
鸡源锌指蛋白ZEB1基因解析及其在新城疫病毒感染中的作用研究
2017-06-05 15:19张耀丹任亭亭孟春春孙英杰宋翠萍仇旭升罗廷荣
中国动物传染病学报 2017年2期
汪 伟,张耀丹,任亭亭,孟春春,谭 磊,孙英杰,宋翠萍,廖 瑛,仇旭升,丁 铲,罗廷荣
(1.广西大学动物科技学院,南宁 530003;2.中国农业科学院上海兽医研究所,上海 200241)
·研究论文·
鸡源锌指蛋白ZEB1基因解析及其在新城疫病毒感染中的作用研究
汪 伟1,2,张耀丹2,任亭亭2,孟春春2,谭 磊2,孙英杰2,宋翠萍2,廖 瑛2,仇旭升2,丁 铲2,罗廷荣1
(1.广西大学动物科技学院,南宁 530003;2.中国农业科学院上海兽医研究所,上海 200241)
锌指蛋白ZEB1(zinc finger E-box binding homeobox 1)是一种在不同物种中相对保守的转录因子,尤其在发生肿瘤的组织中具有较高的表达水平。为了证实禽类锌指蛋白ZEB1在新城疫病毒(Newcastle disease virus,NDV)感染中的作用,本研究扩增获得了鸡源ZEB1的基因序列,并通过生物信息学软件对不同物种的ZEB1基因进行了同源性比对。通过Real-time PCR技术测定NDV感染DF-1细胞、CEF细胞以及SPF鸡后ZEB1的表达量变化,证实NDV感染可使细胞中的ZEB1含量显著升高。但是在细胞中过表达ZEB1对NDV的增殖和蛋白表达没有明显的影响,其生物学意义有待进一步研究。
新城疫病毒;锌指蛋白;病毒增殖
锌指蛋白ZEB1(zinc finger E-box binding homeobox 1)又称为TCF8或δ EF1,是一种在不同物种中相对保守的转录因子,在人类器官中广泛表达,尤其是在发生肿瘤的组织中具有较高的表达水平。ZEB1在癌细胞上皮细胞向间充质细胞转化(epithelial-to-mesenchymal transition,EMT)的调控中起到了重要作用,在肿瘤转移过程中扮演着关键作用[1,2]。众所周知,细胞的迁移运动不仅参与肿瘤的发生发展,同时也在炎症反应、病毒感染以及多种细胞重要生理活动中发挥关键作用。已有研究指出,锌指蛋白ZEB1可能以不同的方式参与了病毒的感染过程。ZEB1可以通过抑制柯萨奇病毒和腺病毒相关受体的表达从而降低病毒的感染性。在EB病毒(Epstein barr virus)的研究中,ZEB1也表现出了对病毒的抑制作用[3-5]。尽管如此,ZEB1对于其他病毒感染的影响尚未有相关报道。
新城疫病毒(Newcastle disease virus,NDV)强毒引起的新城疫是一种重要的禽类烈性传染病,可导致多种禽类急性死亡,严重威胁世界养禽业的发展,被国际兽疫局(OIE)列为必须上报的动物传染病之一。近年来,越来越多的研究集中在了NDV的溶瘤机制。早在1955年,NDV就被发现可以在肿瘤细胞中增殖并杀死肿瘤细胞,但不感染正常细胞。NDV因快速增强免疫细胞对肿瘤细胞的杀伤力而对人无致病性的特点,已经作为一种溶瘤生物制剂被广泛研究,一些毒株已进入临床试验阶段[6]。但NDV特异性杀灭肿瘤细胞的机制尚不明确。在前期研究中,我们通过高通量测序技术发现NDV感染细胞会引起肿瘤相关因子ZEB1和ZEB2基因的大幅上调。为了验证ZEB1在NDV感染中表达量的变化,本研究中首次扩增了鸡源的ZEB1基因,建立了Realtime PCR检测方法,并对ZEB1的体内外表达情况进行了检测。
1 材料与方法
1.1 菌株、载体、病毒、细胞和试剂 BL21和DH5α购自天根生化科技有限公司;Herts /33、La Sota株购自中国兽医药品监察所,由本实验室保存;DF-1细胞用含5% 胎牛血清(fatal bovine serun,FBS)的
DMEM 培养基(购自Gibco),维持液为含有2%胎牛血清的DMEM 培养基;SY BR Green和高保真酶Phanta Max Super-Fidelity DNA Polymerase购自南京诺唯赞生物科技有限公司;RNasin和MLV Reverse Transcriptase购自Promega公司;琼脂粉、琼脂糖、抗生素购自上海生工生物工程有限公司;Poly (I:C)(HMW)购自Invivogen公司。
1.2 方法
1.2.1 ZEB1基因扩增引物的设计 以GenBank中收录的鸡的ZEB1基因(登录号:KY 362200)做参考序列设计引物,送生工生物工程(上海)股份有限公司进行合成。ZEB1-F:5'-GCGAGCTCGAGATGG CGGATGGCCCCAGGTGTAA-3';ZEB1-R:5'-CG CGGATCCCGTTAAGCTTTATTTGTTTTTTCTTCT GAC-3',目的片段大小为3345 bp。
1.2.2 细胞总R NA提取及cDNA样品的制备 按常规方法制备鸡胚成纤维细胞(chicken embryo fibroblast,CEF)原代细胞。用Poly I:C刺激CEF细胞,24 h后收样,使用Trizol试剂(Invitrogen)提取CEF细胞中总RNA,用Mo-MLV(Promega)反转录制备cDNA模板后置于-20℃备用,具体操作步骤参照说明书进行。
1.2.3 PCR反应及基因克隆 使用设计好的引物,以CEF制备的cDNA样品为模板进行PCR,反应条件:95℃预变性3 min;95℃变性20 s,58℃退火30 s;72℃延伸4 min,30个循环;最后72℃延伸10 min。PCR产物经琼脂糖凝胶电泳回收后用Xho I和BamH I酶切,连接至pEGFP-C3载体。4℃连接过夜后,连接产物转化感受态细胞DH 5α。挑取细菌菌落,提取质粒,并使用Xho I和BamH I酶切鉴定。阳性质粒送生工生物工程有限公司测序。
1.2.4 不同物种ZEB1基因的差异性比对及进化分析
使用软件DNAStar软件的MegAlign程序,将获得的鸡的ZEB1基因序列与其他物种进行比较,并构建系统发育进化树。同时将获得的鸡的ZEB1基因序列上传至GenBank数据库,并对其结构域进行在线生物信息学分析。
1.2.5 ZEB1基因绝对荧光定量PCR标准曲线的建立
根据扩增获得的鸡ZEB-1基因序列,设计荧光定量引物,送生工生物工程(上海)股份有限公司进行合成。鸡ZEB1的qPCR引物对为:5'-ATGGAAC CTCTTGACCT-3'和5'-CACTTGGTGGGATTACA T-3';β-actin的qPCR引物对为:5'-GATCTGG CACCACACCTTCT-3'和5'-GGGGTGTTGAAGG TCTCAAA-3'。用S.W.稀释标准共稀释8个梯度,选取6个稀释度,分别为1×103、1×104、1×105、1×106、1×107、1×108copies/μ L。SYBR GreenⅠ采用Real-time PCR进行测定。20 μ L反应体系:SYBR Green qPCR Master Mix 10 μ L、上下游引物各1 μ L、模板1 μ L、S.W. 8 μ L。PCR反应程序:94 ℃预变性1 min;95℃变性15 s;60℃退火15 s;72 ℃延伸45 s,40个循环;最后插入仪器自带的溶解曲线步骤。根据荧光值的变化规律,系统将自动生成标准曲线和溶解曲线。
1.2.6 健康成年鸡组织中ZEB1表达量检测 取6周龄SPF级别鸡3只,静脉注射空气处死后分别取其脑、胸腺、肝脏等器官或组织,标记后分装保存。用组织匀浆机研磨后使用Trizol法提取总RNA,反转录制备cDNA模板。按标准曲线的换算关系计算出每克不同器官中ZEB1基因的含量。
1.2.7 SPF鸡感染NDV后组织脏器ZEB1表达量检测
取12只1日龄SPF级别小鸡,分成实验组和对照组,分开饲养。实验组小鸡进行肌肉注射NDV。分别在注射病毒1 d和3 d后,各组分别取3只小鸡静脉注射空气致死,取其肺脏、脾脏、肝脏等器官或组织,用组织匀浆机研磨后使用Trizol法提取总RNA,反转录制备cDNA模板。以β-actin基因为内参,每个样品设3个重复。结果处理采用常规的2-Δ Δ CT法,首先对所有的测试样本和校准样本,用内参基因的CT值归一目标基因的CT值,然后用校准样品的Δ CT值归一试验样本的Δ CT值,最后计算表达水平比率。
1.2.8 NDV感染CEF后ZEB1在不同时间点的表达水平
学生对该课程的认知度不高 针对机械电子工程本科生开设的机器人学课程,教学大纲要求学生熟悉机器人技术发展概况及其在生物生产、仿生机械、特种机器人等方面应用的具体案例,掌握基本构成、工作原理、动力学分析、传感控制系统等基本知识[4]。但在教学过程中发现,学生对该门课程的积极性不高,认为内容多、难度大,不易理解,特别是对运动学中矩阵的变换,运动学、动力学等理论的推导分析,产生消极态度,久而久之,导致失去学习的兴趣。
将CEF或DF-1细胞均匀铺于6孔细胞培养皿上,待细胞生长到60%~70%密度后使用。实验组细胞感染1MOI的Herts/33,对照组不感染,30 min后用PBS洗涤3次,维持液继续培养。在感染后4、8、12、24、36 h分别收样,提取总RNA后反转录制备cDNA,Real-time PCR检测ZEB1含量。1.2.9 ZEB1在NDV感染中的作用 将构建好的质粒pEGFP-ZEB1用转染试剂FUGEN转染DF-1细胞,48 h后以0.01MOI病毒剂量的Herts/33毒株感染细胞,以不感染病毒的细胞为对照组,不同时间收取上清和细胞样品。其中上清用Trizol提总RNA,反转录制备cDNA模板,Real-time PCR检测病毒含量;细胞样品用loading buffer 处理后使用Western blot方法检测NP蛋白。
2 结果
2.1 扩增ZEB1基因 在PolyI:C和刀豆素刺激CEF细胞的cDNA样品中检测出鸡的ZEB1基因,条带大小为3500 bp,与设计的检测性引物的扩增片段大小一致。随后,将ZEB1基因连接到pEGFP-C3载体上,酶切后电泳鉴定结果正确(图1)。测序结果显示所获取的ZEB1基因与网上公布序列一致,仅有3个核苷酸位点差异。其中,第905位和第3042位核苷酸为同义突变;第908位核苷酸由突变赖氨酸(Lys)突变为天冬氨酸(Asn)。
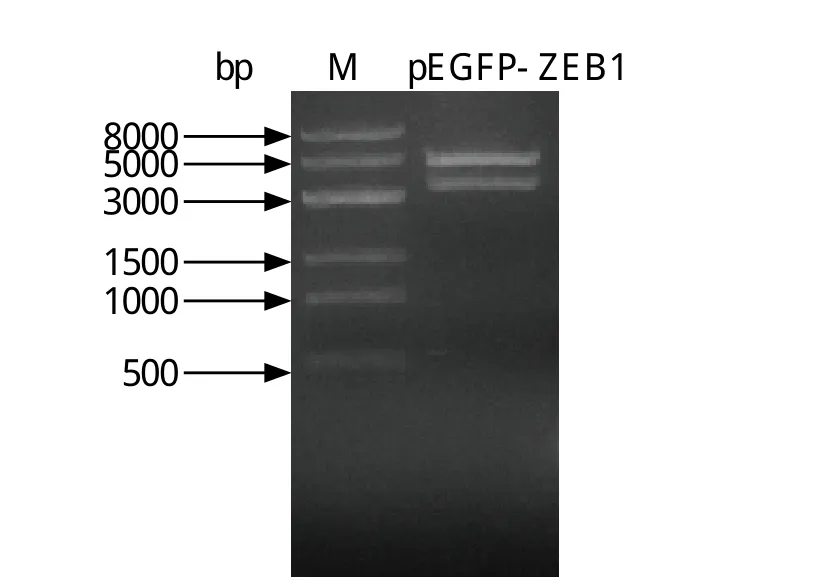
图1 鸡ZEB1基因的PCR扩增Fig. 1 PCR-amplified ZEB1 gene from CEF cDNA
2.2 ZEB1基因的序列对比分析 通过测序确定ZEB1基因的完整的阅读框序列。使用软件DNAStar软件将获得的ZEB1基因与其他物种进行比较,构建了系统发育进化树。结果显示,不同物种间ZEB1基因和蛋白质同源性很高。鸟类ZEB1之间核苷酸同源性较高,鸡源ZEB1与人、家鼠、非洲蟾蜍及海龟的核苷酸同源性也分别高达81.9%、78.2%、75.2% 和89.8%。禽类ZEB1基因与爬行动物如海龟的亲源性较近,其次是两栖类动物,最低的是哺乳动物(图2)。

图2 不同物种ZEB1基因的系统进化分析Fig.2 Phylogenetic analysis of ZEB1 gene from different species
2.3 ZEB1蛋白结构域的生物信息学分析 通过EBI的InterProScan数据库对ZEB1蛋白序列进行在线分析,结果显示ZEB1蛋白在C端有3个连续的锌指结构域,在N端则有4个不连续的锌指结构域。其中N端的最后1个和C端的3个锌指结构域还具有识别并结合特定的DNA序列,发挥调节靶基因转录的功能(图3)。在ZEB1蛋白的中部有一个同源盒基因序列(homeobox)。分析发现,不同物种间zinc finger结构域与homeobox结构域氨基酸序列同源性非常高,说明在物种进化过程中,ZEB1的zinc finger结构域与
homeobox结构域进化高度保守(图4)。通过分析发现人与鸡的ZEB1蛋白不但具有相同的结构域,其结构域内的氨基酸序列同源性也非常高,由此我们推断鸡源ZEB1蛋白与人源ZEB1蛋白的结构可能相同,而蛋白结构决定其功能,所以鸡源ZEB1蛋白功能应该与人十分接近。
2.4 ZEB1基因的真核表达 将构建好的pEGFP-ZEB1质粒转染到DF-1细胞中,24 h后收样,分别用免疫荧光法和Western blot方法鉴定ZEB1蛋白的表达。用荧光显微镜观察到了绿色荧光(结果未显示)。用GFP抗体进行Western blot,结果显示条带正确,说明该质粒在DF-1细胞中过表达成功(图5)。
2.5 绝对定量标准曲线 通过Real-time PCR仪扩增,自动检测并输出标准曲线、溶解曲线及溶解峰。溶解曲线表现为单一的峰值,说明引物的特异性很好,标准曲线的函数表示为:Y=-4.4346 X+50.419(R2=0.9984),拷贝数和Ct值在标准质粒稀释质量浓度范围内具有良好的线性关系。结果表明本研究建立的ZEB1基因标准曲线能够用于检测ZEB1基因(图6)。
2.6 NDV感染DF-1细胞后ZEB1基因表达的时相变化 通过Real-time PCR相对定量NDV感染DF-1细胞后,细胞中的ZEB1含量。结果显示,NDV感染早期,ZEB1出现了小量下调;感染后8 h,ZEB1表现出明显的上调;感染后24 h上调了160多倍(图7)。
2.7 NDV感染CEF对ZEB1表达的影响 为了进一步确认ZEB1在细胞中的表达情况,制作了鸡胚原代细胞,使用Herts/33感染后,收样检测ZEB1的表达情况,发现NDV感染原代CEF细胞后同样能够引起ZEB1大幅上调(图8)。
2.8 ZEB1在鸡组织器官中的表达水平 Real-time PCR检测发现,健康鸡体内心脏、肝脏、肾脏、肺脏、脑、胸腺、法氏囊和气囊等均含有一定量的ZEB1,并且在脑中含量最高;肠中未检测到ZEB1基因的表达(图9)。
2.9 NDV感染1日龄雏鸡后ZEB1表达的时相变化 随着时间推移,脾脏中的ZEB1基因表达量上调了3000多倍,而肝脏、肺脏和法氏囊上调不明显,十二指
肠中未检测到ZEB1基因的表达(图10)。
2.10 ZEB1在NDV感染中的作用 前期研究结果表明,DF-1或CEF细胞受到病毒刺激后,细胞中ZEB1含量均有明显上调,推测可能与病毒的感染与增殖有关。本研究将ZEB1在DF-1细胞上过表达24 h后感染NDV,结果发现过表达ZEB1不具有抑制或促进病毒的增殖作用(图11)。
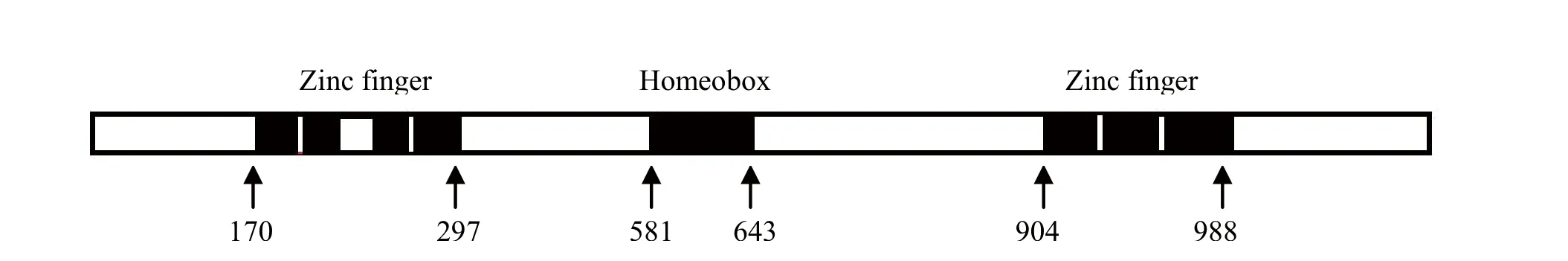
图3 鸡源ZEB1蛋白的结构域分析Fig.3 Domain analysis of chicken ZEB1 protein
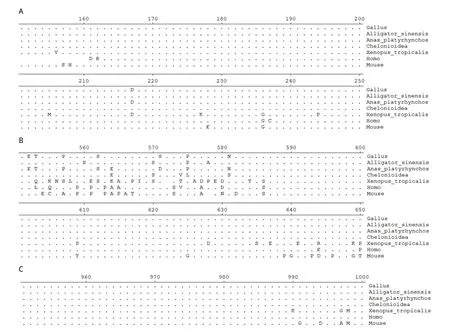
图4 鸡与其他物种ZEB1蛋白功能区序列对比Fig.4 Alignment of amino acid sequences of encoding region of ZEB1 protein between chicken and other speciesA: N端锌指结构域; B: Homeobox结构域2; C: C端锌指结构域A: N-zinc finger domain; B: Homeobox domain2; C: C-zinc finger domain
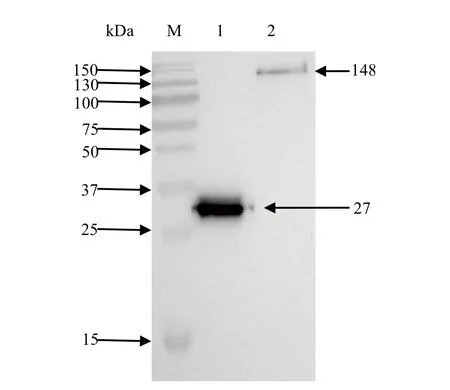
图5 pEGFP-ZEB1在DF1细胞中真核表达的检测Fig.5 Detection of expression of pEGFP-ZEB1 in DF1 cells
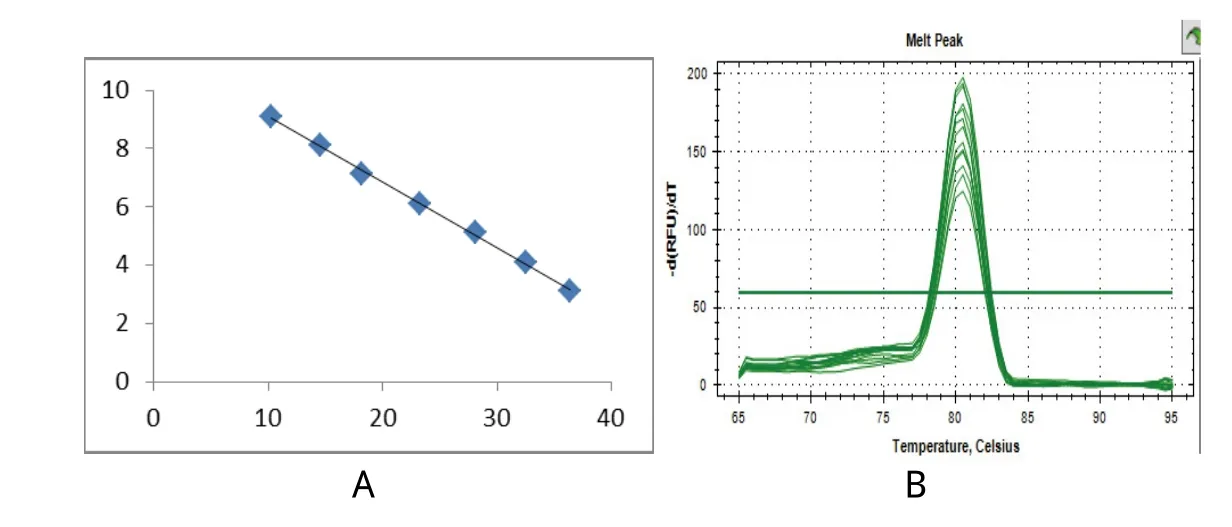
图6 Real-time PCR引物的优化和标准曲线的建立Fig.6 Optimization of the primers and construction of the standard curve for Real-time PCRA: 标准曲线; B: 熔解峰A: Standard curve; B: Melt peak
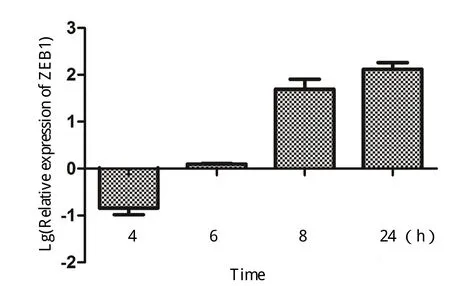
图7 NDV感染DF1细胞中ZEB1表达的Real-time PCR检测Fig.7 Expression of ZEB1 mRNA in NDV-infected DF-1 cells by Real-time PCR
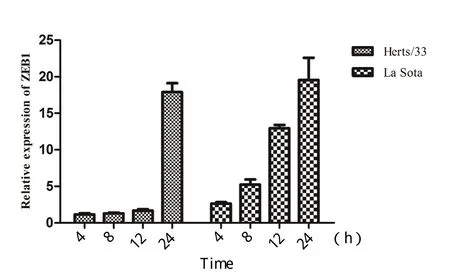
图8 NDV感染CEF细胞中ZEB1表达的Real-time PCR检测Fig.8 Expression of ZEB1 mRNA in NDV-infected CEF cells by Real-time PCR
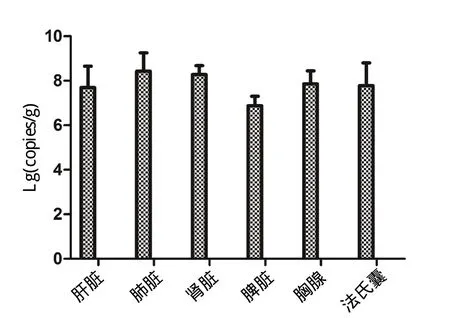
图9 每克鸡脏器组织中ZEB1的拷贝数Fig.9 Expression of ZEB1 gene in different tissues and organs of chicken
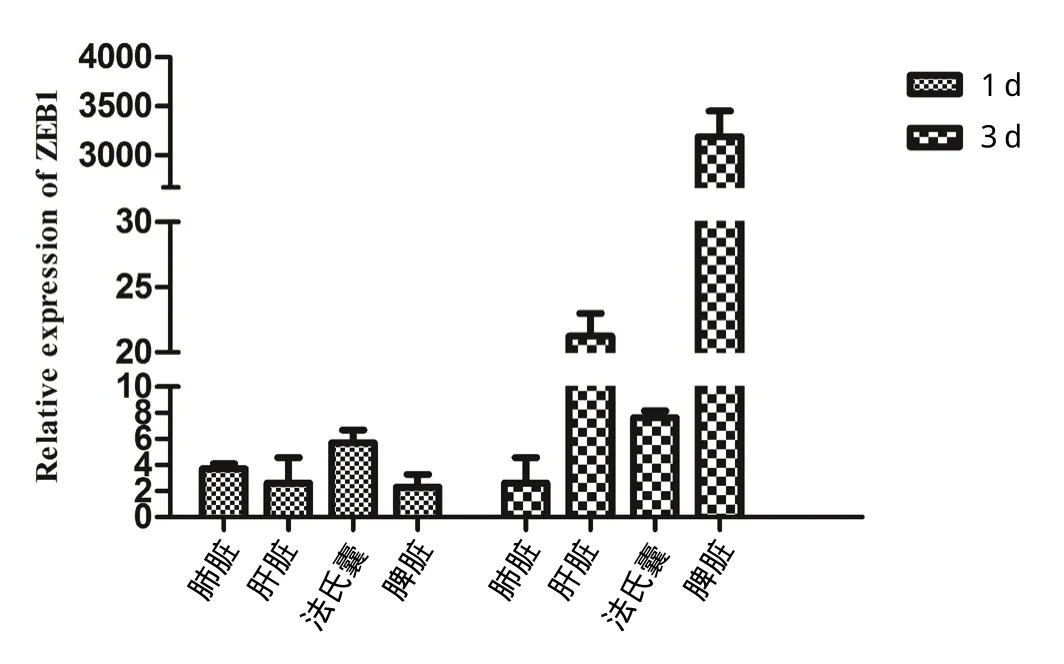
图10 NDV感染鸡组织脏器中ZEB1表达的Real-time PCR检测Fig.10 Expression of ZEB1 mRNA in different tissues and organs of NDV-infected SPF chickens by Real-time PCR
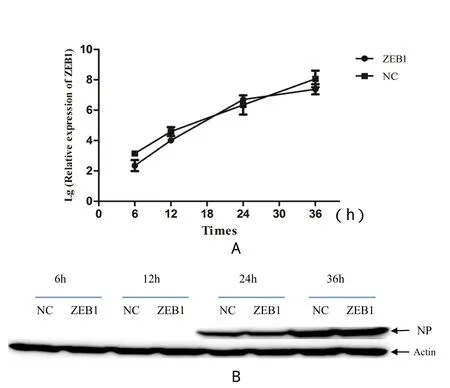
图11 过表达ZEB1对NDV的增殖影响Fig. 11 NDV replication level under ZEB1 overexpressionA: 细胞上清中的病毒量; B: Western blot检测DF-1细胞中NP蛋白; NC: 阴性对照组(转染pEGFP-C3); ZEB1: 实验组(转染pEGFPZEB1)A: The amount of virus in cell supernatant; B: NP protein detected in DF-1 cell by Western blot; NC: Negative control; ZEB1: Experimental group
3 讨论
本实验室前期研究中,通过高通量测序发现NDV感染细胞中ZEB1和ZEB2蛋白出现显著上调。ZEB1是一种锌指类转录因子,在人类组织器官中广泛表达,参与肿瘤干细胞干性维持、肿瘤侵袭转移及肿瘤血管生成等过程,可作为肿瘤的一项检测指标[7,8]。尽管如此,由于禽类ZEB1尚无研究报告,其上调的生物学意义以及分子机制不清楚。
本研究从CEF细胞的cDNA样品中扩增获得了鸡源ZEB1的基因序列,并建立了Real-time CPR检测方法。通过检测鸡源成纤维细胞系DF-1和原代成纤维细胞CEF感染NDV后ZEB1的表达情况,发现NDV缺失能够引起ZEB1的上调,且DF-1细胞中上调水平明显高于CEF细胞。NDV感染CEF后,ZEB1的表达量在感染后随时间推移不断上调,在24 h达到最高,上调了近20倍。NDV在感染DF-1后ZEB1表达量峰值出现在感染后24 h,上调了160倍。为了验证这一现象,我们使用SPF鸡进行了体内实验。正常动物组织中除了肠道外,在脾脏、肝脏、肺脏等器官中ZEB1均有表达,表达量差异不大,说明ZEB1在正常组织中广泛表达。SPF级雏鸡感染NDV后1 d,组织脏器中ZEB1小幅度上调,仅为3~5倍;而在感染3 d后各器官中ZEB1的含量就有明显地差异,尤其在免疫器官脾脏中表现最为明显,上调3400倍,提示ZEB1可能参与到NDV感染后的免疫应答过程。
人源ZEB1的研究显示,ZEB1可以降低柯萨奇病毒的感染性,抑制EB病毒的增殖[3,4]。为了确定ZEB1是否也能够抑制NDV的感染和增殖过程,本研究构建了ZEB1真核表达质粒并在DF-1细胞上进行过表达。结果显示,ZEB1在NDV感染后并没有发挥协同或拮抗作用。这与其他研究者在柯萨奇病毒、腺病毒以及EB病毒上的检测结果不一致,因此可以推测ZEB1并不直接参与NDV的感染过程。
通过生物信息学分析可以初步解析蛋白功能。我们研究发现,ZEB1基因在不同物种间高度保守,鸡源与人源ZEB1的核苷酸同源性高达81.9%。ZEB1蛋白的结构域也同样保守。通过EBI的InterProScan数据库对ZEB1蛋白序列进行在线分析,结果显示和人源ZEB1蛋白一样,鸡源ZEB1蛋白存在3个主要的功能结构域,即氨基端的锌指结构域、羧基端的锌指结构域以及蛋白中间的同源盒基因序列(homeobox)[9]。锌指结构域是ZEB1蛋白最主要的功能区域,具有识别并结合特定的DNA序列发挥调节靶基因转录的功能。氨基酸序列比对数据显示,鸡源ZEB1蛋白锌指结构域和homeobox基序与人源ZEB1蛋白的高度同源,表明鸡ZEB1蛋白的功能可能与人源ZEB1相似,即具有抑制内皮细胞(endothelial cell,EC)分化,进而抑制正常血管生成的作用;另一方面ZEB1可以降低信号素3F的表达从而提高细胞内HIF-1α的水平,以及HIF-1α靶分子V EGF水平,促进肿瘤血管的生成及肿瘤的扩散[2]。ZEB1是一种EC分化的负调节因子。研究表明,ZEB1能够直接与EC分化基因上游调控基因结合,从而抑制相关基因的表达。敲除内源性ZEB1基因后,EC分化相关基因显著上升[10]。EC分化是血管形成的重要环节,这一生理过程在肿瘤形成、胚胎发育过程中发挥关键作用[11]。NDV具有快速增强肿瘤细胞杀伤力而对人无致病性的特点,已作为一种溶瘤生物制剂被广泛研究并进入临床试验阶段。NDV感染后引起的ZEB1大幅上调可能在其溶瘤过程中有着重要的作用,其生物学意义需要进一步的研究。
本研究成功克隆了鸡ZEB1基因,通过Real-time PCR技术对NDV感染DF-1细胞、CEF细胞以及SPF鸡后ZEB1的表达量变化进行了检测,确定NDV感染可使细胞中的ZEB1含量升高提高。然而,过表达ZEB1对NDV没有明显的抑制作用,ZEB1在NDV感染中的作用有待进一步研究。
[1] 陈广, 韩梅芳. 人ZEB1基因真核表达载体的构建及其对肝癌细胞株HepG2增殖凋亡的影响[J]. 中西医结合肝病杂志, 2014, 24(1): 33-36.
[2] 苏文敬, 徐苗, 陈雪芹, 等. ZEB1在肿瘤中的研究进展[EB/OL]. 北京: 中国科技论文在线 [2013-12-13]. http:// www.paper.edu.cn/releasepaper/content/201312-359.
[3] Lacher M D, Shiina M, Chang P, et al. ZEB1 limits adenoviral infectability by transcriptionally repressing the coxsackie virus and adenovirus receptor [J]. Mol Cancer, 2011, 10: 91.
[4] Ellis A L, Wang Z, Y u X, et al. Either ZEB1 or ZEB2/ SIP1 can play a central role in regulating the Epstein-Barr virus latent-lytic switch in a cell-type-specific manner [J]. J Virol, 2010, 84(12): 6139-6152.
[5] Feng W H, Kraus R J, Dickerson S J, et al. ZEB1 and c-Jun levels contribute to the establishment of highly lytic Epstein-Barr virus infection in gastric AGS cells [J]. J Virol, 2007, 81(18): 10113-10122.
[6] 古长庆, 金宁一. 鸡新城疫病毒抗肿瘤作用及其机制的研究进展[J]. 中国肿瘤生物治疗杂志, 2000, 7(3): 238-240.
[7] 冉静. 锌指转录因子ZEB1促进子宫内膜EMT在胚胎着床的功能及机制[D]. 广州: 南方医科大学, 2015.
[8] 余波. ZEB1蛋白在人骨肉瘤组织中表达及其与临床病理因素的相关性研究[D]. 长沙: 中南大学, 2012.
[9] 韩冰, 谢汝佳, 洪琴, 等. 肝纤维化病理过程中ZEB1和ZEB2的动态表达变化及意义[J]. 中国病理生理杂志, 2012, 28(9): 1639-1643.
[10] Jung S Y, Choi J H, Kwon S M, et al. Decursin inhibits vasculogenesis in early tumor progression by suppression of endothelial progenitor cell differentiation and function[J]. J Cell Biochem, 2012, 113(5): 1478-1487.
[11] Pan Y, Zhang J, Fu H, et al. miR-144 functions as a tumor suppressor in breast cancer through inhibiting ZEB1/2-mediated epithelial mesenchymal transition process[J]. Onco Targets Ther, 2016, 9: 6247-6255.
BIOINFORMATIC ANALYSIS OF CHICKEN ZEB1 AND ITS CORRELATION WITH NEWCASTLE DISEASE VIRUS INFECTION
WANG Wei1,2, ZHANG Yao-dan2, REN Ting-ting2, MENG Chun-chun2, TAN Lei2, SUN Ying-jie2, SONG Cui-ping2, LIAO Ying2, QIU Xu-sheng2, DING Chan2, LUO Ting-rong1
(1. College of Animal Science and Technology, Guangxi University, Nanning 530003, China; 2. Shanghai Veterinary Research Institute, CAAS, Shanghai 200241, China)
Zinc protein ZEB1(zinc finger E-box binding homeobox 1) is a transcription repressor, which is highly conserved in different species. To determine the correlation of ZEB1 with Newcastle disease virus(NDV) infection, the gene of chicken ZEB1 was PCR-amplified from DF-1 cells and used for phylogenetic and bioinformatic structure analysis. The expression of ZEB1 in NDV-infected DF1 cells, CEF cells and SPF chickens were examined in Real-time PCR. The results showed that NDV infection introduced high expression of chicken ZEB1, suggesting ZEB1 protein might play a role in the life cycle of NDV. Nevertheless, higher expression of ZEB1 did not affect the growth and protein expression of NDV. Therefore, the biological role of Zinc protein ZEB1 need to be further studied.
Newcastle disease virus; zinc protein; virus replication
S852.659.5
A
1674-6422(2017)02-0014-08
2016-06-06
公益性行业(农业)科研专项经费(201303033);国家自然科学基金(31101822,31530074)
汪伟,男,硕士研究生,预防兽医学专业
罗廷荣,E-mail:trluo@tom.com;仇旭升,E-mail:xsqiu1981@shvri.ac.cn
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
红蜻蜓·低年级(2022年5期)2022-05-11
河北农业大学学报(2022年2期)2022-04-26
中国畜牧杂志(2022年4期)2022-04-15
昆明医科大学学报(2022年1期)2022-02-28
幼儿园(2021年16期)2021-12-06
实用临床医药杂志(2021年13期)2021-01-10
实用肿瘤学杂志(2020年4期)2020-12-08
红领巾·探索(2020年5期)2020-05-19
家教世界·创新阅读(2020年12期)2020-01-11
