
水翁幼苗生长规律研究
2017-06-05 15:18:31李碧洳翁殊斐冯嘉仪欧泳欣
浙江林业科技 2017年1期
李碧洳,翁殊斐,冯嘉仪,欧泳欣,董 淼
(华南农业大学 林学与风景园林学院,广东 广州 510642)
水翁幼苗生长规律研究
李碧洳,翁殊斐,冯嘉仪,欧泳欣,董 淼
(华南农业大学 林学与风景园林学院,广东 广州 510642)
对水翁Cleistocalyx operculatus幼苗的生长过程进行研究。结果表明,水翁幼苗的株高、地径生长具有阶段性的特点,利用Logistic曲线拟合了水翁幼苗株高与地径的生长模型,决定系数均在0.95以上;其生长进程可划分为4个阶段,即出苗期、生长前期、速生期和生长后期;根冠比总体呈下降趋势,生长重心逐渐由地下部分向地上部分转移;叶面积比与叶根比呈先下降后上升的趋势;地径与主根长、单株总叶面积等形态指标以及地上生物量、地下生物量以及单株总生物量等生物量指标之间均存在较强的正相关关系(r>0.92,P<0.01);根冠比与株高、地径、单株总叶面积、主根长、地上生物量、地下生物量、单株总生物量均存在负相关关系;地上生物量、地下生物量、单株总生物量之间均存在极显著的正相关关系(r>0.97,P<0.01)。
水翁幼苗;Logistic生长模型;形态指标;生物量指标;相关性分析
华南地区属亚热带季风气候,冬季干旱,夏季多雨,且由城市化建设引起的水涝时有发生,城市特殊的立地条件制约了园林树木的生长。水翁 Cleistocals op erculatus,又名水翁蒲桃,为桃金娘科 Myrtaceae 水翁属Cleistocals常绿乔木[1],主要分布于中南半岛、印度、马来西亚、印度尼西亚及大洋洲等地,枝叶繁茂,聚伞花序,花白色,具有一定的芳香,常作为绿荫树与园景树栽植。水翁是我国华南地区常见的、适应性较广的一种优良乡土树种,在干旱、潮湿、半淹、全淹条件下均能生存[2],且具有固土护坡、净化水体的能力[3-4],可作为海绵城市建设和滨水绿化的参考树种之一[5-6]。
苗木的生长节律受到树木遗传特性以及所在的环境条件等因素的影响而表现不同,在栽培养护、种源筛选等方面的研究具有重要的意义[7-8],对水翁的苗期生长节律进行研究,了解水翁的苗木生长变化规律,有助于培育高质量苗木。目前关于水翁的研究主要集中在耐涝性研究方面[2-4,9],关于其幼苗生长变化的研究很少。本文以水翁幼苗为研究对象,通过定期观察,测定其株高、地径生长、生物量等方面的变化,揭示生长发育规律特点,以期为水翁的栽培管理和华南地区乡土植物的生长发育研究提供借鉴和参考。
1 材料与方法
1.1 试验地概况
试验地设在广东省广州市华南农业大学校内的塑料大棚,属于南亚热带季风气候区,年平均气温 21.5 ~22.2℃,1月为全年最冷月,月平均气温为9 ~ 16℃,7月为全年最热月,月平均气温达28.7℃;平均年降水量1 800 mm,降水主要集中在4-9月的汛期,占全年雨量的80 %左右;年平均相对湿度78 %。
1.2 试验材料
挑选华南农业大学校园绿地内生长10 ~ 20年生生长健康、无病虫害的水翁,2014年7月底采集其成熟果实,淘洗去皮肉得纯净种子后于室内摊开晾干。2014年8月1日,选择颗粒饱满、无病虫害、外表完整无损伤、千粒重约为233 g的种子,用0.1 %的次氯酸钠溶液浸泡l min消毒,用清水冲洗干净后采用条播的方式,在宽0.8 ~ 1.0 m、基质为细河沙的沙床上播种,播种后细沙覆盖1 ~ 2 cm,期间保持沙床湿润。种子萌芽后3个月,将置于沙床内的幼苗定植于上口径为16 cm,深18 cm的无纺布袋中,栽培基质:壤土5份、泥炭土3份、河砂2份混合,每袋移植1株幼苗。幼苗上袋后用透光度60% ~ 70 %的遮光网遮荫,期间保持正常管理。
1.3 试验方法
水翁种子在播种后9 d开始发芽,15 d后(2014年8月15日)出苗基本整齐。选择株高、地径基本一致的幼苗100株,随机抽取其中5株作为固定样株,8月20日进行第1次测量,此后每月15日用直尺测定其株高、用游标卡尺测定地径;挑选5株标准木,用WinFolia软件测定叶面积,用直尺测量主根长,将植株分为地上部分、地下部分,置于烘箱内烘至恒重,分别测量地上生物量、地下生物量,并计算根冠比、单株总生物量、叶面积比以及叶根比[11],2015年7月15日最后1次测量时幼苗平均株高为33.5 cm,平均地径为4.73 mm。
1.4 数据处理
所有试验数据采用Excel进行整理;利用SPSS 21.0统计软件进行数据处理;对水翁幼苗的株高及地径生长量作Logistic生长曲线拟合[7],以此为依据对水翁幼苗生长进行生长阶段划分;用ANOVA方差分析法探究生长时间变化对各指标的影响;采用 Duncan’s多重比较法分析各指标不同生长时间的差异性;用 Pearson product-moment correlation coefficient 分析法探究各测量指标间的相关性,用OriginPro 8.5进行图像处理。
2 结果与分析
2.1 幼苗生长规律
2.1.1 株高生长规律 从图1可知,水翁幼苗的株高在种子萌芽后前30 d(9月15日)增长明显,净生长量达5.55 cm,占总生长量的22.27%,但增长幅度相对较小,持续时间较短;幼苗在9月至翌年2月高生长变缓,净生长量仅为3.9 cm,为总生长量的11.64%,尤其12月至翌年2月生长几乎停滞,估计是逐渐进入冬季,气温不断下降,幼苗处于休眠或相对缓慢生长期。3月开始进入春季,气温不断升高,苗木生长加快,生长高峰出现在5月,净生长量达9.54 cm,其后有所减缓;而6-7月生长速度明显减缓。
2.1.2 地径生长规律 从图2可知,种子萌芽后的前30 d(9月15日)水翁幼苗的地径生长较快速,净生长量占总生长量的26.12%;9月至翌年2月生长相当缓慢,净生长量为0.43 mm,占总生长量的9.19%。3月开始,地径生长逐渐加快,进入快速生长期,生长高峰出现在6月,比株高生长的高峰稍晚,表明水翁幼苗的茎、枝是先进行伸长生长再进行粗生长;6-7月地径生长有所减缓。
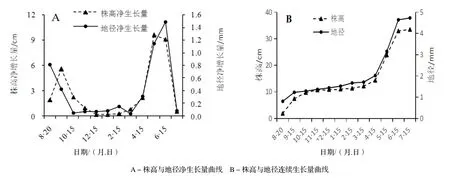
图1 水翁幼苗株高地径生长变化Figure 1 Growth of height and ground diameter of C. operculatus seedlings
2.2 生长模型建立与生长阶段划分
利用SPSS 21.0统计软件拟合水翁幼苗株高和地径的Logistic生长曲线模型(表1)。从表1可知,拟合情况较好(决定系数R2>0.95),可将水翁幼苗的株高生长阶段划分为4个阶段,出苗期8月1日至8月15日,生长前期8月15日至翌年3月4日,速生期3月4日至6月26日,生长后期6月26日之后;地径的生长阶段也可划分为4个阶段,出苗期8月1日至8月15日,生长前期为8月15日至翌年3月8日,速生期3月8日至7月10日,生长后期7月10日之后。株高和地径的速生期在春初至夏中,持续时间分别为109 d以及120 d。

表1 水翁幼苗株高和地径的Logistic生长曲线模型Table 1 Logistic regression for height and ground diameter growth of C. operculatus seedlings
2.3 根系和叶面积变化
由图2可知,生长时间对水翁幼苗的单株总叶面积、主根长的生长变化有极显著影响(P<0.01)。总叶面积随着生长时间总体呈上升趋势。单株总叶面积在8月至翌年3月(种子萌芽后5 ~ 180 d)生长缓慢,仅占全年生长量的7.41%,差异不明显,这可能是由于冬季低温以及较短的日照时间抑制了叶片生长;3月开始,随着气温的升高,单株总叶面积生长速度明显加快。水翁幼苗的主根长在萌芽后前30 d(9月15日)生长速度较快,净生长量占全年总生长量的38.37%,增长明显;9月至翌年3月生长速度减缓,差异较小,净增长量占总生长量的10.74%;3-7月主根长的生长速度明显增快,净增长量约占全年的生长量的50.89%,各月之间差异显著。总体而言,单株总叶面积与主根长的生长趋势基本一致,8月至翌年3月,根系的生长速率比单株总叶面积快,说明生长前期幼苗以地下根系生长为主,对土壤水分及养分的需求较大;3-7月,单株总叶面积的生长速率比根系快,说明速生期内幼苗的光合能力加强。生长时间对水翁幼苗的叶面积比、叶根比的变化有极显著影响(P<0.01)。叶面积比、叶根比都呈现随时间先下降后上升的趋势(图3),8月至翌年3月之间呈下降趋势,8-10月下降速度最快,10月至翌年3月下降速度减缓;3-6月期间先上升后下降,3-4月明显上升,4-6月有所下降;6-7月稍有上升,但差异不显著。
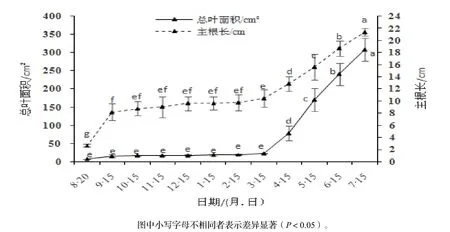
图2 水翁幼苗单株总叶面积和主根长生长变化Figure 2 Growth of total leaf area and main root length of C. operculatus seedlings
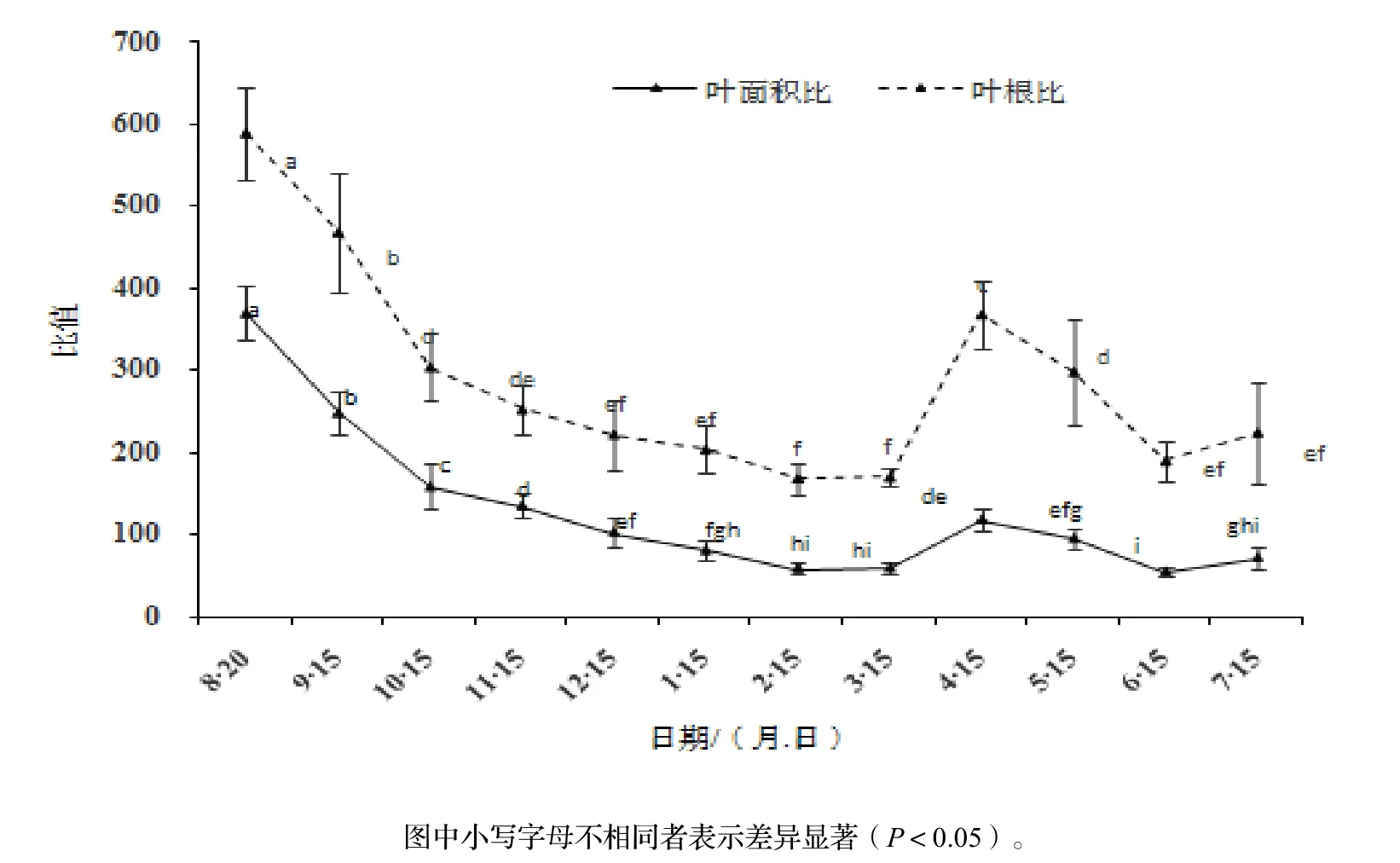
图3 水翁幼苗叶根比和叶面积比变化Figure 3 Leaf area ratio and leaf area root ratio of C. operculatus seedlings
2.4生物量分配变化
由图 4可知,根冠比随时间总体呈下降趋势,表明水翁幼苗的生长逐渐从地下部分转移到地上部分。8月20日根冠比达到最大,表明此时幼苗的生长重心在地下部分;8-9月根冠比明显下降,下降幅度33.41%,表明地上生物量积累速度明显超过地下部分;而9-11月的根冠比值没有明显差异,表明这段时间的地上、地下部分的生长速度差异不大。
11月至翌年 2月各月的根冠比明显下降,各月下降幅度分别为上月的 25.02%,23.83%,18.94%;2-6月根冠比依然呈下降趋势,但下降速度明显减少,相邻月份之间没有明显差异;6-7月根冠比略有上升,但幅度较小,差异不明显。8-11月根冠比均大于 1,表明这个时期幼苗把更多的营养物质分配到地下部分;12月中旬及之后根冠比值均小于 1,表明更多的光合产物被分配到幼苗的地上部分。
2.5 生物量与形态指标的相关分析
对各指标之间进行相关性分析(表2)。由表2可知,株高与其他形态指标间的相关性较弱(平均r<0.55);地径、主根长以及单株叶面积三个形态指标之间有较强的正相关关系(r>0.85),其中地径与单株叶面积的相关系数较高。地上生物量、地下生物量、单株总生物量与地径的相关系数最高(r>0.97);其次为单株总叶面积、主根长和株高。地上生物量与地下生物量之间存在密切的正相关关系(r=0.977),单株总生物量与地上生物量、地下生物量之间的关系极为密切(r>0.98),可以相互之间预测生长情况。根冠比与株高、地径、单株总叶面积形态指标之间存在极显著的负相关性,与叶面积比、叶根比之间存在正相关关系,其中与叶面积比的关系较密切;根冠比与地上生物量、地下生物量、单株总生物量均呈负相关,但相关性不强。
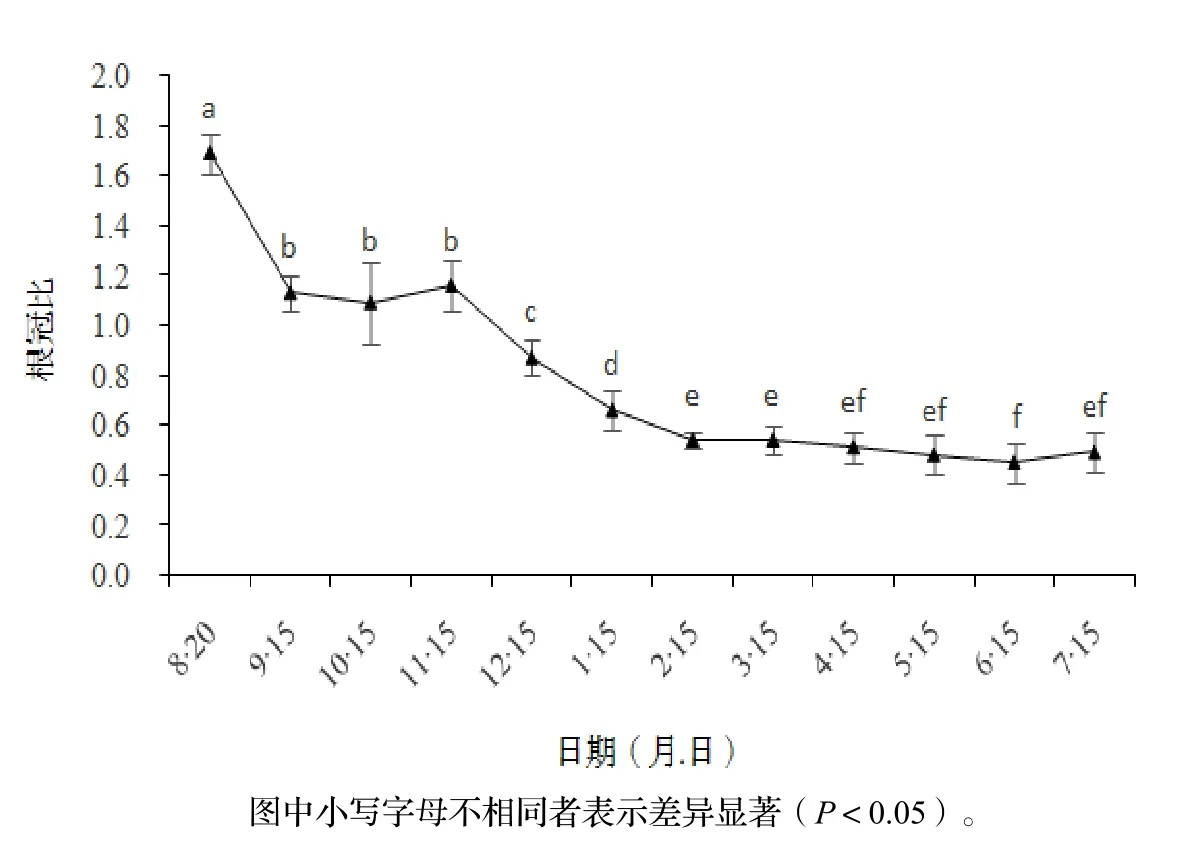
图4 水翁幼苗生物量分配变化Figure 4 Biomass allocation of C. operculatus seedlings
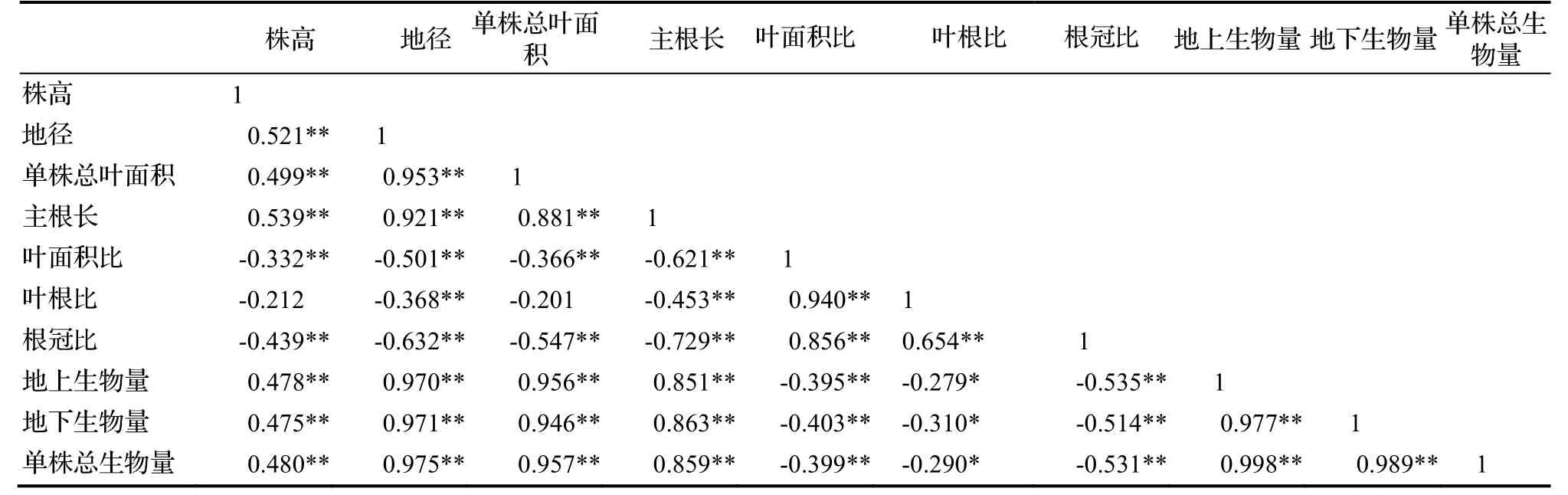
表2 生物量与形态指标的相关性分析Table 2 Correlation analysis on biomass with morphological indicators
3 结论与讨论
苗木的生长节律受到树木遗传特性以及所在的环境条件等因素的影响而表现不同[8]。研究发现水翁幼苗株高、地径的生长趋势基本一致,具有明显的阶段性。利用Logistic曲线拟合了水翁幼苗株高与地径生长量变化,决定系数均大于0.95。根据水翁幼苗的生长特点将株高与地径的生长阶段划分为:出苗期(8月1日至8月15日),生长前期(株高8月15日至翌年3月4日,地径8月15日至翌年3月8日),速生期(株高3月4日至6月26日,地径3月8日至7月10日)、生长后期(株高6月26日之后,地径7月10日之后)。研究结果表明,水翁属于春夏生长型的苗木,主根长以及单株叶面积在速生期内生长速度也明显加快,苗木的水肥消耗量也逐渐增大,因此期间要加强对苗木的水肥供给,并提供合适的生长环境,以保证苗木质量。地径与株高的年生长规律基本相似,但地径的生长期比株高滞后,说明水翁幼苗是先进行高生长再进行粗生长,这与南酸枣Choerospondias axillaris,山杜英Elaeocarpus sylvestris等阔叶树种的生长规律有一定的相似性[10]。南方红豆杉Taxus chinensis var. mairei,降香黄檀Dalbergia odorifera等树种在速生期内出现第一次生长高峰后进入暂缓期,随后进入第二次或更多次生长高峰[8,11]。研究结果显示夏播的水翁幼苗翌年的3-7月的速生期内出现一次生长高峰后下降,而水翁幼苗在第二年的生长期内是否存在二次或多次生长高峰还需进一步的试验。
生物量分配策略反映了光合产物在地上与地下各器官之间的不同投资分配[12-13],根冠比反映了光合产物在地上部分与地下部分之间的分配做出调整[14-15]。水翁幼苗的根冠比总体呈下降趋势,这与前人的研究结果相似[12]。种子萌芽初期(8月20日)根冠比最大,说明种子内贮藏的营养更多地分配到根系,促进根系生长以获得土壤的养分和水分来满足植株较高的生长发育需求,以支持植株形态建成;此后,根冠比逐渐降低,幼苗生长中心逐渐由地下部分向地上部分转移,更多的光合产物被分配到幼苗的地上部分;速生期内(3-7月)根冠比下降速度明显减缓,叶面积比和叶根比有所上升,可能是由于速生期内叶面积、茎干等地上部分的生长速度明显加快,地上部分形成了更为致密的冠层空间,光合面积增大,更能充分捕获和利用光资源来促进地上生物量的形成。
地径能较好地反映苗木质量[16]。本研究的相关分析显示,地径与主根长、单株总叶面积等形态指标极显著正相关,可以相互预测其生长情况;地径与地上生物量、地下生物量、单株总生物量等指标之间存在最显著的正相关关系(r>0.97,P<0.01),比主根长、单株总叶面积等形态指标更能反映幼苗的生物量累积情况,这与周鹏等的结果基本一致[17];地上生物量与地下生物量之间具有极显著的正相关(r=0.977,P<0.01),表现出较好的生长一致性,这表明可通过地上部分的生长预测地下根系的生长情况,也可通过改善光照、温度等的条件促进地上生物量的积累,从而促进地下根系的生长[18]。构建植物指标间的回归模型是研究植物生长发育规律的一种常用方法,而指标间的相关性分析是构建回归模型的基础,下一步的研究可尝试结合多个指标建立形态指标、生物量指标、或形态指标与生物量指标之间的回归模型,用于其生长规律的研究。
[1] 中国科学院中国植物志编辑委员会. 中国植物志:第53(1)卷[M]. 北京:科学出版社,1984:118.
[2] 靖元孝,程惠青,彭建宗,等. 水翁(Cleistocalyx operculatus)幼苗对淹水的反应初报[J] .生态学报,2001,21(5):810-813.
[3] 靖元孝,杨丹菁,任延丽,等. 水翁(Cleistocalyx operculatus)在人工湿地的生长特性及对污染物的去除效果[J]. 环境科学研究,2005,18(1):9-12.
[4] 郑中华,许大彬,孙谷畴. 湖榕、水翁混交护岸林带绿化固土效果研究[J]. 中国水土保持,2000,(11):15-17.
[5] 中华人民共和国住房和城乡建设部组织编制. 海绵城市建设技术指南――低影响开发雨水系统构建(试行)[M]. 中国建筑工业出版社, 2015.
[6] 付奇峰,方华,林建平. 华南地区水库消涨带生态重建的植物筛选[J]. 生态环境,2008,17(6):2325-2329.
[7] 邝雷,邓小梅,陈思,等. 4个任豆种源苗期生长节律的研究[J]. 华南农业大学学报,2014,35(5):98-101.
[8] 李苏珍,温莉娜. 南方红豆杉一年生苗木生长规律及相关关系研究[J]. 浙江林业科技,2014,34(04):76-78.
[9] 应梦云,冯志坚,曹中元. 水翁幼苗耐涝试验[J]. 福建林业科技,2013,40(4):79-81.
[10] 刘鲁平,李闪金,刘光正,等. 25种优良阔叶树种苗木生长节律研究[J]. 江西林业科技,2001,(3):10-13.
[11] 吴国欣,王凌晖,俞建妹,等. 降香黄檀幼苗年生长节律研究[J]. 浙江林业科技,2010,30(03):56-60.
[12] 杨昊天,李新荣,刘立超,等. 荒漠草地4种灌木生物量分配特征[J]. 中国沙漠,2013,33(5):1340-1348.
[13] 李刚,赵祥,张宾宾,等. 不同株高的柠条生物量分配格局及其估测模型构建[J]. 草地学报,2014,22(4):769-775.
[14] 李旭东,张春平,傅华. 黄土高原典型草原草地根冠比的季节动态及其影响因素[J]. 草业学报,2012,21(4):307-312.
[15 马晓东,朱成刚,李卫红. 多枝柽柳幼苗根系形态及生物量对不同灌溉处理的响应[J]. 植物生态学报,2012,36(10):1024-1032.
[16] Bayala J, Dianda M, Wilson J, et al. Predicting field performance of five irrigated tree species using seedling quality assessment in Burkina Faso, West Africa[J]. New For,2009,38(3):309-322.
[17] 周鹏,翁殊斐,柯羽,等. 6种园林花灌木幼苗生长及生物量的分配[J]. 西北林学院学报,2015,30(6):134-138.
[18] 周鹏,翁殊斐,杭夏子. 5种龙船花属植物幼苗生长与生物量的研究[J]. 西北林学院学报,2014,29(3):75-78.
Growth Rhythm of Cleistocalyx operculatus Seedlings
LI Bi-ru,WENG Shu-fei,FENG Jia-yi,Ou Yong-xin,DONG Miao
(College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642, China)
Experiments were conducted on growth rhythm of Cleistocalyx operculatus seedlings in Guangzhou, Guangdong province in 2014. The results show that the growth of height and ground diameter of C. operculatus had different stages. Logistic growth curve was fitting the growth changes, with determination coefficient above 0.95. Height and diameter growth of seedlings could be divided into 4 stages with nearly similar date. Root-shoot ratio decreased indicating underground growth increased while aboveground growth decreased. Leaf area ratio and leaf area root ratio decreased and then increased. There was a significant positive correlation among ground diameter and main root length, total plant leaf area, aboveground biomass, underground biomass , the total plant biomass(r >0.92,P<0.01). Significant negative correlation existed among root-shoot ratio and height, ground diameter, total plant leaf area, main root length, aboveground biomass, underground biomass, the total plant biomass. Significant positive correlation between aboveground biomass, underground biomass, and the total plant biomass was observed(r>0.97,P<0.01).
Cleistocalyx operculatus seedling; logistic growth model; morphological index; biaomass index; correlation analysis
S723.1+31
A
1001-3776(2017)01-0041-06
10.3969/j.issn.1001-3776.2017.01.007
2016-08-30;
2016-11-01
广东省科技计划项目(2013B020305008),广东省林业厅项目(14KJCX005-02)
李碧洳,硕士研究生,从事园林植物造景与应用研究;E-mail:549239038@qq.com。通信作者:翁殊斐,博士,副教授,从事园林植物研究;E-mail:shufeiweng@scau.edu.cn。
猜你喜欢
绿色科技(2021年21期)2021-11-26 09:12:06
内蒙古林业调查设计(2021年3期)2021-07-06 14:12:00
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
新疆农业科学(2020年1期)2020-02-14 03:22:46
中国科技纵横(2018年14期)2018-08-30 09:15:34
浙江农林大学学报(2018年4期)2018-08-22 01:54:12
现代园艺(2018年1期)2018-03-15 07:56:13
现代农业科技(2017年16期)2017-09-22 14:47:46
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中国麻业科学(2015年5期)2015-12-28 06:22:11
