
马蔺重金属ATP酶基因HMA2的RNAi 载体构建及其遗传转化
2017-06-05 14:14张亚楠蔺银鼎毛培春田小霞
草业科学 2017年5期
张亚楠,蔺银鼎,孟 林,毛培春,田小霞,郭 强
(1.山西农业大学林学院,山西 太谷 030801; 2.北京市农林科学院 北京草业与环境研究发展中心,北京 100097)
马蔺重金属ATP酶基因HMA2的RNAi 载体构建及其遗传转化
张亚楠1,2,蔺银鼎1,孟 林2,毛培春2,田小霞2,郭 强2
(1.山西农业大学林学院,山西 太谷 030801; 2.北京市农林科学院 北京草业与环境研究发展中心,北京 100097)
重金属ATP酶基因HMA2介导的Cd长距离运输对植物体内Cd的分配及解毒起重要作用。然而,有关马蔺(Irislactea)体内Cd长距离运输的分子机制仍不清楚。本研究以马蔺根中总RNA为模板,采用RT-PCR方法获得了重金属ATP酶基因片段,以中间载体pHANNIBAL和植物表达载体pART27为基础,采用酶切和连接的方法,构建了CaMV35S启动子驱动的含有IlHMA2基因片段反向重复序列的RNAi植物表达载体pARTG1G2,用冻融法将此重组质粒导入农杆菌GV3101,并转入到马蔺幼苗体内获得了IlHMA2-RNAi转化株系。这将为阐明IlHMA2介导的Cd长距离运输在马蔺适应重金属镉污染环境中的作用机制奠定基础。
马蔺;重金属ATPase酶;镉富集与耐受性;RNAi载体构建
近年来我国重金属污染事件频发,受镉(Cd)、砷(As)、铅(Pb)、汞(Hg)、锌(Zn)等重金属污染的耕地面积达到总耕地面积的1/5,重金属污染与防治已成为不容忽视的环境问题[1]。与其它重金属元素相比,土壤中Cd2+极易被植物根系吸收,导致植物体内积累大量的Cd2+,造成农作物产量和品质下降,更为严重的是,Cd2+可通过食物链进入人体,对人类的健康构成严重威胁[2-4]。因此,如何行之有效地控制和减轻重金属对环境的污染和危害已成为一个亟待解决的问题。
植物修复技术是一项新兴的能源节约型和生态友好型环境修复技术,即利用植物对重金属的吸收能力,将土壤中的重金属富集并转运至植株地上部,以达到降低土壤中重金属含量的目的[5-7]。马蔺(Irislactea)又名马莲,是鸢尾科鸢尾属多年生宿根草本植物,广泛分布于我国东北、西北和华北等地区,其根系发达、叶量丰富,对环境适应性强、长势旺盛、养护成本较低,是节水、抗旱、耐盐碱的优良观赏地被植物。叶片翠绿柔软耐践踏,花淡雅美丽,在生态园林建设中具有良好的应用前景[8-10];尤其是在80 mg·L-1镉处理42 d后,马蔺地上部Cd2+含量为529 μg·g-1(>100 μg·g-1,属于镉超富集植物),占植株体内Cd2+总含量的46%,而且植株仍处于旺盛生长状态,并未出现中毒症状[11],说明马蔺具有强大的Cd长距离运输能力,能够将根系吸收的大量Cd转运至地上部,降低Cd对根系的毒害,从而使其对Cd表现出很强的耐受和富集能力,具备植物修复的潜力。研究表明,重金属ATP酶 (heavy metal ATPase,HMA)广泛存在于细菌、人及高等植物中,其在植物体内主要参与重金属Cu+、Ag+、Zn2+、Cd、Co2+与Pb2+转运,其中HMA2/4在植物体内Cd2+的长距离运输中发挥着重要作用[12-15]。鉴于此,Guo等[10]从马蔺中克隆得到了重金属ATP酶基因IlHMA2,但该基因在马蔺体内Cd2+长距离运输中的作用机制仍不清楚。
RNA干扰技术(RNA interference,RNAi)是基因功能鉴定与基因表达调控研究常用的方法,已广泛的应用于水稻(Oryzasativa)、小麦(Triticumaestivum)等多种作物中[16-17]。Hannon[18]在牵牛花(Pharbitisnil)转基因的研究中首次发现共抑制沉默现象,并认为与甲基化作用有关。Fire等[19]在研究RNA干扰所需的结构和传递条件的试验中发现,双链RNA诱导抑制同源性序列基因表达比任意一个单独的正义链或反义链效果好。RNAi具有高效性、特异性和遗传性,通过抑制靶基因的表达,获得部分功能丧失的突变体,进而研究该基因与细胞发育、分化及功能间的关系[17]。RNAi通过导入与内源性mRNA同源序列的双链RNA进而阻断体内特定基因的表达,促使mRNA降解,进而诱发基因沉默以达到鉴定该基因功能的目的[13,15,19-21],如Shimizu等[22]利用RNAi沉默了水稻矮缩病毒的一个病毒原质基质蛋白基因Pns12,结果获得了病毒抗性的转基因水稻;郭志鸿等[23]用RNAi干扰技术创造高直链淀粉马铃薯(Solanumtuberosum)材料;马建等[24]应用RNAi干扰技术创造低脂肪氧化酶活性大豆(Glycinemax)新种质。本研究以马蔺IlHMA2基因为靶标,采用酶切连接的方法,构建具有反向重复序列的发卡结构的IlHMA2基因-RNAi植物表达载体,旨在为研究IlHMA2基因在马蔺体内Cd长距离运输中作用机制奠定基础。
1 材料与方法
1.1 材料
试验所用植物材料为8周龄的野生马蔺幼苗,种子采自北京市农林科学院小汤山草资源试验基地;根癌农杆菌GV3101、pHANNIBAL载体与pART27载体由北京草业与环境研究发展中心实验室保存。
1.2 主要试剂
TaKaRa MiniBEST Plant RNA Extraction Kit试剂盒、TaKaRa MiniBEST Universal Genomic DNA Extraction Kit试剂盒、PCR扩增试剂盒、SanPrep柱式DNA胶回收试剂盒、TA克隆、PrimeScriptTM1st Strand cDNA Synthesis Kit、质粒提取试剂盒、pMD@19-T Simple载体、大肠杆菌菌株EscherichiacoliDH5α、限制性内切酶及其DNA marker等试剂盒均购自宝生物工程有限公司,引物合成购自上海生物工程有限公司,其它生化试剂均为进口或国产分析纯。
1.3 植物的培养
选取色泽光亮饱满的马蔺种子放于烧杯中,加入5%次氯酸钠溶液中消毒5 min后,再置于40 ℃的水浴锅中浸泡56 h。之后将其均匀播撒在装有营养土和细沙(体积比为2∶1)的小钵中,并放置入人工气候箱培养(如同下述光照培养条件)。长至6周龄时,挑选长势一致的幼苗将其冲洗干净移入水培盒(长19 cm、宽13.5 cm、高7.5 cm)中,浇灌Hoagland营养液[0.7 μmol·L-1(NH4)6Mo7O24·4H2O,2 mmol·L-1KNO3,92 μmol·L-1H3BO3,0.5 mmol·L-1NH4H2PO4,0.6 μmol·L-1CuSO4·5H2O,0.1 mmol·L-1Ca(NO3)2·4H2O,0.5 mmol·L-1Fe-citrate,18 μmol·L-1MnC12·4H2O,0.25 mmol·L-1MgSO4·7H2O,1.6 μmol·L-1ZnSO4·7H2O]继续培养两周,营养液每3 d更换一次。培养室光照培养16 h(昼)25 ℃/8 h(夜)18 ℃,光强约为600 μmol·(m2·s)-1,空气相对湿度为50%~80%[9]。
1.4IlHMA2基因RNAi靶位片段的获得
将培养至8周龄的马蔺幼苗移入含100 mg·L-1CdCl2·2.5H2O的营养液中,进行胁迫处理24 h后,剪其根部经无菌水冲洗后,置于消毒滤纸上迅速吸干水分进行取样,取样量为200 mg,于液氮中充分研磨,使用TaKaRa MiniBEST Plant RNA Extraction Kit试剂盒提取总RNA,利用Prime Script RTase cDNA第一链合成试剂盒进行反转录,合成cDNA。首先通过酶切位点分析,根据RNAi目标序列选取原则,确定siRNA序列,再通过Blast进行同源序列的对比,而后利用OligoEngine 2.0软件对IlHMA2基因(GenBank登录号:KY696282)的靶位区段进行筛选,并结合IlHMA2基因结构特点,最终确定第1 261~1 682 bp处共422 bp为最佳靶片段,然后利用Primer 5.0软件设计两对特异性引物分别为P1和P2、P3和P4,分别在上下游引物处引入酶切位点(XhoⅠ和KpnⅠ、XbaⅠ与和ClaⅠ)以便连入载体pHANNIBAL。
P1:5′-CCCTCGAGTCTGTCCTATTGGCTGCC-3′;
P2:5′-GGGGTACCAGTGCTATCTCCAGTAAG-3′;
P3:5′-GCTCTAGATCTGTCCTATTGGCTGCC-3′;
P4:5′-CCATCGATAGTGCTATCTCCAGTAAG-3′.
PCR扩增反应体系:在50 μL PCR管中依次加入2×Gflex PCR Buffer (Mg2+,dNTP plus)25 μL、Tks Gflex DNA Polymerase 1 μL、P1和P2(或P3和P4)各1 μL、cDNA 4 μL,加水补至50 μL。反应条件:94 ℃ 1 min;98 ℃ 10 s,55 ℃ 15 s,68 ℃ 30 s,30个循环;最后72 ℃延伸10 min。将PCR扩增产物用1%琼脂糖凝胶进行检测,并使用宝生物DNA胶回收试剂盒对其进行回收和纯化。随后利用宝生物Mighty TA-cloning Reagent Set试剂盒对回收得到的PCR纯化产物3′平末端加A,而后立即将加A产物连接到pMD@19-T Simple载体,转化到感受态大肠杆菌DH5α中,将其涂布于含有50 mg·L-1氨苄青霉素的LB固体培养基37 ℃黑暗倒置培养,次日进行阳性克隆蓝白斑筛选,将转化后白斑菌株进行质粒PCR鉴定确认阳性克隆后,编号送去测序。
1.5 RNAi表达载体构建
用XhoⅠ与KpnⅠ双酶切pMD@19-T Simple载体并回收正向小片段G1,与用同样双酶切的pHANNIBAL载体连接,重组子转化感受态细胞DH5α,获得重组质粒pHG1。然后,再用XbaⅠ与ClaⅠ双酶切pMD@19-T Simple载体并回收反向小片段G2,与用同样双酶切的pHG1载体连接,重组子转化感受态细胞DH5α,获得具有CaMV35S-G1-PDK intron-G2-OCS terminator发卡反向重复序列的重组质粒pHG1G2(图1)。
用SacⅠ/SpeⅠ双酶切表达载体pART27并回收大片段,与用同样双酶切的重组质粒pHG1G2连接,重组子转化感受态细胞DH5α,获得具有片段G正反向重复序列的植物RNAi表达载体pARTG(图2)。
1.6 农杆菌转化
用冻融法将构建好的表达载体转化到农杆菌GV3101感受态中,将转化的菌液均匀涂布于LB固体培养基上(含50 mg·L-1卡那霉素、50 mg·L-1壮观霉素和50 mg·L-1利福平),28 ℃避光倒置培养,2~3 d后长出单菌落,用接种环挑取单菌落,接种于10 mL含50 mg·L-1卡那霉素、50 mg·L-1壮观霉素和50 mg·L-1利福平的液体LB培养基中,振荡过夜培养。而后提取质粒,用特异引物P5和P6扩增内含子intron基因片段,长度473 bp。
P5:5′-TATGACAAGTGATGTGTAAGACG-3′;
P6:5′-AATAAGAATGTTGATTGAAAAT-3′.
PCR扩增反应体系:在50 μL PCR管中依次加入2×Gflex PCR Buffer (Mg2+,dNTP plus) 25 μL、Tks Gflex DNA Polymerase 1 μL、P5和P6各1 μL、质粒2 μL,加水补至50 μL。反应条件:94 ℃ 1 min;98 ℃ 10s,55 ℃ 15 s,68 ℃ 45 s,25个循环;最后72 ℃延伸10 min。确认阳性克隆后,扩大培养保存在-80 ℃冰箱备用。

图1 中间载体pHANNIBAL-G1G2的构建图
1.7 马蔺幼苗遗传转化
参考Ma等[25]的方法和Weeks等[26],略有修改。具体如下:在上述Hoagland营养液中培养3周龄大小的马蔺幼苗,在其根茎结合点2~3 cm处去除地上部,然后用注射器针头将携带pARTG1G2质粒的根癌农杆菌GV3101菌液(OD=0.4~0.5)注射至伤口处,并用无菌脱脂棉轻轻包裹住伤口,置于黑暗中培养;期间,每隔45 min用其菌液打湿脱脂棉一次,使得其脱脂棉在3 h内保持湿润,4 d后去除脱脂棉;大约两周后马蔺幼苗伤口处长出嫩叶,转入至Hoagland营养液中继续培养4周后即可用于检测转基因植株T0,共转化300株幼苗。野生型株系(WT)采用上述方法用空载pART27根癌农杆菌GV3101侵染。而后,使用TaKaRa MiniBEST Universal Genomic DNA Extraction Kit试剂盒提取WT和T0植株叶片的总DNA,用特异引物P5和P6扩增内含子intron基因片段,长度473 bp。PCR扩增反应体系如上所述。

图2表达载体pART27-G1G2构建图
2 结果与分析
2.1IlHMA2基因RNAi靶位片段的选择及扩增
以马蔺根系总RNA cDNA为模板,用设计的特异性引物P1和P2、P3和P4分别进行PCR扩增,并对其PCR产物进行凝胶电泳检测,发现在400 bp左右处有一条亮带,且上下无杂带,与目的片段的大小一致。将其进行胶回收纯化后,连接到pMD@19-T Simple克隆载体上,并转化到DH5α中。从转化的平板上随机挑取多个阳性克隆进行编号处理,从中任选3个质粒进行提取,通过PCR检测,显示得到的扩增片段大小约在400 bp处(图3),送去测序得到的结果与克隆到的HMA2 cDNA片段比对同源性为100%,表明已成功克隆到目的靶位片段。
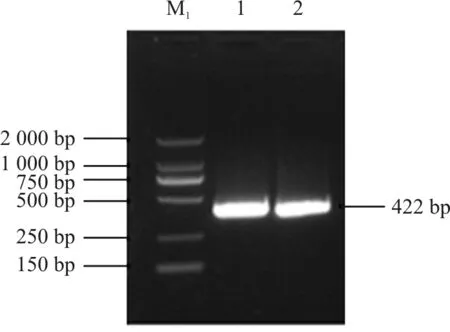
图3 IlHMA2基因片段的扩增产物
2.2 以IlHMA2基因为靶标的RNAi载体构建
通过酶切连接的方法,构建了具有启动子CaMV35S的IlHMA2反向重复序列的植物RNAi表达载体pARTG。根据构建的pARTG载体上的限制性酶切位点分别对其进行酶切分析鉴定。经XhoⅠ/KpnⅠ和KpnⅠ/ClaⅠ双酶切后,在422和800 bp处出现目的条带(图4,泳道1和泳道2),分别与正向片段G1和PDK intron片段大小相符;经XbaⅠ/ClaⅠ双酶切后,在422 bp处也出现了目的条带(图4,泳道3),与反向片段G2相符;经XhoⅠ/ClaⅠ双酶切后,在大约1 700 bp处出现特异条带(图4,泳道4),与G1-PDK intron-G2片段大小相符;最后经SacⅠ/SpeⅠ双酶切后,在约4 500 bp处出现目的条带(图4,泳道5),与CaMV35S-G1-PDK intron-G2-OCS termina-tor片段大小相符。以上结果表明,IlHMA2基因RNAi植物表达载体IlHMA2-RNAi构建成功。

图4 马蔺HMA2-RNAi载体的双酶切验证
2.3农杆菌转化及其IlHMA2-RNAi株系分子鉴定
用冻融法将重组质粒pARTG1G2导入根癌农杆菌GV3101中,挑取单克隆,随后从这些阳性克隆编号中任选3个进行质粒提取,通过特异引物P5和P6进行PCR扩增,凝胶泳检测出473 bp目的片段,证实重组表达载体已成功导入农杆菌GV3101中(图5)。

图5 农杆菌转化阳性克隆PCR检测
提取上述WT和T0叶片的总DNA,用intron基因特异引物对它们进行PCR扩增,扩增得到473 bp片段的仅有15株T0代株系,转化率为5%(图6),这为后续验证马蔺IlHMA2基因的功能奠定了基础。

图6 马蔺IlHMA2-RNAi转化株系的PCR检测
3 讨论与结论
在RNA干扰机制中,分为起始、效应和信号扩增3个阶段。通常只需少量siRNA就可以使得特定的基因发生沉默[27-29]。这种沉默效应可在植物的胞间连丝与韧皮部进行扩散,通过嫁接将沉默效应在砧木和接穗之间双向传递,甚至在不同的物种个体间也可传递。RNA靶区的扩散是从起始siRNA同源区扩散向临近的5′和3′端[30]。构建高效表达载体是进行植物遗传转化的前提,必须在适当的酶切位点连接上目的基因,而强启动子下游插入反向重复片段是最有效的双链RNA的表达载体构件[32-33]。本研究中PCR扩增的HMA2 cDNA片段分别以正向和反向插入双元植物表达载体,由强启动子驱动,正反片段由内含子intron序列隔开以维持载体的稳定性;该系统在植物细胞中转录后可形成带有一个30~50 bp环的ihpRNA,高水平转录形成双链结构引发RNAi,从而抑制HMA2的翻译。沉默的效果远远高于反义的或共抑制结构[33-34]。
选取目标基因的结构序列以及位置,载体结构不同,得到的dsRNA所产生的效应存在着显著差异[33]。通常针对靶基因设计反向重复序列,构建成含发夹结构的双链hpRNA或含内含子的ihpRNA表达载体[35],然后利用农杆菌介导转化,通读反向重复序列转录形成dsRNA,进而诱导RNAi的产生。Helliwell等[36]发现ihpRNA表达载体比hpRNA沉默效率高,且其发夹结构臂序列的长度在100~1 200 bp,具有一定的可塑性;靶基因序列还应在基因转录起始位点下游100个核苷酸以后,且GC含量在40%~52%内[37],这样dsRNA才能够高效、特异地发挥基因沉默作用[38-40]。本研究中构建的RNAi载体的两个反向重复序列臂间具有内含子,且两臂长度也在100~1 200 bp,选取的IlHMA2基因靶基因序列GC含量为42%。因此,理论上构建的RNAi干扰载体能够有效沉默IlHMA2基因。
目前,根癌农杆菌介导法是一种技术较为完善且有效的转基因方法。但由于单子叶植物不是根癌农杆菌的天然宿主,因此阻碍了根癌农杆菌介导法在单子叶植物遗传转化研究方面的应用。随着人们对农杆菌介导法的转化机理、影响因素不断探索与研究,使得根癌农杆菌介导法被逐步应用于单子叶植物中,尤其是禾本科作物[41-42]。选择合适的农杆菌菌株与高效的表达载体是农杆菌转化中至关重要的因素。而对于不同的外植体其细胞生长状态、生活时期、生理代谢及基因表达都存在一定程度上的差异,因而不同生理状态下的细胞对农杆菌的敏感性及农杆菌对该受体植物的感染性都不同。本研究,借鉴Ma等[25]和Weeks等[26]的RNAi农杆菌介导转化方法,用农杆菌侵染马蔺幼苗的根茎结合位点伤口处,以诱导转化得到了马蔺IlHMA2-RNAi转化株系,说明选择适合的RNAi转化的方法,可有效地提高农杆菌侵染单子叶植物的转化效率。以上这些结果,将为解析IlHMA2介导的Cd长距离运输在马蔺适应镉污染环境中的作用机制奠定基础。
References:
[1] 黄益宗,郝晓伟,雷鸣,铁柏清.重金属污染土壤修复技术及其修复实践.农业环境科学学报,2013,3(3):409-417. Huang Y Z,Hao X W,Lei M,Tie B Q.The remediation technology and remediation practice of heavy metals-contaminated soil.Journal of Agro-Environment Science,2013,3(3):409-417.(in Chinese)
[2] Clemens S.Toxic metal accumulation,responses to exposure and mechanisms of tolerance in plants.Biochimie,2006,88(11):1707-1719.
[3] Ueno D,Koyama E,Yamaji N,Ma J F.Physiological,genetic,and molecular characterization of a high-Cd-accumulating rice cultivar,Jarjan.Journal of Experimental Botany,2011,62(7):2265-2272.
[4] Guo Q,Meng L,Mao P C,Tian X X.An assessment ofAgropyroncristatumtolerance to cadmium contaminated soil.Biologia Plantarum,2014,58(1):174-178.
[5] Salt D E,Prince R C,Pickering I J,Raskin I.Mechanisms of cadmium mobility and accumulation inIndianmustard.Plant Physiology,1995,109(4):1427-1433.
[6] 王欣.苎麻镉耐性机制及应用研究.长沙:湖南大学博士学位论文,2010. Wang X.Study ofNovelbioassaysbased on functionalized nanoparticles and bionic materials.PhD Thesis.Changsha:Hunan University,2010.(in Chinese)
[7] Nouet C,Motte P,Hanikenne M.Chloroplastic and mitochondrial metal homeostasis.Trends in Plant Science,2011,16(7):395-404.
[8] 陈默君,贾慎修.中国饲用植物.北京:中国农业出版社,2002:1116-1117. Chen M J,Jia Z X.Forage plants in China.Beijing:China Agriculture Press,2002:1116-1117.(in Chinese)
[9] 郭强,孟林,李杉杉,张琳,毛培春,田小霞.马蔺NHX基因的克隆与基因表达分析.植物生理学报,2015,51(11):2006-2012. Guo Q,Meng L,Li S S,Zhang L,Mao P C,Tian X X.Cloning ofIrislacteaNHXand analysis of gene expression.Plant Physiology Journal,2015,51(11):2006-2012.(in Chinese)
[10] Guo Q,Meng L,Zhang Y N,Mao P C,Tian X X,Li S S,Zhang L.Antioxidative systems,metal ion homeostasis and cadmium distribution inIrislacteaexposed to cadmium stress.Ecotoxicology and Environmental Safety,2017,139:50-55.
[11] Han Y L,Yuan H Y,Huang S Z,Guo Z,Xia B,Gu J G.Cadmium tolerance and accumulation by two species ofIris.Ecotoxicology,2007,16(8):557-563.
[12] Axelsen K B,Palmgren M G.Inventory of the superfamily of P-Type ion pumps inArabidopsis.Plant Physiology,1998,126(2):696-706.
[13] Hanikenne M,Nouet C.Metal hyperaccumulation and hypertolerance:A model for plant evolutionary genomics.Current Opinion in Plant Biology,2011,14(3):252-259.
[14] Guo Q,Meng L,Humphreys M W,Scullion J,Mur L A.Expression ofFlHMA3,a P1B2-ATPase fromFestuloliumloliaceum,correlates with response to cadmium stress.Plant Physiology and Biochemistry,2017,112:270-277.
[15] Williams L E,Mills R F.P(1B)-ATPases——An ancient family of transition metal pumps with diverse functions in plants.Trends in Plant Science,2005,10(10):491-502.
[16] 崔喜艳,孙小杰,刘忠野,张继伟.RNAi机制及在植物中应用的研究概述.吉林农业大学学报,2013,35(2):160-166. Cui X Y,Sun X J,Liu Z Y,Zhang J W.A research review of RNA interference mechanisms and its applications in plants.Journal of Jilin Agricultural University,2013,35(2):160-166.(in Chinese)
[17] 张森浩,严学兵,王成章,文开新,许来俊.RNA干扰及其在植物中的研究进展.草业科学,2011,85(5):823-830. Zhang S H,Yan X B,Wang C Z,Wen K X,Xu L J.RNA interference and its research proceedings in plants.Pratacultural Science,2011,85(5):823-830.(in Chinese)
[18] Hannon G J.RNA interference.Nature,2002,418:244-251.
[19] Fire A,Xu S,Montgomery M K,Kostas S A,Driver S E,Mello C C.Potent and specific genetic interference by double-stranded RNA inCaenorhabditiselegans.Nature,1998,391:806-811.
[20] Axelsen K B,Palmgren M G.Evolution of substrate specificities in the P-type ATPase superfamily.Journal of Molecular Evolution,1998,46(1):84-101.
[21] 言普,沈文涛,黎小瑛,周鹏.植物hpRNA干扰载体构建的研究进展.生物技术通报,2013,9:7-12. Yan P,Shen W T,Li X Y,Zhou P.Progress in construction of hpRNA ector for plant RNAi.Biotechnology Bulletin,2013,9:7-12.(in Chinese)
[22] Shimizu T,Yoshii M,Wei T Y,Hirochika H,Omura T.Silencing by RNAi of the gene for Pns12,a viroplasm matrix protein of Rice dwarf virus, results in strong resistance of transgenic rice plants to the virus.Plant Biotechnology Journal,2009,7(1):24-32.
[23] 郭志鸿,张金文,王蒂,陈正华.用RNA干扰技术创造高直链淀粉马铃薯材料.中国农业科学,2008,41(2):494-501. Guo Z H,Zhang J W,Wang D,Chen Z H.Using RNAi technology to produce high-amylose potato plants.Scientia Agricultura Sinica,2008,41(2):494-501.(in Chinese)
[24] 马建,张君,曲静,王云鹏,魏益凡,王丕武.应用RNA干扰技术创造低脂肪氧化酶活性大豆新种质.中国农业科学,2009,42(11):3804-3811. Ma J,Zhang J,Qu J,Wang Y P,Wei Y F,Wang P W.Development of novel soybean germplasms with low activity of lipoxygenases by RNAi method.Scientia Agricultura Sinica,2009,42(11):3804-3811.(in Chinese)
[25] Ma Q,Li Y X,Yuan H J,Hu J,Wei L,Bao A K,Zhang J L,Wang S M.ZxSOS1 is essential for long-distance transport and spatial distribution of Na+and K+in the xerophytesZygophyllumxanthoxylum.Plant and Soil,2014,374(1):661-676.
[26] Weeks J T,Ye J,Rommens C M.Development of an in planta method for transformation of alfalfa (Medicagosativa).Transgenic Research,2008,17(4):587-597.
[27] Meister G,Tuschl T.Mechanisms of gene silencing by double-stranded RNA.Nature,2004,431:343-349.
[28] Baulcombe D.RNA silencing in plants.Nature,2004,431:356-363.
[29] Mallory A C,Mlotshwa S,Bowman L H,Vance V B.The capacity of transgenic tobacco to send a systemic RNA silencing signal depends on the nature of the inducing transgene locus.The Plant Journal,2003,35(1):82-92.
[30] 白描,杨国顺,陈石,张美玲.植物RNAi的特点及其应用研究进展.生物技术通报,2009(8):6-10. Bai M,Yang G S,Chen S,Zhang M L.Progress of the RNAi characteristics and its application in plants.Biotechnology Bulletin,2009(8):6-10.(in Chinese)
[31] 马建,魏益凡,厉志,王丕武.植物RNA干扰表达载体构建方法的研究.安徽农业科学,2009,37(18):8364-8366. Ma J,Wei Y F,Li Z,Wang P W.Study on the construction method of plant RNAi expression vector.Journal of Anhui Agricultural Sciences,2009,37(18):8364-8366.(in Chinese)
[32] 许德晖,黄辰,刘利英,宋土生.高效siRNA设计的研究进展.遗传,2006,28(11):1457-1461. Xu D H,Huang C,Liu L Y,Song T S.New progress of the highly efficient siRNA design.Hereditas,2006,28(11):1457-1461.(in Chinese)
[33] 张付云,李妍,李云冰,张瑛,赵小明,杜昱光.NtSKP1基因的RNAi载体构建及对烟草的转化.西北农业学报,2010,19(8):95-100. Zhang F Y,Li Y,Li Y B,Zhang Y,Zhao X M,Du Y G.Construction of RNAi expression vector ofNtSKP1 and transformation in tobacco.Acta Agriculturae Boreali-Occidentalis Sinica,2010,19(8):95-100.(in Chinese)
[34] 徐博,任伟,王英哲,孙启忠,郭玮,娄玉杰.农杆菌介导的朝鲜碱茅PuP5CS基因转化紫花苜蓿的研究.草业科学,2015,32(6):895-901. Xu B,Ren W,Wang Y Z,Sun Q Z,Guo W,Lou Y J.Transformation ofPuccinelliachinampoensisPuP5CSgene into alfalfa withAgrobacterium-mediated method.Pratacultural Science,2015,32(6):895-901.(in Chinese)
[35] 段丽婕,王沛,陈梦词,王锁民.盐生植物小花碱茅外整流K+通道PtSKOR基因的克隆及RNAi载体构建.分子植物育种,2015,13(4):877-886. Duan L J,Wang P,Chen M C,Wang S M.Cloning outward-rectifying potassium channelPtSKORgene and constructing its RNAi vector in halophytePuccinelliatenuiflora.Molecular Plant Breeding,2015,13(4):877-886.(in Chinese)
[36] Helliwell C,Waterhouse P.Constructs and methods for high-throughput gene silencing in plants.Methods,2003,30(4):289-295.
[37] 马清,王锁民.多浆旱生植物霸王质膜Na+/H+逆向转运蛋白基因RNAi载体构建.草业科学,2012,29(4):549-553. Ma Q,Wang S M.Construction of RNAi expression vector of the plasma membrane Na+/H+antiporter gene in succulent xerophyteZygophyllumxanthoxylum.Pratacultural Science,2012,29(4):549-553.(in Chinese)
[38] 徐春波,王勇,李兴酉,赵海霞.含有拟南芥转录因子CBF4基因植物表达载体的构建.草业科学,2010,27(6):88-92. Xu C B,Wang Y,Li X Y,Zhao H X.Construction of plant expression vector containingArabidopsistranscriptionfactorCBF4.Pratacultural Science,2010,27(6):88-92.(in Chinese)
[39] 段珍,狄红艳,张吉宇,霍雅馨,孔令芳.无芒隐子草CsLEA基因超表达载体和反义表达载体构建.草业科学,2014,31(8):1475-1480. Duan Z,Di H Y,Zhang J Y,Huo Y X,Kong L F.Construction of the over and antisense expression vector ofCsLEAgene.Pratacultural Science,2014,31(8):1475-1480.(in Chinese)
[40] 彭小群,张博雅,刘洁雯,王亚,解新明.象草PpCCR基因正、反义表达载体的构建及对烟草的转化.草业科学,2015,32(10):1586-1593. Peng X Q,Zhang B Y,Liu J W,Wang Y,Xie X M.Construction of sense and antisense expression vectors ofPpCCRgene inPennisetumpurpureumand transformation to tobacco plants.Pratacultural Science,2015,32(10):1586-1593.(in Chinese)
[41] 魏开发,刘逸萍,林子英,杨雅芳,张泽宏,贾文锁.农杆菌介导单子叶植物遗传转化问题与对策.植物学通报,2008,25(4):491-496. Wei K F,Liu Y P,Lin Z Y,Yang Y F,Zhang Z H,Jia W S.Problems and solutions inAgrobacteriumtumefaciens-mediated genetic transformation of monocotyledons.Chinese Bulletin of Botany,2008,25(4):491-496.(in Chinese)
[42] 贺晨霞,夏光敏.农杆菌介导单子叶植物基因转化研究进展.植物学通报,1999,16(5):567-573. He C X,Xia G M.Recent advances in gene transformation of monocotyledons mediated byAgrobacteriumtumefaciens.Chinese Bulletin of Botany,1999,16(5):567-573.(in Chinese)
(责任编辑 武艳培)
Construction of RNAi expression vector of heavy metal ATPases geneHMA2 and genetic transformation inIrislactea
Zhang Ya-nan1,2, Lin Yin-ding1, Meng Li2, Mao Pei-chun2, Tian Xiao-xia2, Guo Qiang2
(1.Shanxi Agricultural University, Taigu 030801, China; 2.Beijing Research and Development Center for Grasses and Environment, Beijing Academy of Agriculture and Forestry Science, Beijing 100097, China)
Heavy metal ATPasesHMA2 mediated Cd long-distance transport plays important roles in the distribution and detoxification of Cd2+in plants. However, the molecular mechanism underlying Cd2+long-distance transport inIrislactearemains unknown. Total RNA was extracted from the roots ofI.lacteato obtain a fragment ofIlHMA2 by RT-PCR. Then RNAi expression vector pARTG1G2 with inverted repeats ofHMA2 driven by the promoter CaMV35S was constructed based on the intermediate vector pHANNIBAL and the plant expression vector pART27 by restriction enzyme digestion and ligation. Moreover, the recombinant plasmid was transformed into agrobacterium GV3101 by freezing and thawing method, and then was transformed intoI.lacteato obtainIlHMA2-RNAi lines. This would be a good basis for clarifying theIlHMA2 mediated Cd2+long distance transport in the adaptation ofI.lacteato Cd2+contaminated environments.
Irislactea; heavy metal ATPases; Cadmium accumulation and tolerance; RNAi vector construction
Lin Yin-ding E-mail:sxnd_lyd@sohu.com Guo Qiang E-mail:guoqiang@grass-env.com
10.11829/j.issn.1001-0629.2016-0571
张亚楠,蔺银鼎,孟林,毛培春,田小霞,郭强.马蔺重金属ATP酶基因HMA2的RNAi载体构建及其遗传转化.草业科学,2017,34(5):988-996.
Zhang Y N,Lin Y D,Meng L,Mao P C,Tian X X,Guo Q.Construction of RNAi expression vector of heavy metal ATPases geneHMA2 and genetic transformation inIrislactea.Pratacultural Science,2017,34(5):988-996.
2016-11-15 接受日期:2017-01-17
北京市自然科学基金(6152008);北京市农林科学院科技创新能力建设专项(KJCX20140103、KJCX20170110)
张亚楠(1990-),女,山西晋城人,在读硕士生,主要从事园林植物与观赏园艺研究。E-mail:972388289@qq.com
蔺银鼎(1955-),男,山西临汾人,教授,博士,主要从事园林生态与景观规划设计研究。E-mail:sxnd_lyd@sohu.com共同通信作者:郭强(1981-),男,甘肃天水人,助理研究员,博士,主要从事草类植物逆境生理与基因工程研究。E-mail:guoqiang@grass-env.com
Q943.2
A
1001-0629(2017)05-0988-09
猜你喜欢
环球时报(2022-09-20)2022-09-20
食品安全导刊(2021年21期)2021-08-30
江西农业学报(2021年4期)2021-04-20
今日农业(2020年24期)2020-12-15
现代检验医学杂志(2016年2期)2016-11-14
小资CHIC!ELEGANCE(2015年15期)2015-09-01
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
现代检验医学杂志(2015年6期)2015-02-06
