条作为茎段水培接种测定材料。选择6个月龄期、60~70 cm高、根系发达、生长一致的木麻黄K18、A14无性系营养袋苗为小苗盆栽接种试验苗木。所有参试材料均为不带菌健康苗木。
选择经测序鉴定的22个青枯菌菌株作为测定菌株。各菌株在TTC固体培养基上活化48 h,再扩大培养于液体培养基中摇床培养36 h。培养液于5 000 r·min-1离心15 min收集菌体,用无菌水配成浓度3×108cfu·mL-1(平板计数法)菌悬液用于茎段水培接种,2.7×109cfu·mL-1(平板计数法)的菌悬液用于小苗盆栽接种。
1.4.2 室内水培接种法及病情调查 设计2个无性系、22个菌株的双因素交叉试验。采用恒温水培法接种[13]。将木质化褐梗枝条剪成15~20 cm的茎段,每个茎段含有8~10个小枝。每个处理将3~4段褐梗茎段浸入盛有200 ml细菌悬浮液的玻璃瓶内,每瓶含有20~30个小枝。每个处理重复3次,无菌水作对照,置于温度30℃,相对湿度80%,光照时间16 h,强度为8 000 Lux 的人工气候箱中培养,连续调查5 d,每天观察记录植株发病情况,第5天数据进行统计分析。
抗性水平的划分方法参考相对病害强度(RDI)[5]及Yamazaki[24]的分级标准,将侵染后的木麻黄小枝分为4个等级(0级,分枝无症状;1级,分枝萎蔫下垂,保持绿色;2级,分枝枯黄、萎蔫下垂;3级,小枝干枯死亡)进行病情分级,计算病情指数。病情指数=(∑(各级病级分枝数×相对级数值)/(调查总分枝数×发病最高级数值))×100。
1.4.3 小苗盆栽接种法及病情调查 设计2个无性系、22个菌株的双因素交叉试验。参考移栽浸根法[25]进行青枯菌致病性检测。抖落苗木土壤,洗净根部,除去基部黄化叶片,剪去1/3根系,浸入配制好的细菌悬浮液中,30~31℃保湿浸根培养30 min后种植于装有草木灰与黄心土(体积比为1:2)的塑料盆中。草木灰与黄心土提前3 d装好,消毒,搅拌均匀。每个处理一盆,每盆种植25~30株,重复3次,无菌水作对照。种植后每天浇水保持盆内湿润,昼夜温度为28~35℃,相对湿度80%以上。每天观察记录植株发病情况,第9天数据进行统计分析。
以株为单位调查,记录无病植株数与死亡植株数,计算死亡率。死亡率(%)= 死亡植株数 / 总株数×100。
1.5 统计分析
试验数据表示为平均值±标准差(SD),采用SAS V8.1统计软件对试验结果进行方差分析,相关分析及Duncan多重比较检验不同平均值间的差异等。
2 结果与分析
2.1 两种分离方法的比较分析
采用稀释分离法及根系溢出法均能较好地从木麻黄根系分离出青枯病菌。稀释分离法分离率较高,90%以上的病根可以分离出青枯菌典型菌落,根系溢出法只有60%左右的病根溢出菌脓并分离出典型菌落(表2)。但根系溢出法直接从溢出的菌脓上挑菌划线培养,操作更加简便,杂菌含量低,通常不需要二次分离即可获得较纯的菌株。方差分析表明,无性系及无性系与分离方法的交互作用对分离率影响不显著,分离率的高低主要是由分离方法不同引起的(表3)。

表2 两种分离方法的比较
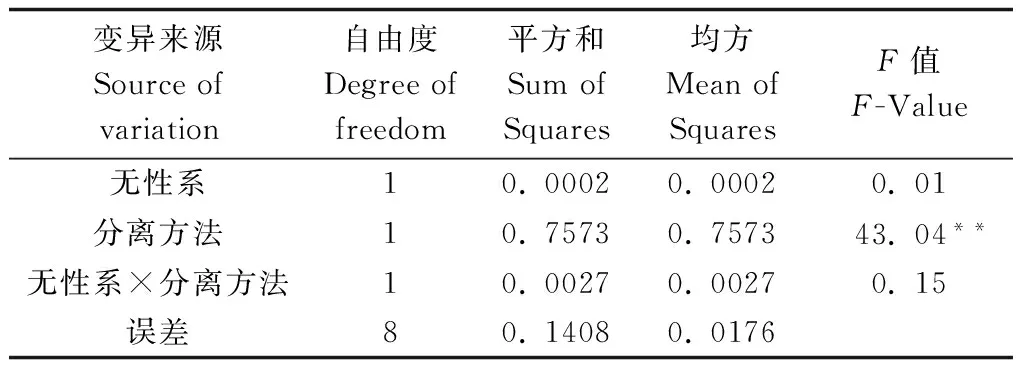
表3 不同分离方法方差分析
注:**示0.01极显著水平。
本试验利用两种分离方法共分离出了31个在TTC培养基上具有典型菌落特征的病原菌菌株。分离获得的31个菌株具体情况见表1。
2.2 病原菌16s rRNA测序鉴定
以 OLI1/Y2 为特异性引物扩增 16S rRNA 基因序列,分离出的31个菌株有22个可在2 h内扩增出280 bp左右的目标条带,且无杂带出现,电泳结果如图1。对这22个病原菌作进一步测序鉴定,而A、C、E、G、J、L、T、W、AC等9个菌株未扩增出特异性条带,为阴性,不再做进一步测序鉴定。

图1 部分菌株16S rRNA序列扩增电泳检测结果Fig.1 Amplified electrophoresis detection results for 16S rRNA sequence of some strains
将22个菌株16S rRNA正反向测序结果进行拼接后与 GenBank 核酸数据库的基因序列进行比对,结果表明:B、D、F、H、I、K、M、N、O、P、Q、R、S、U、V、X、Y、Z、AB、AD、GL-2及TC-1菌株与数据库中Ralstonia solanacearum 菌株(登录号为CP012943.1、CP012939.1)16S rRNA的同源性高达100%,结合菌落培养特征,确认这些致病菌为青枯菌(R. solanacearum)。利用稀释分离法与根系溢出法分离出的同一个菌株在形态,特异性条带及同源性比较中没有差异。其中M菌株16S rRNA与GenBank中一种青枯菌16S rRNA序列的比较见图2。
2.3 菌株致病性测定
2.3.1 室内水培接种 由表4可见,参试菌株对K18、A14无性系褐化茎段均具有致病性,病情指数在20.0 ~ 94.4之间。22个菌株接种K18茎段后致病性差异极显著(P<0.01),利用 Duncan法进行多重比较,将菌株致病性分成若干等级,AD、Y菌株致病性最强,病情指数分别为94.4和90.3;致病性最弱的菌株是N、R和U,病情指数分别为41.9、43.6、45.4。22个菌株接种A14茎段后致病性差异极显著(P<0.01),利用 Duncan法进行多重比较,TC-1、GL-2、D菌株致病性最强,病情指数分别为65.6、64.1和61.2;致病性最弱的菌株是I、O、S及V,病情指数低于25。虽然在不同无性系中菌株的致病性强弱排列次序有一定差别,如V、Y、Z菌株在A14无性系中致病性等级下降,TC-1、Q、D、AB菌株在A14无性系中致病性等级提高,但两次测定结果也指示出一定的规律性,菌株AD、Y、GL-2、H、M、TC-1、F、Q、D在两个无性系中致病性均较强,而菌株N、I、U、S、O、R致病性较弱。
双因素方差分析结果表明:无性系、菌株及无性系×菌系的F检验都是极显著的(P<0.01),说明茎段水培接种中不同菌株和不同无性系对木麻黄的发病程度有极显著影响,菌株与无性系的交互作用对病情指数也存在极显著影响(表5)。

图2 M-16S rRNA序列与GenBank中一种青枯菌16S rRNA序列的比较Fig.2 Comparison of rRNA sequence of M-16S and one of the Ralstonia solanacearum in GenBank

菌株Strain茎段水培接种WaterplantinginoculationK18病情指数Diseaseindexa=0.01A14病情指数Diseaseindexa=0.01小苗盆栽接种PottinginoculationK18死亡率mortality/%a=0.01A14死亡率mortality/%a=0.01B66.2±9.7bcd45.9±11.0abcd56.0±16.9de33.9±15.1bcdefgD64.1±20.6bcd61.2±14.8ab4.9±2.2ijk0.0±0.0hF83.2±5.6abc49.6±8.6abcd90.4±8.6ab56.2±14.2abcH80.6±12.4abc59.3±14.7abc93.3±6.1a65.2±19.2abI50.1±19.7cd20.0±5.2f0.0±0.0k0.0±0.0hK51.4±17.0cd41.2±9.2bcde26.0±5.2fgh47.6±8.2abcdefM83.7±7.6abc56.1±3.8abc84.8±6.1abc69.2±19.6aN41.9±8.2d32.9±3.5def1.9±3.2jk0.0±0.0hO58.5±10.7cd21.2±2.4ef6.5±3.0hijk0.0±0.0hP62.9±10.8cd40.9±4.1bcdef24.2±10.7ghi23.9±13.9defgQ70.6±3.6bcd56.2±7.7abc89.2±4.5abc49.7±15.4abcdeR43.6±12.5d41.5±12.0bcde19.3±6.4ghi38.7±26.0abcdefgS56.9±24.7cd21.1±4.9ef0.0±0.0k0.0±0.0hU45.4±2.6d30.8±2.7def25.4±4.1fgh16.0±4.0gV78.7±18.7abc22.2±7.1ef41.8±31.0efg18.8±14.0fgX67.7±3.5bcd44.1±8.2abcd64.6±19.3cde31.1±10.2cdefgY90.3±11.7ab49.0±14.5abcd40.8±9.3efg21.5±7.1efgZ72.8±19.5bcd37.5±8.7cdef13.5±5.1hij18.9±8.2efgAB60.6±19.3cd51.6±18.3abcd74.1±15.7bcd20.1±13.0efgAD94.4±9.6a56.3±11.3abc54.1±10.3def32.0±8.2cdefgGL-281.4±9.6abc64.1±14.2a92.2±3.4ab54.5±15.8abcdTC-172.0±19.2bcd65.6±13.5a89.8±5.8ab59.7±29.1abc
注:所标字母相同表示差异不显著。
2.3.2 小苗盆栽接种 由表4可见, 22个菌株接种K18小苗后致病性差异极显著(P<0.01),死亡率在4.9% ~ 93.3%之间,利用 Duncan法进行多重比较,将菌株致病性分成若干等级,H、F、GL-2菌株致病性最强,造成小苗死亡率90%以上;菌株I、S对K18无性系无致病性,接种2 d后出现轻微萎蔫症状,这主要是由于伤根移栽引起的正常生理现象,培养4 d后植株恢复生长,叶色健康浓绿。22个菌株接种A14小苗后致病性差异极显著(P<0.01),利用 Duncan法进行多重比较,将菌株致病性分成若干等级,H、M菌株致病性最强,造成小苗死亡率分别为65.2%、69.2%;菌株D、I、N、O、S对A14无性系无致病性。综合比较分析GL-2、H、M、TC-1、F、Q、AB、X、B在两个无性系中致病性均较强,而菌株N、I、S、O、D、U、P致病性较弱。
双因素方差分析结果表明,菌株致病性在无性系间、菌株间及菌株与无性系间的交互作用均具有极显著差异(P<0.01),表明小苗盆栽接种中不同无性系、不同菌株及菌株与无性系的交互作用对菌株致病性具有极显著的影响(表5)。

表5 菌株致病性方差分析
注:**示0.01极显著水平;*示0.05显著水平。
2.3.3 不同接种方法对菌株致病性的影响 研究发现N、I、S、O、D菌株对A14、 K18无性系小苗无致病性或弱致病性(死亡率0% ~ 6.5%),但这些菌株却能使两个无性系褐化茎段迅速致病,病情指数在20.0 ~ 64.1之间(表4)。茎段水培接种与小苗盆栽接种相关分析结表明(表6),不同接种方法之间相关系数值较小,介于0.496 6 ~ 0.754 0之间,表明不同接种方法间菌株致病性线性相关程度为中度相关,即不存在密切的直线相关关系,在茎段水培接种表现强致病性的菌株在小苗盆栽接种中不一定表现出强致病性,反之亦然。

表6 不同接种方法菌株致病性相关分析
注:A:K18无性系茎段水培接种;B:A14无性系茎段水培接种;C:K18无性系小苗盆栽接种;D:A14无性系小苗盆栽接种。*:显著相关;**:极显著相关。
经比较分析,综合选择在不同无性系及不同接种方法中均具有较强致病性的GL-2、H、M、TC-1、F、Q菌株作为下一步木麻黄种质资源抗性鉴定及抗病育种研究试验菌株。
3 讨论
(1)不同方法分离出的同一菌株在形态、鉴定及致病性等方面没有差异。与传统稀释分离法相比,采用根系溢出法也能较好地分离出青枯菌菌株,大大减少实验操作,简便快速,杂菌含量少,但分离成功率略低,主要原因可能是部分木麻黄根系组织材质致密,不利于菌脓的溢出,二是植株感病时间太短而没有足够的菌脓流出。
(2)R. solanacearum 的核糖体基因序列(即 16S rRNA 间隔区)具有高度保守性和特异性,在同一物种的相似度可高达 98%[22, 26-27],已成为病原细菌检测和鉴定的常用方法[28],本研究首次将这种技术应用于木麻黄青枯菌的检测鉴定,利用TTC培养基法[29]分离出的31个病株有22个病株扩增出了特异性条带,经测序比对确定这22个菌株为青枯菌,青枯菌检出率约为71% 。
(3)茎段水培接种试验时,将侵染小枝症状分为无症状、分枝萎蔫下垂但保持绿色、分枝枯黄并萎蔫下垂、干枯死亡4个等级,这种病情分级方法考虑到了感病植株的发病进程,比王军[5]简单将小枝分级为萎蔫与正常2个等级更科学合理,结论更准确可靠。小苗盆栽接种试验时,由于伤过根,前3天小苗会有不同程度的萎蔫下垂甚至基部小枝枯黄脱落,4天后未侵染的苗木逐渐恢复生长,受侵染的小苗则逐渐枯萎死亡。由于小枝萎蔫下垂与脱落伴随着未侵染的苗木较长的时间,作者认为小苗盆栽试验以最终苗木死亡率作为测定指标比王军[12]用萎蔫下垂小枝数的病情判定方法可靠、准确。
(4)小苗盆栽接种试验中,D、I、N、O、S菌株对A14、K18无性系小苗无致病性或弱致病性(死亡率0% ~ 6.5%),但这些菌株却能使两个无性系褐化茎段迅速致病,病情指数在20.0~ 64.1之间,表明这些菌株不易通过根系侵染植株,但却能直接侵入茎干维管束组织使植株迅速致病。这是可能由于青枯菌从茎段侵入时,可直接粘附于维管束组织横断面,随着小枝的蒸腾作用快速侵入茎段维管束组织,大量繁殖并迅速扩散至整个植株,迅速引起病害[30];而从根部侵入时,致病性弱的青枯病菌则被幼根细胞周围的浓密物质所包围,而阻止病菌的活动,从而抑制病菌在根部增殖[31],Vasse[32]研究也发现致病菌能侵入到根部内皮层和维管束中,非致病菌能侵入到内皮层,但在维管束中未发现病菌。从本次试验看,能通过茎段侵染而不易通过根系侵染的菌株占了参试菌株的23%。木麻黄青枯菌的这种侵染特性部分解释了广东省木麻黄青枯病大爆发经常发生在台风过后的现象。台风过后,一方面根系受到伤害,部分菌株可通过根系侵染植株;另一方面大量小枝或茎干折断、破裂,茎干维管束组织暴露出来,那些不易通过根系侵染的菌株在风雨作用下侵入茎干维管束组织使林木迅速发病,引起沿海木麻黄青枯病大爆发。这些不易通过木麻黄根系侵染的菌株与容易通过根系侵染的菌株在生理及致病机理上的差异还有待进一步研究。
(5)木麻黄青枯菌菌株的致病性在不同无性系间、不同菌株间存在极显著差异,而且菌株与无性系之间也存在着极显著的交互作用,即二者的交互作用对寄主的发病程度有直接影响,此结论与王军[13]提出的木麻黄对青枯菌存在水平与垂直抗性及王胜坤[33]在桉树青枯菌致病性测定结果相一致,因此在田间生产中运用单一或少数几个木麻黄无性系大面积推广造林具有较高的风险,应选用多个抗病无性系,有效控制青枯病的发生。相关分析表明,茎段水培接种与小苗盆栽接种试验结果不存在密切的直线关系,即茎段水培接种试验中表现出强致病性的菌株在小苗盆栽接种试验中其致病性有可能会降低,反之亦然。因此,木麻黄青枯病致病菌株筛选时,应开展菌株、无性系、接种方法的交叉接种试验,综合选择。
4 结论
通过稀释分离法及根系溢出法对分离出的31个病株进行16s rRNA测序鉴定,经测序比对其中22个菌株为青枯菌。青枯菌株致病性测定结果显示菌株致病性在无性系间、菌株间及菌株与无性系间的交互作用均具有极显著差异,不同接种方法间菌株致病性不存在密切的直线相关关系。综合比较分析,选择在不同无性系、不同接种方法中都具有较强致病性的GL-2、H、M、TC-1、F、Q等菌株作为下一步木麻黄种质资源抗性鉴定及抗病育种试验菌株。
[1] Salanoubat M,Genin S,Artiguenave F,etal. Genome sequence of the plant pathogen Ralstonia solanacearum[J]. Nature,2002,415:497-502.
[2] 乔俊卿,陈志谊,刘邮洲,等. 茄科作物青枯病研究进展[J]. 植物病理学报,2013,43(1):1-10.
[3] 孙 思,伍慧雄,王 军. 木麻黄青枯病研究概述[J]. 中国森林病虫,2013,32(5):29-34.
[4] 郭 权,梁子超. 木麻黄抗青枯病品系的筛选技术和综合防治措施[J]. 林业科技通讯,1986(4):7-9.
[5] 王 军. 影响木麻黄青枯病抗性测定的几项因素研究[J]. 林业科学,1996,32(3):225-229.
[6] 何学友. 木麻黄病害研究概述[J]. 防护林科技. 2007,(2):27-30.
[7] 黄金水,何益良,郑辉棋. 几种木麻黄抗病虫性调查报告[J].福建林业科技,1985,50(2): 41-45.
[8] 魏素梅,谭天泳. 木麻黄地理种源的苗期试验[J]. 林业科学研究,1990,3(2):119-126.
[9] 梁子超,陈小华. 木麻黄抗青枯病品系的筛选[J]. 华南农学院学报,1984,5(1) : 53-59.
[10] 彭国强. 木麻黄抗青枯病无性系造林对比试验[J]. 广东林业科技,2000,16(3):35-37.
[11] 柯玉铸,黄金水,林延生,等. 普通木麻黄抗逆无性系的筛选[J]. 福建林业科技,1994,21(1):39-43.
[12] 苗立祥. 番茄抗青枯病的AFLP分子标记及其相关基因的克隆[D]. 杭州:浙江大学,2008.
[13] 王 军. 木麻黄对青枯菌的水平及垂直抗性研究[J]. 林业科学,1997,33(5):427-431.
[14] Kelman A. The relationship of pathogenicity of Pseudomonas solanacearum to colony appearance in tetrazolium medium[J]. Phytopathology,1954,(44):693-695.
[15] 陈炳铨,张景宁. 木麻黄无性系对青枯菌抗性及菌株变异初探[J]. 广东林业科技,1995,11(2):33-36.
[16] 晏立英,黄家权,雷 永,等. 花生青枯菌红安分离物的鉴定[J]. 中国油料作物学报,2010,32(1):144-146.
[17] 王胜坤,王 军,徐大平. 四种桉树青枯菌DNA提取方法及PCR检测灵敏度比较[J]. 中国森林病虫,2007,26(5):4-7.
[18] 戴凡炜,李 磊,唐翠明,等. 引起广西桑树“褐枯”的病原菌及致病机制研究[J]. 热带作物学报,2013, 34(10):2004-2008.
[19] Stephane P,Jacques L. Specific detection of biovars ofRalstoniasolanacearumin plant tissues by Nested-PCR-RFLP[J] . European Journal of Plant Pathology,2000,106:255-265.
[20] Mitsuo H,Kazutaka Y,Kenichi T. PCR-based specific detection of Ralstonia solanacearum race 4 strains[J] . Journal of Genernal Plant Pathology,2004,70:278-283.
[21] 朱圣杰. 番茄抗青枯病生理生化机制的研究[D]. 合肥:安徽农业大学,2004.
[22] Seal S E, Jackson L A,Young J P,etal. Differentiation ofPseudomonassolanacearum,Pseudomonassyzygii,Pseudomonaspickettiiand blood disease bacterium by partial 16S rRNA sequencing: construction of oligonucleotide primers for sensitive detection by polymerase chain reaction[J]. Microbiology,1993,139 (7):1587-1594.
[23] 王胜坤,王 军,徐大平. 四种桉树青枯菌DNA提取方法及PCR检测灵敏度比较[J]. 中国森林病虫,2007,26(5):4-7.
[24] Yamazaki H. Relation between resistance to bacterial wilt and calcium nutrition in tomato seedlings[J]. Jarq-Japan Agricultural Research Quarterly,2001,35(3):163-169.
[25] 尹贤贵,王小佳,张 斌,等. 我国番茄青枯病及抗病育种研究进展[J]. 云南农业大学学报,2005,20(2):163-167.
[26] Taghavi M,Hayward C,Sly L I,etal. Analysis of the phylogenetic relationships of strains ofBurkholderiasolanacearum,Pseudomonassyzygii, and the blood disease bacterium of banana based on 16S rRNA gene sequences[J]. International Journal of Systematic Bacteriology,1996,46(1):10-15.
[27] Roberts S J,Eden-Green S J,Jones P,etal.Pseudomonassyzygii,sp. nov., the cause of Sumatra disease of cloves[J]. Systematic and Applied Microbiology,1990,13(1):34-43.
[28] 晏立英,黄家权,雷 永,等. 花生青枯菌红安分离物的鉴定[J]. 中国油料作物学报,2010,32(1):144-146.
[29] 董汉松. 植病研究方法[M],北京:中国农业出版社,2012: 90.
[30] Schell M A. Control of virulence and pathogenicity genes ofRalstoniasolanacearumby an elaborate sensory network[J]. Annu Rev Phytopathol,2000,38:263-292.
[31] 王 卉,任欣正. 青枯菌(Pseudomonassolanacearum)在番茄抗、感病品种根部的吸附、侵入和繁殖[J]. 植物病理学报,1993,23(2):143-149.
[32] Vasse J,Frey P,Trigalet A. Microscopic studies of intercellular infection and protoxylem invasion of tomatoroots byPseudomonassolanacearum[J]. Molecular Plant-Microbe Interactions,1995,8(2):241-251.
[33] 王胜坤. 桉树青枯菌菌株致病力分化、吸附识别及PCR快速检测研究[D]. 北京:中国林业科学研究院,2007.
(责任编辑:崔 贝)
Isolation of Pathogen ofCasuarinaBacterial Wilt and Screening of High Pathogenic Strains
XU Xiu-yu1,2, XU Bin2, GANG Xian-hua2, ZHONG Chong-lu3, ZHANG Hua-xin1
(1. Research Centre on Saline and Alkali Lands of State Forestry Administration, Chinese Academy of Forestry, Beijing 100091, China; 2. Guangdong Provincial Key Laboratory of Silviculture, Protection and Utilization, Guangdong Academy of Forestry, Guangzhou 510520, Guangdong, China; 3. Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou 510520, Guangdong, China)
[Objective] To screen high pathogenic strains for studies on germplasm resources resistance identification and resistance breeding ofCasuarina. [Method] TheCasuarinaroots were collected from heavily infected areas in western Guangdong Province. Two methods, dilution isolation and root overflow, were used in strains isolation.Ralstoniasolanacearumstrains were identified by 16S rRNA sequencing and the pathogenicity of these strains were measured. [Result] 31 strains were isolated. The method of root overflow is simple to operate and the isolation rate was about 60%, thus was supplementary to regular dilution method. Only 22 strains were amplified from the 31 strains by using 16S rRNA sequencing, and the amplified strains were identified asR.solanacearum. The results of pathogenicity measurement showed that the pathogenicity of different strains was significantly different (p<0.01), and the correlation coefficients between two different inoculation approaches (water planting and potting) were significant but relatively low (0.496 6 ~ 0.731 0), suggesting that the two approaches were independent. [Conclusion] Six strains were selected for next-step study.
Casuarina;Ralstoniasolanacearum;isolation;molecular identification;strain screening
10.13275/j.cnki.lykxyj.2017.03.007
2016-07-14
广东省林业科技创新项目(2014KJCX017和2013KJCX018-03)
许秀玉,教授级高级工程师,博士研究生,从事林木遗传育种及森林生态研究。E-mail:81250908@163.com
* 通讯作者:张华新,研究员,博士生导师,从事植物抗逆育种研究。E-mail:18611572609@163.com
S432.4+2
A
1001-1498(2017)03-0409-08
猜你喜欢
华南农业大学学报(2022年1期)2022-01-14 05:42:56
中学生博览(2020年23期)2020-12-10 10:56:15
防护林科技(2019年1期)2019-03-13 05:27:50
天然产物研究与开发(2018年3期)2018-05-07 06:38:39
闽南风(2017年12期)2018-01-04 18:13:52
西南农业学报(2016年5期)2016-05-17 05:42:34
现代农业(2015年1期)2015-02-28 18:39:51
植物营养与肥料学报(2012年5期)2012-10-26 03:28:36
中国烟草学报(2012年5期)2012-04-12 06:21:10
中国烟草学报(2012年1期)2012-04-09 07:16:08
