
毛竹凋落叶生态化学计量特征季节变化及其对氮沉降的响应
2017-05-17 08:50翁俊,徐璐
山东林业科技 2017年3期
翁 俊 ,徐 璐
(江苏省泰州市兴化市,江苏 泰州 225724)
20世纪以来,生物学的发展已经逐渐走向成熟,其可预测性也越来越强。然而这种知识的逐渐分化,却使得生物学系统化面临着困难[1]。生态化学计量学在这方面进行了尝试,并使之具备了相当的可能性。生态化学计量主要是指植物的元素组成及其相互关系,特别强调植物主要组成元素C、N、P之间的相互关系[2]。因此,目前国内外生态化学计量学的研究主要集中在C、N、P的计量关系上,包括限制因素判定及生态指示作用等方面[3]。
森林凋落物,一般是指植物在生长发育的过程中所产生的新陈代谢产物[4-5]。凋落叶是森林凋落物的主要组成部分。森林凋落物是森林生态系统养分循环的一个重要过程。德国Ebermager早在19世纪60年代,便对森林凋落物在养分循环中的作用进行了科学研究,而后,国际林学界又深入的进行了一系列的研究[6]。
目前我国已成为世界上第三大氮沉降集中区[7],仅次于欧洲、美国,并且氮沉降问题将越来越严重[8]。氮沉降的增加将不可避免地对森林生态系统产生影响[9]。大气N沉降进入森林地表,会引起森林凋落物N含量及其它化学元素含量的变化。大气N沉降数量的迅速增加已经严重影响到陆地和海洋生态系统元素的供应状况[10]。
中国是世界上竹类资源最丰富的国家,其中毛竹以其生长快、产量高、用途广的特点,成为我国重要的森林资源。高强度的自然氮沉降必然会加重毛竹林的氮输入量。在各生长阶段,植物叶片的N含量、P含量具有较大的变异性[11-12],那么两者的比值(N:P)变化剧烈吗?因此,弄清楚植物生态化学计量随时间而变化的规律就显得十分重要。现有的大范围植物叶片C、N、P含量及其计量学特征的研究结果,大多是基于不同年份的数据集合,其叶片采集的时间能相差1~3个月(如样品采集时间为7-9月)[13-16],那么,不同生长季节植物叶片C、N、P化学计量比是否变化且变化范围又有多大?
为此,本试验以粗放经营毛竹林为对象,通过施氮的方式模拟N沉降增加情景,研究不同水平的氮沉降处理对毛竹凋落叶生态化学计量特征的影响和毛竹凋落叶化学计量特征在1年中的动态变化。
1 材料与方法
1.1 试验地概况
试验地位于浙江省杭州市临安市青山镇(119°42′E,30°14′N)。该区气候温和,年均无霜期 230 d左右,土壤为黄土壤,地形地貌为低山丘陵。研究区内的毛竹林为粗放经营类型,经营措施主要为保留林下灌木、杂草,不进行施肥和林地垦复,出笋时采笋,林中灌木种类主要有檵木(Loropetalum chinensis)、青冈栎(Cyclobalanopsis glauca)、乌饭(Vaccinium bracteatum)和木荷 (Schima superba)和杨桐(Adinandra millettii)等。本研究采用典型选样方法,在研究区设立林分条件和环境状况较一致的代表性样方12个。每个样方面积为20 m×20 m,样方间相隔20 m以免相互影响。样地的林分和土壤特征见表1。

表1 试验毛竹林林分结构和土壤特征Table.1 Stand and soil characteristics of study sites in extensively managed Moso bamboo forest
1.2 氮沉降模拟
依据我国亚热带地区的实际氮沉降量及未来增加趋势[17–18],氮沉降处理设置4个梯度水平:低氮(L,30 kg·N·ha-1·yr-1)、中氮(M,60 kg·N·ha-1·yr-1)、高氮(H,90 kg·N·ha-1·yr-1)和对照(CK,0 kg·N·ha-1·yr-1),每个处理设3个样方即3个重复。每个月进行模拟氮沉降喷施一次,每年12次。具体方法为:每月月初将每个样方所需喷施的一定量的NH4NO3溶解在10升自来水中(相当于年增加降水0.3 mm)。对照处理样方则喷洒同样多的水但不加任何氮。
1.3 毛竹凋落叶的收集,处理与测定
2012年12月底在每个毛竹林样方内随机布设6个1m×0.5m的凋落叶收集框,收集框距地表1m高。于2013年3月开始每月底收集一次凋落叶。把凋落叶带回实验室后放在65℃的烘箱里烘干至恒量。再把凋落叶粉碎,分析C、N和P含量。
1.4 C、N和P元素含量测定方法
C含量用重铬酸钾容量法-外加热 (油浴加热)法测定,N含量用H2SO4-H2O2消煮后,半微量凯氏法测定,P含量用H2SO4-H2O2消煮后,钼锑抗比色法测定[19]。C、N、P含量用质量百分比表示。
1.5 数据处理
比较不同氮沉降强度样地内和不同月份的凋落叶化学计量学特征的差异,统计分析用SPSS 13.0实现,用氮沉降强度水平和凋落叶收集时间作为影响因子进行双因素方差分析。
2 结果与分析
2.1 毛竹凋落叶C、N、P含量的月份变化及其对氮沉降的响应
从图1a可见,凋落叶中的C含量在1年中随着季节的变化没有较明显的变化,总体的趋势较平稳,变幅较小。对各月凋落叶C含量的进一步分析显示,在2013年3月份,只有高氮处理显著增加了C含量(P<0.05),其他处理与对照间差异均不显著;4月份,低氮处理显著增加了C含量(P<0.05),但在超过低氮后则下降;5月份,各处理之间的差异均不显著;6月份和7月份,低氮和中氮处理均显著增加了C含量(P<0.05),但在氮沉降超过中氮后则下降;8月份,C含量在氮沉降超过中氮后下降,低氮和中氮处理与对照间差异不显著;9月份各处理之间的差异均不显著;10月份,高氮处理显著增加了C含量(P<0.05),低氮和中氮处理与对照间差异不显著;11月份,各处理之间的差异均不显著;12月份,高氮处理显著降低了C含量(P<0.05),低氮和中氮处理与对照间没有显著性差异;2014年1月份,低氮处理显著增加了C含量(P<0.05),但在氮沉降强度超过低氮后则下降;2月份,低氮和中氮处理均显著增加了C含量(P<0.05),但在氮沉降强度超过低氮后增加的幅度则开始逐渐变小。1年中,不同水平的氮沉降处理对毛竹凋落叶C含量的影响没有表现出明显的规律性。
植物叶片的元素特征与其自身的结构特征及生长节律间存在着相关关系。毛竹在不同月份的生理及代谢特征不同,尤其是光合作用,呼吸作用及同化物运输和分配的不同,导致毛竹体内C、N和P含量的变化。一般认为,植物细胞内贮藏性物质的比例会随着植物物候期的推迟而逐渐增多,因此,植物叶片C、N、P含量可能会随着植物物候期的不同表现出很大的差异[15]。本试验中,不同月份毛竹凋落叶C含量总体上差异不大。生态系统中,C是组成植物体的结构性物质,N和P则为生物体的功能性物质,其分布和贮量直接关系到生态系统功能的正常发挥。对于同一植物种而言,功能性和贮藏性物质的含量受环境影响变化较大,结构性物质受环境的影响较小,含量相对较稳定[11],这可能是不同月份毛竹凋落叶C含量总体上差异不大的原因。适度的氮沉降能增强毛竹的C同化能力,但在氮沉降超过一定强度后则下降。

图1 试验毛竹凋落叶C、N和P含量Figure 1 The contents of C,N and P of leaves of Moso bamboo forest under different N deposition level
从图1b可见,凋落叶中的N含量随着生长季节变化而发生改变,总体上呈现出先下降后增加的趋势,N含量在夏季较低,其中在7月份是最低的。在2013年3月份和4月份,高氮处理显著降低了凋落叶N含量(P<0.05),低氮和中氮处理与对照间差异不显著;5月份,低氮处理显著增加了N含量(P<0.05),但在氮沉降强度超过低氮后则下降;6月份,低氮处理与对照没有显著性差异,中氮处理显著增加了N含量(P<0.05),但N含量在氮沉降强度超过中氮后下降 ;7、8、9、11、12月份和 2014年 1月份,N含量总体上表现出随氮沉降强度增加而增加的趋势;10月份,高氮处理显著增加了凋落物N含量(P<0.05),低氮和中氮处理与对照间差异不显著;2014年2月份,低氮、中氮和高氮处理均显著增加了凋落物N含量(P<0.05),但在氮沉降超过中氮后增加的幅度则开始变小。1年中,不同水平的氮沉降处理对毛竹凋落叶N含量的影响没有表现出明显的规律性。

图2 试验毛竹凋落叶C:N、C:P和N:PFigure2 The characteristics of C︰N,C︰P and N︰P of leaves of Moso bamboo forest under different N deposition level
植物体内的N含量因季节变动很大,生长初期N素浓度最高,随生长季的延长,生物产量的增加而被稀释,直到生长季结束,植物体内N素浓度降到最低值,相对于C积累而言,叶片积累在初期速度较快,而后期较慢,且往往因淋溶而积累减少[20]。本试验中,从1月开始,由于气温下降,土壤温度也随之降低,毛竹地上部分生长缓慢甚至停止,导致毛竹凋落叶N含量上升;到3月,随着气温的回升,土温也跟着上升,土壤开始解冻,毛竹地上部分生长加剧,同时3-6月又正值毛竹出笋成竹季节,对养分需求特别是N的需求量增加,毛竹凋落叶N含量开始下降;毛竹凋落叶N含量在夏季较低,原因可能是在夏季高温湿润条件下,毛竹凋落叶的分解和养分离子释放速度都会加快,此外,这季节的毛竹叶子都是新叶,伴随着水分的输送,营养元素向幼嫩组织迁移,毛竹体内的N含量较高,其中凋落叶N含量在7月是最低的,即返回到林地土壤的N含量在1年中最小,这与5-8月的毛竹林地土壤养分耗竭最严重的研究结论是一致的[21],此时的毛竹需要补充一定量的肥料。因此,该区域内,未来氮沉降的适量增强仍有利于粗放经营毛竹林的生长,国内外大量的研究也表明:适度的氮沉降,能使森林生态系统的生产力有所提高[22]。Aber[23]等认为,在N输入持续增加直到饱和之前,植物体会吸收大部分N;7月后,急剧地上升;9月后开始有小幅的回落;10月-12月份维持相对稳定的状态。
从图1c可见,凋落叶中的P含量也随着生长季节变化而发生改变,总体上呈现出逐渐下降的趋势,在 12、1、2、3、4、5 月份较高,费世民[24]等也发现凋落物中的P含量呈下降的趋势,和本研究结果基本一致。在2013年3、5月份,低氮处理显著增加了凋落叶P含量(P<0.05),但在氮沉降强度超过低氮后则下降;4、8、9和 10月份,P含量总体上表现出随氮沉降强度增加而增加的趋势,表明适量的氮沉降会提高毛竹对P的利用效率;6月份,低氮处理与对照没有显著性差异,但在氮沉降强度超过低氮后则下降;7月份,P含量总体上呈现出随氮沉降强度增加而减少的趋势;11月份,低氮、中氮处理和高氮处理均显著增加了P含量,但在氮沉降超过中氮后增加的幅度则开始变小,11月份,低氮、中氮处理和高氮处理均显著增加了P含量,但在氮沉降超过中氮后增加的幅度则开始变小。1年中,不同水平的氮沉降处理对毛竹凋落叶P含量的影响亦没有表现出明显的规律性。
一般来说,不同元素的内稳性是不同的。N元素的内稳性高于P,这主要是因为在植物体中,N元素的含量远远大于P元素。关于浮游动物的研究表明浮游动物的大量元素的内稳性高于微量元素,且微量元素高于非必要元素[25],本试验中,凋落叶中的N含量也远远大于P含量。此外费世民[24]等研究表明,毛竹凋落叶中N素的生物自肥潜力高于P元素,因此,毛竹林土壤施肥制度要特别注意保持生态系统养分平衡和土壤肥力的提高。
2.2 毛竹凋落叶C:N、C:P和N:P的月份变化及其对氮沉降的响应
从图2(a)可见,凋落叶的C:N比值在7月份最高,其他月份的比值变幅较小,接近一个常数。在2013年3月和4月份,C:N比值总体上随氮沉降强度增加而增加,在高氮处理下最高;5、6月份,C:N比值在氮沉降强度超过低氮后则开始降低,在低氮处理下最高;7月份,C:N比值亦随氮沉降强度增加而增加;8月份至14年2月份,C:N比值总体上表现出随氮沉降强度增加而降低的趋势。适量的氮沉降可以提高毛竹对N的利用效率,此外,生态系统组分的C:N增加是生态系统碳储存量增加的重要途径。
Mcgroddy[26]等发现尽管生物群系的叶C:N比存在较大差异,但是全球凋落物C:N比接近一个常数,表明了植物的养分再吸收过程对氮磷关系有着广泛的影响。本试验中,除了7月份,其他月份凋落叶C:N比值相对稳定,和McGroddy等的研究结果一致。其中7月份的凋落叶C:N比值较高,表明7月份毛竹对N的利用效率较高,原因可能是因为7月份,毛竹叶都是新叶,处于生长初期。
从图 2(b)可见,从 2013年 3月至 11月份,凋落叶的C:P比值总体上表现出逐渐增加的趋势,11月份出现一个峰值,然后急剧下降。在2013年3月、4月份,C:P比值总体上亦表现出随氮沉降强度增加而增加的趋势;5、6、7月份,C:P比值亦呈现出随氮沉降强度增加而增加的趋势,但在氮沉降强度超过中氮后则下降;8月份,C:P比值在氮沉降强度超过低氮后则开始下降;9月份至2014年2月份,C:P比值总体上表现出随氮沉降强度增加而降低的趋势。
植物叶片C:N和C:P可表示植物吸收营养所能同化C的能力,在一定程度上反映了植物的营养利用效率。凋落叶的C:P比值总体上表现出逐渐增加的趋势,11月份出现一个峰值,然后急剧下降。表明毛竹对P的利用效率逐渐增加,11月份利用效率最高。适量的氮沉降会提高毛竹对N、P的利用效率。
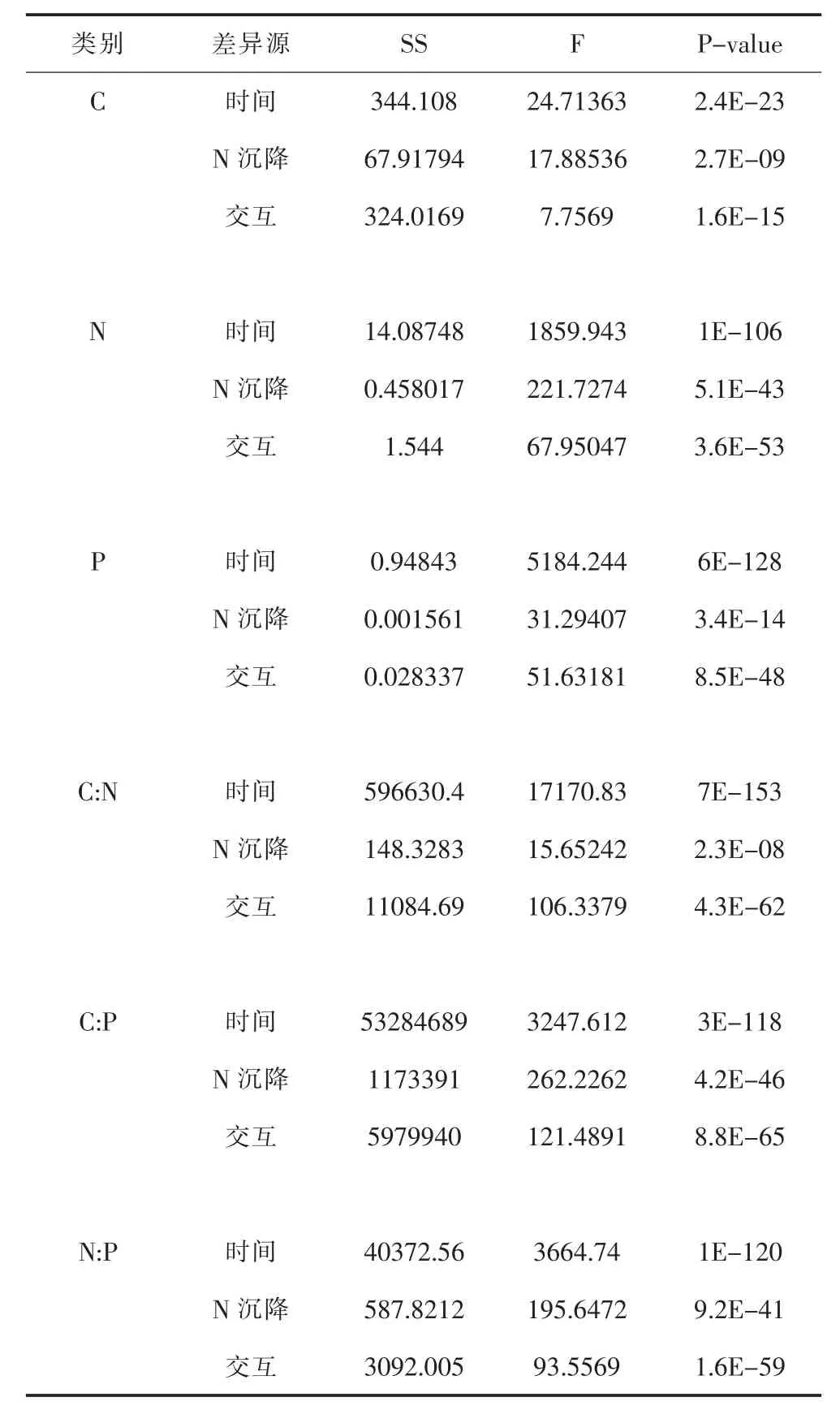
表2 氮沉降和时间对毛竹凋落叶生态化学计量特征影响的双因素方差分析Table 2 The two-way ANOVA of the effects of N deposition and time on stoichiometry of leaves
从图2(c)可见,凋落叶的N:P比值在7月份最低,在 11 月份最高,1 年中,除了 9、10、11 月份,其他月份的N:P比值均小于20。在2013年3、4、6和8月份,N:P比值在氮沉降强度超过中氮后显著降低;5月份,N:P比值在氮沉降强度超过低氮后显著降低;7月份,N:P比值随氮沉降强度增加而增加;9月至11月份,N:P比值总体上呈现出随氮沉降强度增加而降低的趋势;12月至2014年2月份,N:P比值总体上呈现出随氮沉降强度增加而增加的趋势。
Elser[27]等认为,植物体内氮磷比值会随着植物体的高速率生长而变化。本试验中,分别比较不同氮沉降水平之间、不同月份的氮磷比值的变化,可以看到氮沉降总体上改变了毛竹凋落叶的氮磷比值,毛竹在不同月份的氮磷比值是不同的,这也充分证明了植物元素计量的自我调节机制。杨阔[28]等对青藏高原草地植物的研究发现,群落水平叶片的N、P含量及N:P比存在显著的年际变化。研究发现,植物叶片N:P会随着生长季节变化而发生改变[29],但叶片N:P值随季节变化没有一致规律[30]。叶片N:P值的季节变化是否意味着植物生长所需的元素的限制性也会随着生长期的不同而改变?这种变化在不同生态系统间是否具有普遍性也有待于进一步研究。
如表2所示,双因素方差分析结果表明,氮沉降水平、时间及其与氮沉降的的交互作用对凋落叶C含量、N 含量、P 含量、C:N、C:P 和 N:P 均有显著的影响。
毛竹一次造林后可数十年连续收获、年年萌发新竹新鞭自我更新,生长速度非常惊人,其年固碳能力也非常巨大。因此,当前及未来持续增强的氮沉降对毛竹林生产力进而对其固碳能力的影响应该引起足够的重视。
参考文献:
[1]ELSER J J,STERNER R W,GOROKHOVA E,et al.Biological stoichiom-etry from genes to ecosystems[J].Ecology Letters,2000,(3):540-550.
[2]ELSER J J,DOBBERFUHL D R,MACKAY N A,et al.Organism size,life history,and N:P stoichiometry[J].BioScience,1996:674-684.
[3]ZHANG L X,BAI Y F,HAN X G.Application of N:P stoichiometry to ecology studies [J].Acta Botanica Sinica,2003,45(3):1009-1018.
[4]王凤友.1989.森林凋落量研究综述 [J].生态学进展,6(2):82~89.
[5]林波,刘庆,吴彦,等.森林凋落物研究进展[J].生态学杂志,2004,23(1):60-64.
[6]Wardle D A,Nilsson M C,Zackrisson O,et al.Determinants of litter mixing effects in a Swedish boreal forest [J].Soil Biology and Biochemistry,2003,35(6):827-835.
[7]HOLLAND E A,DENTENE F J R,BRASWELLl B H,et al.Contemporary and pre-industrial global reactive nitrogen budgets [M]//New Perspectives on Nitrogen Cycling in the Temperate and Tropical Americas,Springer Netherlands,1999,46:7-43.
[8]MO J M,BROWN S,XUE J H,et al.Response of litter decomposition to simulated N deposition in disturbed,rehabilitated and mature forests in subtropical China [J].Plant and Soil,2006,282(1-2):135–151.
[9]BOXMAN A W,BLANCK K,BRANDRUD T E.Vegetation and soil biota response to experimentallychanged nitrogen inputs in coniferous forest ecosystems of the NITREX project [J].Forest Ecology and Management,1998,101(1):65-79.
[10]王绍强,于贵瑞.生态系统碳氮磷元素的生态化学计量学特征[J].生态学报,2008,28(8).
[11]Sterner R W,Elser J J.Ecological stoichiometry:the biology of elements from molecules to the biosphere[M].Princeton University Press,2002.
[12]HAN W X,FANG J Y,GUO D L,et al.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J].New Phytologist,2005,168:377–385.
[13]Reich P B,Oleksyn J.Global patterns of plant leaf N and P in relation to temperature and latitude [J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):11001-11006.
[14]Wright I J,Reich P B,Westoby M,et al.The worldwide leaf economics spectrum [J].Nature,2004,428(6985):821-827.
[15]Kerkhoff A J,Enquist B J,Elser J J,et al.Plant allometry,stoichiometry and the temperature‐dependence of primary productivity[J].Global Ecology and Biogeography,2005,14(6):585-598.
[16]李玉霖,毛伟,赵学勇,等.北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究 [J].环境科学,2010,31(8):1716-1725.
[17]LIU X J,ZHANG Y,HAN W X,et al.Enhanced nitrogen deposition over China [J].Nature,2013,494(7438):459-462.
[18]谢迎新,张淑利,赵旭,等.长江三角洲地区雨水中NH4+-N/NO3-N和δ 15 NH4+ 值的变化 [J].应用生态学报,2008,19(9):2035-2041.
[19]张万儒,杨光谨,屠星南,等.中华人民共和国林业行业标准-森林土壤分析方法[M].北京:中国标准出版社,1999.
[20]吴统贵,吴明,刘丽,等.杭州湾滨海湿地 3种草本植物叶片 N,P化学计量学的季节变化 [J].植物生态学报,2010,34(1):23-28.
[21]练佑明,周灵甫.黄脊竹蝗防治指标研究[J].林业科学研究,1992,5(6):717-721.
[22]MAGILL A H,ABER J D,BEMTSON G M,et al.Long-term nitrogen additions and nitrogen saturation in two temperate forests [J].Ecosystems,2000,3:238-253.
[23]ABER J D,NADELHOFFER K J,STEUDLER P,et al.Nitrogen saturation in northern forest ecosystems[J].BioScience,1989,39(6):286-378.
[24]费世民,张旭东.竹林生态研究[J].2011.
[25]Karimi R,Folt C L.Beyond macronutrients:element variability and multielement stoichiometry in freshwater invertebrates[J].Ecology letters,2006,9(12):1273-1283.[26]McGroddy M E,Daufresne T,Hedin L O.Scaling of C:N:P stoichiometry in forests worldwide:Implications of terrestrial redfield‐type ratios[J].Ecology,2004,85(9):2390-2401.
[27]Elser J J,Acharya K,Kyle M,et al.Growth rate–stoichiometry couplings in diverse biota [J].Ecology Letters,2003,6(10):936-943.
[28]杨阔,黄建辉,董丹,等.青藏高原草地植物群落冠层叶片氮磷化学计量学分析 [J].植物生态学报,2010,34(1):17-22.
[29]Santa Regina I,Tarazona T,Calvo R.Aboveground biomass in a beech forest and a Scots pine plantation in the Sierra de la Demanda area of northern Spain[C]//Annales des Sciences forestières.EDP Sciences,1997,54(3):261-269.
[30]gren G I.Stoichiometry and nutrition of plant growth in natural communities [J].Annual review of ecology,evolution,and systematics,2008:153-170.
猜你喜欢
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
河北遥感(2017年2期)2017-08-07
环境科技(2016年1期)2016-11-08
中国医学影像学杂志(2015年9期)2015-12-15
中国铸造装备与技术(2015年5期)2015-12-10
西藏科技(2015年4期)2015-09-26
电子设计工程(2014年18期)2014-02-27
植物营养与肥料学报(2012年5期)2012-10-26
