
黄芪总黄酮对柯萨奇B3病毒感染乳鼠心肌细胞miRNA378和miRNA378*表达的影响*
2017-05-20 02:26崔晓雪冯静宜刘永峰
中国应用生理学杂志 2017年1期
万 莹, 王 羽, 崔晓雪, 冯静宜, 龙 杰, 刘永峰△, 赵 明
(1. 内蒙古民族大学, 2. 内蒙古民族大学附属医院, 通辽 028000)
黄芪总黄酮对柯萨奇B3病毒感染乳鼠心肌细胞miRNA378和miRNA378*表达的影响*
万 莹1, 王 羽1, 崔晓雪1, 冯静宜1, 龙 杰1, 刘永峰1△, 赵 明2△
(1. 内蒙古民族大学, 2. 内蒙古民族大学附属医院, 通辽 028000)
目的:研究柯萨奇B3病毒(CVB3)感染乳鼠心肌细胞miRNA378和miRNA378*表达的作用。方法:原代培养乳鼠心肌细胞分3组(n=6):对照组(正常细胞)、CVB感染组(正常细胞+CVB3)、黄芪总黄酮组(正常细胞+CVB3+黄芪总黄酮)。CVB感染组感染CVB3,黄芪总黄酮组感染CVB3同时给予黄芪总黄酮20 mg/L。 采用免疫组化方法检测乳鼠心室肌细胞α-SMA蛋白,实时荧光定量PCR技术检测各组心肌细胞miRNA378及miRNA378*表达。结果:①与对照组相比较,CVB感染组心肌细胞miRNA378及miRNA378*表达明显减少(P<0.01);②与CVB感染组比较,黄芪总黄酮心肌细胞miRNA378及miRNA378*表达明显增加(P<0.01)。结论:黄芪总黄酮可以减少CVB3感染心肌细胞miRNA378及miRNA378*表达。
柯萨奇B3病毒;黄芪总黄酮;乳鼠;miRNA378;miRNA378*
coxsackie virus B3; total flavonoids of astragalus; suckling mouse myocardium, miRNA378; miRNA378*
【DOI】 10.12047/j.cjap.5379.2017.013
病毒性心肌炎(viralmyocarditis,VMC)是由病毒(主要由柯萨奇病毒引起,另有腺病毒、肠道病毒、EB病毒、人疱疹病毒、细小病毒B19和巨细胞病毒等)侵犯心脏引起的心肌实质或间质的局限性或弥漫性病变[1],其中心肌组织严重而广泛的炎症反应导致心肌收缩力急剧下降引起心力衰竭,是引起患者死亡的重要原因[2,3]。microRNAs (miRNAs)是一类长约22个核苷酸的非编码小分子RNA,通过与靶蛋白mRNA 3' 端非编码区的不完全互补结合,抑制靶mRNA 转录后的表达。研究表明miRNA参与心肌发育、心肌肥大等病理过程[4]。miRNA-378和miRNA-378*的产物,叫正义链和反义链,microRNA前体有发卡结构 在成熟过程中发卡结构被
剪掉 形成miRNA-378和miRNA-378*,也叫左臂和右臂[6]。文献报道:miRNA-378 和miRNA-378*调控心肌重塑与心力衰竭发生密切相关[5]。黄芪总黄酮(total flavonoids of Astragalus,TFA)是从蒙古黄芪中分离出的主要活性成分[7]。本研究团队在前期研究中发现TFA明显改善柯萨奇B3病毒(coxsackievirus B3,CVB3)所致病毒性心肌炎小鼠心肌病理改变和超微结构[8],但作用机制尚未明确。本研究拟观察TFA对CVB3感染乳鼠心肌细胞miRNA378及miRNA378*表达影响,阐明TFA改善CVB3所致病毒性心肌炎小鼠的发病机制。
1 材料与方法
1.1 动物药品与仪器
1~3 d Balb/c新生乳鼠购置吉林大学基础医学院动物中心,许可证号:scxk(吉)2011-0004; Hela细胞由内蒙古民族大学医学院提供。Super M-MLV 反转录酶(BioTeke)公司, RNA simple Total RNA Kit试剂盒(TIANGEN)公司,引物由生工生物工程(上海)有限公司合成;:CVB3,Woodruf亲心肌株由吉林大学农牧学院提供。
1.2 柯萨奇B3病毒感染心肌细胞模型制备
1.2.1 病毒扩增 人宫颈癌细胞(Hela细胞)用含10%小牛血清的DMEM 培养液培养, 在对数生长期接种病毒,当90% 的细胞出现细胞病变效应后, 收集病毒液, 重复3次, 病毒悬液分装于-80℃保存。用Reed-Mnench法测定病毒滴度。
1.2.2 乳鼠心肌细胞的分离与培养 将乳鼠心脏放入培养皿中,用PBS反复清洗。将清洗后的心脏组织剪碎至1~3 mm3小块。加入0.1%Ⅱ型胶原酶以及0.1%胰蛋白酶混合液,37℃消化25 min。消化好后将其移入15 ml离心管1 500 r/min离心7 min去上清。用PBS重悬沉淀,用吸管反复吹打后1 000 r/min离心5 min,用培养基重悬细胞,过200目筛网去除大块组织。PBS清洗细胞2次,1 000 r/min离心10 min,去上清,留沉淀。加入1 ml的培养基重悬细胞,细胞计数后放置到培养板中。成纤维细胞贴壁较快,2 h差速贴壁法可去除大量的成纤维细胞,上清悬液中为较纯的心肌细胞。细胞静置培养24 h,随后每2天换一次液。培养细胞至密度为90%左右,PBS清洗2次。按实验内容将细胞接种于6孔板中,细胞浓度为5×104cells/ml,置于37℃,5% CO2的培养箱内培养过夜。24 h可进行转染,细胞密度一般在70%为宜。
1.2.3 免疫组织化学方法鉴定乳鼠培养心肌细胞 各组乳鼠培养心肌细胞进行免疫组化鉴定:0.1%TritonX-100孵育;3%过氧化氢孵育;血清封闭;一抗孵育;二抗孵育;辣根酶标记;DAB显色;苏木素复染;镜检。
1.2.4 黄芪总黄酮对感染CVB3的乳鼠心肌细胞作用的观察 乳鼠心肌细胞按照细胞浓度为5×104cells/ml接种于6孔板,并将其置于37℃,5% CO2培养箱内培养,心肌细胞随机分成3组,每组3孔,取2组细胞与100 PFU的CVB3共同孵育1 h,弃上清后,其中一组加入正常培养液作为CVB感染组;另一组加入正常培养液,并加入黄芪总黄酮20 mg/L作为黄芪总黄酮组(剂量参照之前的验证)[8];设立细胞对照组(仅加入培养液,不进行病毒感染),将3组心肌细胞在37℃,CO2培养箱中培养3 d。
1.3 Real time PCR 检测各组心肌细胞miRNA378及miRNA378*表达
经下列实验步骤:样本总RNA提取;RNA浓度检测;反转录;荧光定量PCR检测。每种样本每个基因进行4个复孔平行实验,实验重复3次(n=3),实验数据的选取,是舍去误差较大的数值,取剩余数值的平均值作为最终实验保留数据,利用2-△△CT方法分析数据(表1)。

Tab. 1 CVB injured cardiomyocytes model preparation
OD260/OD280: Purity of DNA was assessed; CVB: Coxsackievirus B; AST: Astragalus
1.4 统计学方法
2 结果
2.1 各组乳鼠培养心肌细胞免疫组化
ɑ-SMA是心肌细胞表达的蛋白质,是心肌细胞可靠的标志蛋白。为检测分离的细胞为乳鼠心肌细胞,通过免疫组化检测ɑ-SMA蛋白,各组均检测出ɑ-SMA,确定3组细胞都是心肌细胞(图1)。
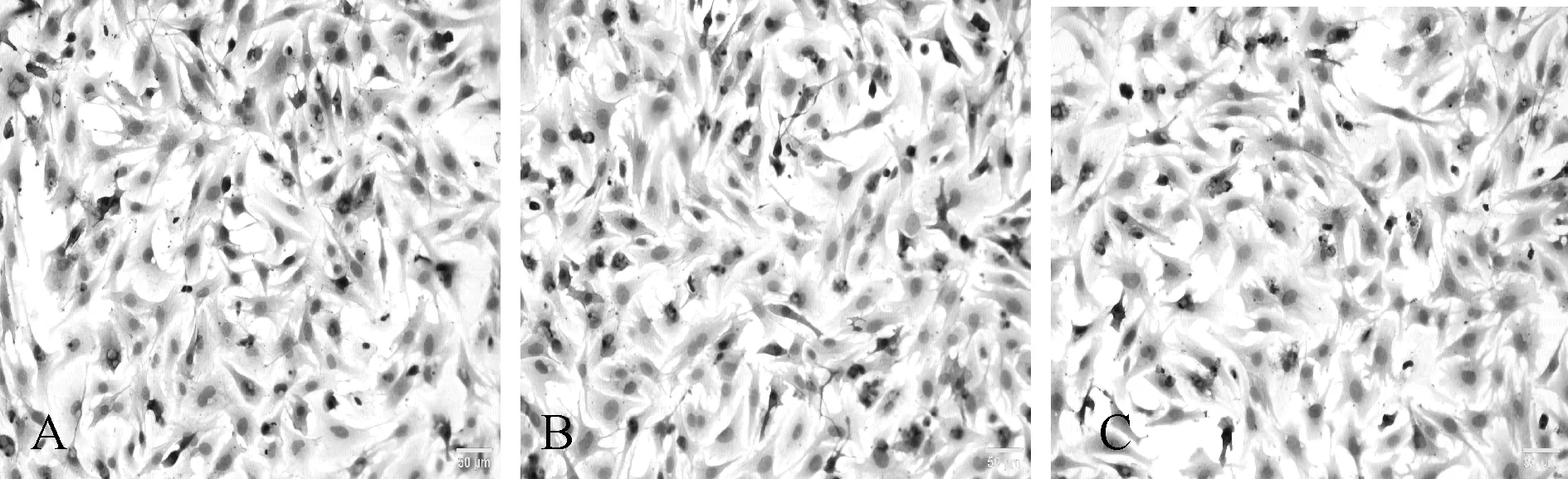
Fig. 1 Immunocytochemistry of culturing myocardial cell (×200) A: Control group; B: Coxsackievirus B3 infecton group; C: Total flavonoids of astragalus group
2.2 real time PCR 检测各组心肌细胞miRNA378和miRNA378*表达:
与对照组比较,柯萨奇病毒感染组心肌细胞miRNA378和miRNA378*表达明显减少(P<0.01)。与CBV3感染组相比较,黄芪总黄酮心肌细胞miRNA378和miRNA378*表达明显增加(P<0.01,表2)。

GroupmiRNA378miRNA378*Control1.00±0.021.00±0.03CVB0.76±0.01**0.56±0.02**AST0.94±0.01##0.78±0.02##
CVB: Coxsackievirus B; AST: Astragalus
**P<0.01vscontrol group;###P<0.01vsCVB group
3 讨论
心脏重构是心脏在对抗外界因素所致的压力和阻力时为了维持稳态而进行自身调节的适应性过程,如果所致压力和阻力作用持续存在,这种过程可发展为不可逆作用,同时在细胞和细胞外水平可发生凋亡、坏死和纤维化等变化。本团队发现病毒性心肌炎时内质网应激介导了心肌细胞凋亡,并且黄芪总黄酮对病毒性心肌炎心力衰竭小鼠内质网应激介导的心肌细胞凋亡有保护作用。文献报道:miRNA-133、miRNA-1、miRNA-21、miRNA-208、miRNA-29等都参与心肌重构,而且miRNA-146b直接参与病毒性心肌炎的病理学发病过程[4]。
研究发现miRNA-378通过影响转化因子β1(TGFβ-1)分泌而参与心肌纤维化过程[9],抑制miRNA-378表达能诱导小鼠心肌纤维化[10],重度心力衰竭患者miRNA-378 和miRNA-378*表达显著下降。随着患者心功能好转,miRNA-378 和miRNA-378*表达也明显增加[11]。黄芪总黄酮通过干预病毒性心肌炎miRNA-378 和miRNA-378*表达来影响心肌重构,进而改善病毒性心肌炎导致的心脏功能下降。
本实验观察到黄芪总黄酮通过干预病毒性心肌炎miRNA-378 和miRNA-378*表达来影响心肌重构,进而改善病毒性心肌炎导致的心脏功能下降。揭示了黄芪总黄酮对病毒性心肌炎作用产生机制,为传统药物走向现代化提供了理论基础。而miRNA378及miRNA-378*表达是如何调控病毒性心肌炎所致心肌重构还需在以后的实验中进一步阐明。
[1] Lv S, Rong J, Ren S,etal. Epidemiology and diagnosis of viral myocarditis[J].HellenicJCardiol, 2013, 54(5): 382-391.
[2] Wei J, Gao DF, Wang H,etal. Impairment of myocardial mitochondria in viral myocardial disease and its reflective window in peripheral cells[J].PLoSOne, 2014, 9(12): e116239.
[3] 谭振宇, 杨大浩, 廖春燕. 主动脉内球囊反搏对急性重症心肌炎心力衰竭患者室性心律失常的影响[J]. 岭南心血管病杂志, 2013, 19(5): 565-567.
[4] 李歆跃, 杨 巍. 心脏重构中微RNA 的作用[J]. 医学综述, 2015, 21(2): 582-584.
[5] Lok SI, de Jonge N, van Kuik J,etal. MicroRNA Expression in Myocardial Tissue and Plasma of Patients with End-Stage Heart Failure during LVAD Support: Comparison of Continuous and Pulsatile Devices[J].PloSOne, 2015, 10(10): e0136-404.
[6] 赵 明, 单晓彤, 赵亚红, 等. 阿霉素损伤心肌细胞miRNA-378 和miRNA-378*与calumenin,GRP78,bax及bal-2相关性研究[J]. 临床心血管病志, 2016, 32(6): 603-606
[7] Zhou X, Xin Q, Wang Y,etal. Total Flavonoids of Astragalus Plays a Cardioprotective Role in Viral Myocarditis[J].ActaCardiolSin, 2016, 32(1): 81-88.
[8] 巴金华, 窦忠霞, 柳文清, 等. 黄芪总黄酮对柯萨奇B_3病毒所致病毒性心肌炎小鼠心肌病理改变和超微结构的影响[J]. 中国心血管病究, 2013, 11(11): 910-912.
[9] Nagalingam RS, Sundaresan NR, Noor M,etal. Deficiency of cardiomyocyte-specific microRNA-378 contributes to the development of cardiac fibrosis involving a transforming growth factor β (TGFβ1)-dependent paracrine mechanism [J].JBiolChem, 2014, 289(39): 27199-27214.
[10]Nagalingam RS, Sundaresan NR, Gupta MP,etal. A cardiac-enriched microRNA, miR-378, blocks cardiac hypertrophy by targeting Ras signaling[J].JBiolChem, 2013, 288(16): 11216-11232.
[11]Ooi JY, Bernardo BC, McMullen JR. The therapeutic potential of miRNAs regulated in settings of physiological cardiac hypertrophy[J].FutureMedChem, 2014, 6(2): 205-222.
国家自然科学基金课题(81360587)
2015-11-16 【修回日期】2016-10-19
R331.3;R332
A
1000-6834(2017)01-055-03
△【通讯作者】Tel: 13474859991; E-mail: langzhe73@163.com, dr.liudanial@gmail.com
猜你喜欢
湖南饲料(2021年4期)2021-10-13
食品安全导刊(2021年21期)2021-08-30
华声文萃(2021年6期)2021-08-25
文萃报·周二版(2021年11期)2021-04-06
四川蚕业(2021年2期)2021-03-09
四川蚕业(2021年3期)2021-02-12
保鲜与加工(2021年1期)2021-02-06
家庭用药(2017年3期)2017-04-17
妇女生活(2016年1期)2016-01-14
中国民族民间医药·下半月(2014年2期)2014-09-26
