
中国瓜馥木属植物叶片结构的解剖观察
2017-05-30 10:48:04孙同兴董运秋侯学良
广西植物 2017年5期
孙同兴 董运秋 侯学良

摘要: 采用叶片表皮离析法、扫描电镜和石蜡制片法,研究了中国分布的番荔枝科瓜馥木属19种植物叶片的形态结构。结果表明:瓜馥木属叶片形态和结构具有较多的相似性,如叶片表皮均具有2-多个细胞的单列丝状毛,表皮细胞内具有一个晶簇,气孔器均为平列型,只分布在叶片的远轴面,远轴面的表皮细胞平周壁形成一个乳突,叶片主脉的维管组织除具有正常的维管组织外,还具有一个小的副维管束等,这些特征有助于区分番荔枝科植物的属间关系,支持瓜馥木属是一个很自然的类群。但叶片表皮毛的形态及分布,表皮细胞的形状、叶肉中栅栏组织和海绵组织的结构、远轴面的乳突大小以及叶主脉维管组织的细微结构则具有种间差异。尤其有助于区分小萼瓜馥木和黑风藤、广西瓜馥木和独山瓜馥木、上思瓜馥木和东方瓜馥木等形态相似、难以区分的植物种类。该研究结果为该属的系统研究和种间正确区分提供了重要资料。
关键词: 瓜馥木属, 番荔枝科, 叶, 形态结构, 比较解剖
中图分类号: Q945
文献标识码: A
文章编号: 10003142(2017)05057209
Abstract: Leaf morphology and structures of nineteen species of Fissistigma from China were studied using epidermal maceration, scanning electron microscope and paraffin section method. The results showed that the anatomical feathers of Fissistigma revealed a remarkable consistency. The leaf surface of Fissistigma were covered with bicellulate or multicellulate uniseriate trichomes. The epidermal cells contained one clustered crystal. The stomatal apparatus were paracytic with two to four subsidiary cells and only distributed on abaxial epidermis. The outer percilinal walls of abaxial epidermal cells were papllate. In the leaf midrib, there was one additional vascular strand except for normal vascular tissues. Therefore, these anatomical feathers should be contributed to differentiate the relationship among genera and support that Fissistigma is a natural group in Annonaceae. However, the form and distribution of trichomes, the shape of epidermal cells, the structures of mesophyll tissues, the sizes of papllate cells and the precise vascular structures in midrib vary obviously with species. Also, these anatomical differences between species may be used to distinguish some perplexing species such as F. minuticalyx and F. polyanthum, F. kwangsiense and F. cavaleriei, F. tungfangense and F. shangtzeense.
Key words: Fissistigma, Annonaceae, leaf, morphology and structure, comparative anatomy
瓜馥木屬(Fissistigma)隶属番荔枝科(Annonaceae)。蒋英和李秉滔(1979)认为该属植物有75种,Heusden(1992)认为有60种,Ban(2000)认为约50种。中国是瓜馥木属的现代分布中心,有24种(侯学良等, 2002)。瓜馥木属植物与我国其它番荔枝科植物不同,比较耐寒冷,分布广,种群数量大,是亚热带森林常见的成分,也是番荔枝科在亚洲分布的最北界,在东线可达28° N的浙江苍南,在西线可达29° N的西藏墨脱。
番荔枝科多数植物形态较为相似,属间分类较为困难。瓜馥木属(Fissistigma)由Griffith (1854)创立(蒋英和李秉滔,1965),但早期瓜馥木属植物曾被Hooker & Thomson(1855)错误地归于Melodorum Lour.属。Melodorum Lour.属由Loureiro(1790)建立,下属2种。Dunal(1817)把Melodorum降为Unona属下的一个组,包括Melodorum属下2个种和应为瓜馥木属的1个种。Blume(1830)又把Melodorum作为紫玉盘属Uvaria下的一个组。 Hooker & Thomson (1855)恢复了Melodorum属的地位。然而,Hooker & Thomson的Melodorum属概念却是Dunal(1817)作为Unona属下一个组的概念,并且排除了创建者Liureiro的最初两个种(Merrill, 1919; 将英和李秉滔, 1965)。实际上Hooker & Thomson的Melodorum属就是瓜馥木属,与Loureiro的Melodorum属无关。 Merrill(1919)对Melodorum属进行了考证,启用Fissistigma Griffith为正确名称,这种处理得到后来研究者(Sinclair, 1955; 蒋英和李秉滔, 1965, 1979; Heusden, 1992; Ban, 2000)的认可。但是,Melodorum属本身存在的分类问题使得刚被Merrill(1919)澄清的问题又扑朔迷离起来。Loureiro的Melodorum属的两个模式都丢失了,Merrill(1919)考证认为Melodorum fructicosum是该属的模式,另一个种M. abrrenus不是该属的成分,可能是银钩花属(Mitrephora)植物。可是后来,Merrill(1935)又考证说前边的处理明显是错误的,M. abrenus应是该属的模式,Melodorum fructicosum应是暗罗属(Polyalthia)的植物。Ban(1974)认为Melodorum fructicosum是Raunoffusus属植物。可见,由于Melodorum的模式问题不能解决,导致相关的属出现一些混乱。这对早期瓜馥木属的名称造成很大混乱,如:Fissistigma latifolium (Dunn) Merr.被Dunal(1817)置于Unona属,1830年Blume把它改为紫玉盘属Uvaria,1855年Hooker & Thomson 改为Melodorum属,1861年Miq又在Melodorum属发表它的晚出异名Melodorum mollissimum,另外还有Blume于1825年在Unona属发表了它的一个晚出异名Unona longifolia,这些错误名称的使用使得瓜馥木属的分类显得很混乱。
另外,导致瓜馥木属分类困难还有以下两个方面:一是瓜馥木属植物多数种类分布广、形态多样化,二是早期的研究基本源自欧洲零散的采集。如此以来,往往在标本较少的情况下发表的种,形态多样的种就被重复发表,在文献记述不详又加之标本采集不够,分类错误就在所难免。如:1919年Merrill组合了Uvaria bicolor为Fissistigma bicolor (Roxb) Merr.,1942年Merrill又组合了1938年Ast在Melodorum villosum为Fissistigma villosum, 后来证明二者同为一种。Merrill是这个属早期研究最多的一个学者,但是他的整理常常出现错误,增加了后来研究的难度。除了上述他先后指定Melodorum属最早的两个种作为属模式这样截然相反的表述外,他有时会犯下令人难以置信的“失误”。如:他1922年发表的Fissistigma maclurei则被蒋英作为蕉木Chieniodendron hainanense重新发表。
有关瓜馥木属植物的形态学研究已有一些报到,Walker(1971a, 1971b, 1972)、Waha & Morawetz(1980)描述了花粉的表面纹饰和花粉壁的结构,认为该属花粉很独特且属内也较为一致。Klucking(1986)详细研究了番荔枝科植物500多种的脉序,其中瓜馥木属植物16种。而有关叶片结构的研究仅在文献(Patel,1971,Bakker & Gerritsen,1992,lowokudejo,1990,Setten & KoekNoorman,1986,孙同兴等,2008)中包括部分种类。
本研究对中国分布的19种瓜馥木属植物叶结构进行观察研究,拟为该属的系统研究和种间正确区分提供重要资料。
1材料与方法
1.1 材料
本研究利用材料均取自FAA固定材料,来源和标本凭证见表1。腊叶标本存放于厦门大学植物标本馆(AU)。
1.2 方法
1.2.1 叶片表皮制片将FAA固定的材料切成5 mm2小片, 放入35% H2O2 和等量的醋酸溶液中, 于
60 ℃烘箱中放置24 h以上, 取出, 仔细剥离表皮, 5% H2O2 溶液漂白l0 min, 1%甘油封片。
1.2.2 扫描电镜制片选取叶片,切成10 mm2 小块,10% H2O2 溶液漂白l0 min,酒精梯度脱水后,用导电胶带黏贴在样品台上,镀膜喷金,在Philip XL30 ESEM扫描电镜下观察并照相。
1.2.3 石蜡制片将叶片中部切成5 mm2 小块, 经FAA 固定24 h以上, 常规石蜡切片法切片, 切片厚度6~8 μm, 番红-固绿对染。表皮制片和石蜡制片在LEICA DMIB 显微镜下观察照相。
2观察结果
2.1 光镜下叶片表皮的结构
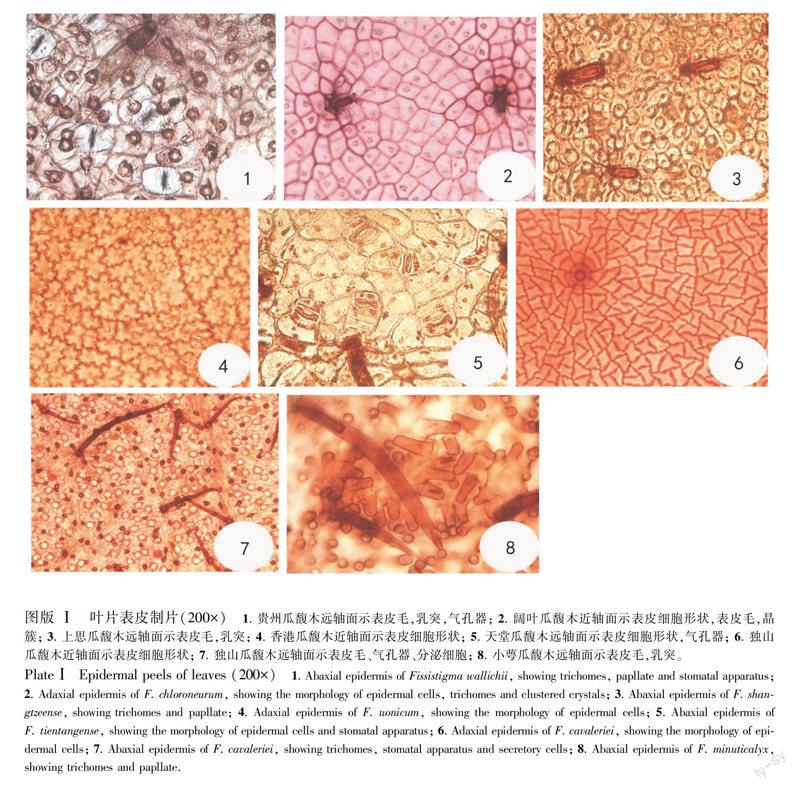
2.1.1 表皮细胞对19种植物叶片离析发现,瓜馥木属植物叶表皮细胞多为不规则排列,等径或不等径,长轴伸向各方向,两个表面的表皮细胞形状基本一致。贵州瓜馥木(图版Ⅰ:1)、阔叶瓜馥木(图版Ⅰ:2)、上思瓜馥木(图版Ⅰ:3)、小萼瓜馥木、多脉瓜馥木表皮细胞的垂周壁略直或浅波状。尖叶瓜馥木、金果瓜馥木、黑风藤、大果瓜馥木表皮细胞的垂周壁呈波状。白叶瓜馥木、香港瓜馥木(图版Ⅰ:4)、大叶瓜馥木、天堂瓜馥木(图版Ⅰ:5)、东方瓜馥木、毛瓜馥木、瓜馥木、广西瓜馥木、疣果瓜馥木的垂周壁为深波狀,而独山瓜馥木表皮细胞较为特殊,呈多边形或三角形(图版Ⅰ:6,7)。另外,瓜馥木属植物每个表皮细胞都含有草酸钙结晶,一般呈晶簇(图版Ⅰ:2),但大小差异较大。由于制片时许多细胞内的晶体有时会脱落,或照相时晶体与表皮细胞不在同一焦平面,经常看不到细胞内的晶体。特别注意的是瓜馥木属植物的远轴面表皮细胞比较特殊,外平周壁大多形成一个乳突,但乳突大小因种而具差异,阔叶瓜馥木、白叶瓜馥木、独山瓜馥木、东方瓜馥木、金果瓜馥木、广西瓜馥木、大果瓜馥木乳突较小,有时看不出来,而其它种类的乳突较为明显,其中贵州瓜馥木(图版Ⅰ:1)、小萼瓜馥木(图版Ⅰ:8)乳突特别突出。大叶瓜馥木、毛瓜馥木、小萼瓜馥木、独山瓜馥木(图版Ⅰ:7)叶远轴面表皮还具有分泌细胞存在, 分泌细胞通常呈圆形,细胞内的挥发性物质被提取后常呈空腔,或因贮存粘液物质而呈不透明。
2.1.2 气孔器19种瓜馥木属植物叶的气孔器排列不规则, 长轴伸向各个方向, 气孔器的大小和密度
因种而异。但整个属的气孔器结构极为相似,气孔器均在远轴面分布,为平列气孔型, 具有2~4 个副卫细胞(图版Ⅰ:1,5)。此外, 有许多种植物的叶片还具有双气孔型(图版Ⅰ:5)。
2.1.3 表皮毛瓜馥木属植物叶表面普遍具有表皮毛,表皮毛类型为由2-多个细胞构成的单列丝状毛, 单列丝状毛的基细胞通常较短且木质化, 而顶细胞壁较薄、细长而尖(图版Ⅰ:1-3,5,7,8)。其中阔叶瓜馥木、尖叶瓜馥木、独山瓜馥木(图版Ⅰ:6)、东方瓜馥木、毛瓜馥木、金果瓜馥木、广西瓜馥木、大果瓜馥木、疣果瓜馥木、小萼瓜馥木(图版Ⅰ:8)远轴面表皮毛密度较大,这些种通常在近轴面也有少量的表皮毛分布(图版Ⅰ:2,6),而其它种类的表皮毛仅分布在叶的远轴面。
2.2 扫描电镜下叶片的表面特征
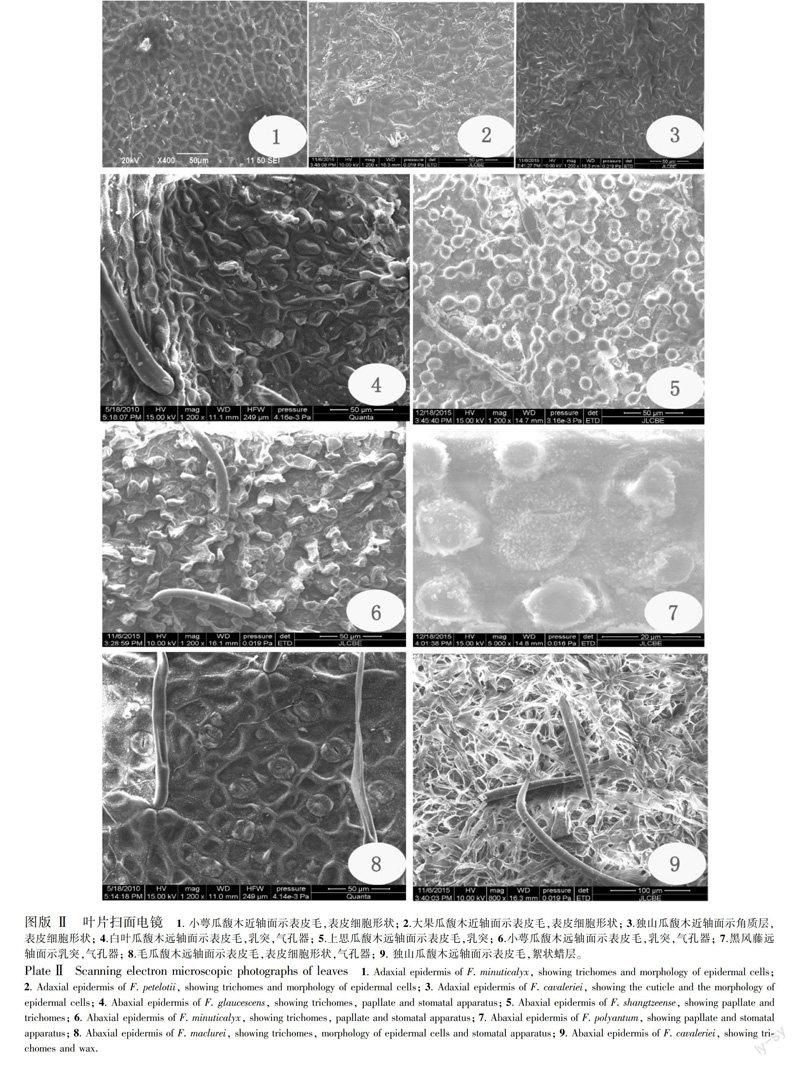
瓜馥木属植物叶的表面由于具有较厚的角质层,所以部分种类的表皮细胞轮廓不清。近轴面表皮细胞能看清的种类有阔叶瓜馥木、白叶瓜馥木、尖叶瓜馥木、大叶瓜馥木、天堂瓜馥木、小萼瓜馥木(图版Ⅱ:1)、大果瓜馥木(图版Ⅱ:2),它们的表皮细胞垂周壁直或波状,有的凸起,如白叶瓜馥木、小萼瓜馥木(图版Ⅱ:1),有的凹陷,如大果瓜馥木(图版Ⅱ:2),而独山瓜馥木近轴面在扫描电镜下也较为特殊,外面的角质层堆积在垂周壁上,也能显示出表皮细胞较小,多角形(图版Ⅱ:3)。瓜馥木属植物叶远轴面普遍具有乳突,阔叶瓜馥木、白叶瓜馥木(图版Ⅱ:4)、大叶瓜馥木、东方瓜馥木、金果瓜馥木、广西瓜馥木、大果瓜馥木乳突略不明显,而其它种类乳突较为明显,特别是上思瓜馥木(图版Ⅱ:5)、小萼瓜馥木(图版Ⅱ:6)、黑风藤的乳突极为明显,乳突较长,有时相邻的乳突还会连成串,如小萼瓜馥木(图版Ⅱ:6)。远轴面的气孔器一般由于乳突的凸起,而显得气孔是凹陷的(图版Ⅱ:5,6,7),而在乳突不明显的种类,气孔可以和表皮细胞平或略凸。表皮毛多分布于远轴面,贵州瓜馥木、香港瓜馥木、天堂瓜馥木、黑风藤、瓜馥木的表皮毛稀少,而其它种类的表皮毛较多(图版Ⅱ:8),毛瓜馥木、金果瓜馥木、广西瓜馥木、小萼瓜馥木(图版Ⅱ:1)近轴面也有少量的表皮毛。这些毛通常较直,而东方瓜馥木、广西瓜馥木的表皮毛较为弯曲。在扫描电镜下有时能看到表皮毛是由2个细胞构成的(图版Ⅱ:5,8)。独山瓜馥木叶远轴面除具表皮毛外,还具有较厚的絮状蜡层(图版Ⅱ:9),由于这些蜡层的存在仅使气孔器隐约可见。
2.3 叶的解剖结构
瓜馥木属19种植物叶片的结构具有极大的相似性, 但又存在着明显的种间差异。从叶片横切面观察, 结果如下。
2.3.1 表皮瓜馥木属19种植物叶表皮均具有较厚的角质层,多数种具一层表皮细胞(图版Ⅲ:1-3,5,9-11)。表皮细胞大多为方形或长方形。独山瓜馥木(图版Ⅲ:4)、上思瓜馥木(图版Ⅲ:7)和小萼瓜馥木(图版Ⅲ:12)叶的近轴面有两层表皮细胞,即复表皮,其中外层细胞较小,正方形或圆形,内层细胞较大,长方形。近轴面和远轴面的表皮细胞内都具有一个簇晶,簇晶的大小因种而异(图版Ⅲ:9,12)。瓜馥木近轴面表皮细胞有两种类型,具大晶簇的细胞和具小晶簇的细胞(图版Ⅲ:9)。瓜馥木属植物叶远轴面表皮细胞的外平周壁明显向外突起而形成乳突, 且一个细胞形成一个乳突, 但乳突的大小也因植物种类而异,阔叶瓜馥木(图版Ⅲ:2)、白叶瓜馥木(图版Ⅲ:3)、大叶瓜馥木、独山瓜馥木(图版Ⅲ:4)、东方瓜馥木(图版Ⅲ:6)、金果瓜馥木、瓜馥木(图版Ⅲ:9)、广西瓜馥木(图版Ⅲ:10)、大果瓜馥木乳突不太明显,其它种类瓜馥木属植物叶远轴面乳突较为明显,而贵州瓜馥木(图版Ⅲ:1)、上思瓜馥木(图版Ⅲ:7)、毛瓜馥木(图版Ⅲ:8)、小萼瓜馥木的乳突尤为突出。
2.3.2 叶肉组织瓜馥木属植物叶片为异面叶, 叶肉组织明显分化为栅栏组织和海绵组织,通常栅栏组织细胞1~ 2 层, 栅栏组织和海绵组织的比值为1∶1~2,而上思瓜馥木(图版Ⅲ:7)、毛瓜馥木(图版Ⅲ:8)、疣果瓜馥木(图版Ⅲ:11)、小萼瓜馥木可达2层以上层, 栅栏组织和海绵组织比值则为2~3∶1。叶片厚度一般决定于栅栏组织的层数或栅栏组织细胞的长度,它们的栅栏组织中通常第1 层栅栏组织细胞长圆柱形, 细长(图版Ⅲ:5,12), 而第2、3 层栅栏组织细胞可呈长圆柱形或呈短棒状, 有的排列较疏散, 略不规则(图版Ⅲ:2),贵州瓜馥木的1层栅栏组织细胞较短,栅栏组织和海绵组织比值则为1∶4左右(图版Ⅲ:1)。瓜馥木属植物叶肉组织中均普遍具有油细胞分布(图版Ⅲ:5), 由于在石蜡制片过程时, 油细胞中的挥发性物质常被提取, 因此, 叶片横切面中, 成熟油细胞常呈圆形或椭圆形, 其细胞壁染色较深, 大部分种类的细胞质己完全解体, 仅剩一个较大的空油囊充满整个细胞腔,油细胞比周围其它类型的细胞大(图版Ⅲ:5),白叶瓜馥木(图版Ⅲ:3)、天堂瓜馥木、广西瓜馥木(图版Ⅲ:10)叶肉中油细胞的分布数量明显大于其它种类。
2.3.3 主脉瓜馥木属19种植物叶主脉周围的薄壁组织中具有分枝状石细胞和油细胞。维管组织极为发达,大都呈完整的一环, 木质部位于内方, 韧皮部位于外方, 多数种类的主脉维管组织被较宽的薄壁或木质化的细胞分隔成束状, 包围其外的维管束鞘也呈束状,这些束状结构有时可达10多个,如贵州瓜馥木(图版Ⅲ:13)、白叶瓜馥木(图版Ⅲ:14)、天堂瓜馥木、广西瓜馥木(图版Ⅲ:21)、疣果瓜馥木,仅瓜馥木(图版Ⅲ:20)、金果瓜馥木、尖叶瓜馥木(图版Ⅲ:15)等维管束数目较少,而东方瓜馥木(图版Ⅲ:17)和天堂瓜馥木的木质部较为发达,其间的薄壁组织不发达。瓜馥木属植物叶的主脉维管束较特殊的是除金果瓜馥木和瓜馥木(图版Ⅲ:20)不明显外,其他17种植物叶的主脉维管组织除了正常的维管束外,还形成一个较小的副维管束,即在靠近近轴面的薄壁组织中还具有由木质部、韧皮部和形成层构成的小维管组织, 该小维管组织由一环状
形成层产生, 韧皮部在内方(远轴面),而木质部在外方(近轴面)形成一环(图版Ⅲ:13-19,21) ,这个小的副维管束也可以是连续的,如毛瓜馥木、小萼瓜馥木(图版Ⅲ:19)的小副维管束木质部之间连续,而贵州瓜馥木(图版Ⅲ:13)、白叶瓜馥木(图版Ⅲ:14)、上思瓜馥木(图版Ⅲ:18)、广西瓜馥木(图版Ⅲ:21)等副维管束由多个小的维管束构成。
3讨论
番荔枝科植物外部形态(蒋英和李秉滔,1979)、木材解剖特征(Vander & Canright, 1956) 和叶片表皮结构(Patel, 1971)、花粉的表面纹饰和壁的结构(Walker,1971a, 1971b, 1972;Waha & Morawetz,1980)、種子的结构(Heijden & Bouman,1988)及植物体内均含有番荔枝内酯都显示了番荔枝科植物为一个非常自然的类群,具有很大的相似性,各属间的形态差异较少,以致于某些属的界限很难确定。而通过解剖结构发现,19种植物叶片形态结构表明瓜馥木属植物具有很大的相似性,具有属级水平的共性。如叶的表面均具有单列丝状表皮毛,表皮毛的基本结构也十分相似,基细胞短且木质化,顶细胞细长而尖,而番荔枝科的紫玉盘属(Uvaria)均具有星状毛、哥纳香属(Goniothalamus)具有短的单列毛和盾状毛两种类型(孙同兴等,2008)。瓜馥木属植物叶表皮细胞内晶体类型均为晶簇,而澄广花属(Orophea)、野独活属(Muliusa)、银钩花属(Mitrephora)和暗罗属(Polyalthia)等主要含有棱晶(孙同兴等,2008),瓜馥木属植物叶远轴面表皮细胞普遍形成乳突, 而其它番荔枝科植物叶远轴面具乳突仅在假鹰爪属(Desmos)、皂帽花属(Dasymaschalon)、哥纳香属及暗罗属的少数种类中存在(Patel,1971;Metcalfe,1987;孙同兴等,2008)。另外,在番荔枝科中,主脉中除具有正常的维管组织外还具有一个副维管束则是瓜馥木属独特的特征(Metcalfe,1987)。叶脉序研究也表明,瓜馥木属植物叶侧脉多而明显,近平行斜升至叶缘,在番荔枝科也属于较为独特的一类(Klucking,1986)。
本文所研究的19种植物叶表皮细胞的形态、表皮毛的密度、叶肉组织和主脉的结构等特征存在的种间差异也有助于瓜馥木属植物的种间分类。如白叶瓜馥木、香港瓜馥木、金果瓜馥木、黑风藤、天堂瓜馥木、东方瓜馥木、毛瓜馥木、瓜馥木、广西瓜馥木、疣果瓜馥木10种植物叶表皮细胞垂周壁呈深波状,而其它种类叶表皮细胞垂周壁浅波状或波状。而独山瓜馥木叶表皮细胞较为特殊,多边形或三角形。大叶瓜馥木、毛瓜馥木、小萼瓜馥木、独山瓜馥木叶远轴面还具有圆形油细胞存在。瓜馥木属植物也普遍含有晶簇,但晶簇的大小也因种而已,而瓜馥木叶近轴面表皮细胞有两种细胞,具大晶簇的细胞和具小晶簇的细胞。瓜馥木属植物叶远轴面形成的乳突大小也不一致,阔叶瓜馥木、白叶瓜馥木、独山瓜馥木、东方瓜馥木、金果瓜馥木、广西瓜馥木、大果瓜馥木乳突较小,而贵州瓜馥木、上思瓜馥木、小萼瓜馥木、黑风藤乳突则极为突出。Metcalfe(1987)记载了瓜馥木属9种植物乳突,其中7种较长,2种较短。另外,瓜馥木属叶表皮毛的密度和分布也因种而异,其中尖叶瓜馥木、独山瓜馥木、东方瓜馥木、毛瓜馥木、金果瓜馥木、广西瓜馥木、大果瓜馥木、疣果瓜馥木、小萼瓜馥木叶远轴面表皮毛密度较大,近轴面也分布少量的表皮毛,而其它种植物的表皮毛仅分布在叶远轴面。大多数种类的表皮毛较直,而东方瓜馥木、广西瓜馥木的表皮毛较为弯曲。独山瓜馥木叶远轴面除具表皮毛外,还具有较厚的絮状蜡层。瓜馥木属多数种类叶仅1层表皮细胞,而独山瓜馥木、上思瓜馥木和小萼瓜馥木叶的近轴面有2层表皮细胞。叶肉组织中栅栏组织细胞形状和层数也具有差别,多数种类的栅栏组织细胞1~2 层, 而上思瓜馥木、毛瓜馥木、疣果瓜馥木、小萼瓜馥木可达2层以上,栅栏组织和海绵组织的比值则因种而异。瓜馥木属叶主脉维管组织也存在种间变化,多数种类的主脉维管组织被分割成束状,如贵州瓜馥木、白叶瓜馥木、天堂瓜馥木、广西瓜馥木、疣果瓜馥木维管束的数目较多,仅瓜馥木、金果瓜馥木、尖叶瓜馥木等维管束数目较少,而东方瓜馥木和天堂瓜馥木的木质部连续成环状,其间的薄壁组织不发达。瓜馥木属植物叶的主脉维管组织是番荔枝科较为独特的类型,主脉维管组织除了正常的维管束以外,在近轴面处还具有一个副维管组织,该副维管组织由一环状形成层产生, 韧皮部在内方,木质部在外方形成一环(Metcalfe,1987)。这个副维管组织的细致结构也具种间区别,在19种植物中,金果瓜馥木和瓜馥木的副维管组织较不明显,毛瓜馥木、小萼瓜馥木的副维管组织维管束木质部之间连续,而白叶瓜馥木、广西瓜馥木、贵州瓜馥木、上思瓜馥木等种类副维管组织之间的薄壁组织较为发达,把维管组织分隔成多个小的维管束。
中國植物志记载了瓜馥木属多种植物形态相似(蒋英和李秉滔,1965,1979),如在所研究的19种中,小萼瓜馥木与黑风藤较为相似,广西瓜馥木与独山瓜馥木较为相似,上思瓜馥木和东方瓜馥木较为接近(蒋英和李秉滔,1965)。而观察叶的解剖结构,小萼瓜馥木近轴面表皮细胞2层,远轴面表皮具有油细胞,栅栏组织和海绵组织的比值较大,这些特征可以有助于小萼瓜馥木和黑风藤的区分。独山瓜馥木叶结构比较特殊,表皮细胞形状多边形或三角形,近轴面表皮细胞2层,远轴面除具表皮毛外,还具有较为特殊的蜡层结构,这些特征可以有助于区分广西瓜馥木和独山瓜馥木。上思瓜馥木近轴面表皮细胞2层,栅栏组织细胞2层以上,与海绵组织的比值较高,远轴面表皮细胞乳突极为明显等特征也有助于同东方瓜馥木的区分。
参考文献:
AST M,1938. Supplement a la flora cenerale de L Indochina.1. [M]. Paris: Tome Proliminate: 59-123.
BAKKER ME, GERRITSEN AF, 1992. Oil and mucilage cells in Annona (Annonaceae) and their systematic significance [J]. Blumea,36(2):411-438.
BAN NT,1974. Critical notes on the genera Melodorum Lour., Mitrella Miq. and Rauwenhoffia Scheff. (Annonaceae Juss. ). Bot Zhurn,
59(2): 237-245.
BAN NT,2000. Flora of Vietnam [M]. Ha Nl: Science & Technics Publishing House: 1-321.
BLUME CL,1825. Bijdragen tot de flora van NederlandschIndi [M]. Batavia: 11-21.
BLUME CL,1830. Flora Javae [M]. Brussels:J Frank: 1-108.
BLUNDEN G, AYE K, JEWERS K,1973. The comparative leaf anatomy of Goniothamus andersonii, G. macrophyllus, G. malayanus and G. velutinus [J]. Bot J Linn Soc,67:361-376.
DUNAL MF,1817. Monographie de la famille des Anonacéae [M]. Paris:Treuttel and Wüirtz: 110-112.
HEIJDENE V, BOUMAN F,1988. Studies in Annonaceae. X:Seed anatomy of the Annona group [J]. Bot Jahrb Syst,110(1):117-135.
HEUSDENECH V, 1992. Flowers of Annonaceae:Morphology classification and evolution [J]. Blumea(Suppl),7:1-216.
HOOKER JD, THOMSON T, 1855. Flora Indica I. [M]. London: W Pamplin: 86-153.
HOU XL,ZHU SX, LI PT, 2002. A newly recorded species of Fissistigma (Annonaceae) from China [J]. Acta Phytotax Sin ,40(6): 546-548. [侯学良,朱世新,李秉滔,2002. 中国瓜馥木属(番荔枝科)一新记录 [J]. 植物分类学报,40(6): 546-548.
HUSSIN KH,SAMAH NA, MATSALLEH K, 2000. Comparative leaf anatomy of Uvaria Linn. , Cyathostemma Griff. and Ellipeia Hook. f. et Thomson (Annonaceae) from Malaysia [J]. J Trop Subtrop Bot, 8(3):215-224.
JING Y, LI BT, 1979. Flora Reipublicae Popularis Sinicae: Vol. 30(2) [M]. Beijing: Science Press: 10-175. [蒋英,李秉滔,1979. 中国植物志,第 30 卷第 2 分册 [M]. 北京:科学出版社,10-175.]
KLUCKING EP,1986. Leaf venation patterns 1: Annonaceae [M]. Berlin, Stuttgart: J Cramer: 1-256.
LOUREIRO J DE,1790. Flora Cochinchinensis I. [M]. Lissabon: Typis. et Expensis Academicus: 352-353.
MERRILL ED,1919. On the application of the generic name Melodorum of Loureiro. [J]. Philipp J Sci Bot,15:125-137.
MERRILLED,1922. Dingnoses of Hainan plants [J]. Philipp J Sci Bot, 21(4):337-342.
MERRILL ED,1935. A commentary on Loureiros “Flora Cochinchinensis” [J]. Trans Amer Philos Soc New Ser, 24(2):160-162.
METCALFE CR,1987. Anatomy of the Dicotyledons [M]. 2nd ed. Oxford: Clarendon Press,3: 34-48.
OLOWOKUDEJO JD,1990. Comparative morphology of leaf epidermis in the genus Annona (Annonaceae) in West Africa [J]. Phytomorphology,40(3, 4):407-422.
PATEL RJ,1971. Epidermal structure and development of stomata in some Annonaceae [J]. Ann Bot, 35:1205-1212.
SETTEN AK V, KOEKNOORMAN J,1986. Studies in Annonaceae. VI. A leaf anatomical survey of genera of Annonaceae in the Neotropics [J]. Bot Jahrb Syst,108(1):17-50.
SINCLAIRJ, 1955. A revision of the Malayan Annonaceae [J]. Gard Bull Sing,14: 149-516.
SUN TX, WU H, LI PT,2008. Comparative anatomy on leaves of Annonaceae [J]. Acta Bot Yunnan, 30(1):19-37. [孫同兴,吴鸿,李秉滔,2008. 番荔枝科植物叶的比较解剖学研究 [J]. 云南植物研究, 30(1):19-37.
TSIANG Y, LI PT,1965. Reviso specierum sinensium Fissistigma Griffith [J]. Acta Phytotax Sin, 10(4):315-328. [蒋英, 李秉滔,1965. 中国瓜馥木属植物的修订 [J]. 植物分类学报, 10(4): 315-328.
VANDER WRW,CANRIGHT JE,1956. The anatomy and relationships of the Annonaceae [J]. Trop Woods, 104:1-24.
WAHA M,MORAWETZ W,1980. Pollen evolution and systematics Annonaceae with special reference to the disulcate Australian endemic genera [J]. Plant Syst Evol,161:1-12.
WALKER JW,1971a. Pollen morphology,phytogeography and phylogeny of the Annonaceae [J]. Contrib Gray Herb Harvard, 202:1-132.
WALKER JW,1971b. Contributions to the pollen morphology and phylogeny of the Annonaceae I. [J]. Grana, 11:45-54.
WALKER JW,1972. Contributions to the pollen morphology and phylogeny of the Annonaceae II. [J]. Bot J Linn Soc,65: 173-178.
