
向日葵分枝和株高性状的关联分析定位研究
2017-05-15 08:23:33刘胜利柳延涛王沛政
海南热带海洋学院学报 2017年2期
刘胜利,段 维,王 鹏,柳延涛,王沛政
(1.新疆农垦科学院,新疆 石河子 832000; 2.海南热带海洋学院 生命科学与生态学院,海南 三亚 572022;3.新疆康地种业科技股份有限公司,乌鲁木齐 830011)
向日葵分枝和株高性状的关联分析定位研究
刘胜利1,段 维3,王 鹏1,柳延涛1,王沛政2
(1.新疆农垦科学院,新疆 石河子 832000; 2.海南热带海洋学院 生命科学与生态学院,海南 三亚 572022;3.新疆康地种业科技股份有限公司,乌鲁木齐 830011)
经过structure软件分析,128份向日葵自交系材料基因型依据△K的变化,可分为2大类群.标记间的连锁不平衡分析显示,SSR标记间的r2值范围在0-1之间,平均为0.02;在选用D′来计算标记位点间的连锁不平衡时, 25%的成对位点组合标记中存在较高水平的连锁不平衡(D′>0.4).关联分析定位到 8个标记位点与向日葵分枝性状相关联,其中有6个位点p<0.001,分别是ORS679、ORS 227-1、ORS16、 ORS22、 ORS33和ORS38;解释表型变异8%-16%不等,2个标记位点p<0.01,分别是标记ORS803和ORS 227-2;在株高性状中,定位到3个位点与其显著关联(p<0.01),分别是ORS679,ORS9、ORS22,解释表型变异4%-6%不等.该研究成果将会为今后利用分子标记技术在向日葵新品种选育方法上提供支持.
向日葵;群体结构; 关联分析
0 引言
向日葵(Helianthus annuus L.)是特别适合于我国西北部干旱地区种植,具有耐盐碱、耐瘠薄、抗旱、适应性强的农作物[1].油用向日葵籽实含油率较高,亚油酸含量在60%以上,目前为世界第4大油料作物[2].当前国内向日葵品种与国外品种综合农艺性状相比还有一定差距,主要原因是当前我国向日葵品种科研育种基础研究还比较薄弱[3].我国向日葵育种主要采用常规育种,向日葵常规育种方法存在着盲目性大、周期长、效率低的问题,尤其当对多基因控制的数量性状进行育种时,容易导致偏离预期的育种目标.近几年,分子标记技术已发展成为提高向日葵育种工作效率的强有力武器,成为当前向日葵生物学研究的热点.
向日葵的分枝农艺性状是在向日葵生长初期,由向日葵叶腋处的腋芽萌发,形成分枝.向日葵多头分枝性状主要由单显性基因控制、单隐性基因控制和双隐性基因控制,在不同材料中,多头分枝性状可能受到不同基因的调控[4].向日葵株高主要是基因加性遗传效应控制,也受非加性效应影响,属于较复杂的数量性状遗传,受多基因调控外,环境条件一定程度影响遗传表达[5-7].
在向日葵多头和分枝定位研究方面,文献[8]报道了利用RFLP标记,在F2群体中将向日葵多头基因b1定位在LG 7上.文献[9]利用了重组自交系群体将向日葵多头基因定位于两个SSR标记(ORS1088, ORS930)之间.文献[10]则利用F2群体将多头基因定位在标记TBR11-107和TBR4-720/TBRB-555 之间.文献[11]指出了向日葵矮化突变基因由半显性等位基因Rht1所控制,并将其定位到连锁群12上.
综上所述,以上向日葵分枝和株高性状的基因定位研究均使用单一亲本的杂交分离群体,研究所用标记也不尽相同,缺乏对向日葵整个群体资源材料进行数量性状的筛选定位研究,因此研究所得出的结果有较大的局限性.由于不同研究者使用的分子标记不同,所得的研究结论在不同实验室间也较难进行比较,限制了其在向日葵育种中的有效应用.利用关联分析方法进行数量性状基因定位研究周期短、分辨率高,可以同时对多个基因进行筛选,确定不同种质资源中所具有的等位基因及其对目标的贡献,是目前研究复杂数量性状基因变异的常用方法.影响向日葵分枝和株高性状的等位基因变异在其基因组中广泛分布,因此利用关联分析方法可以有效地鉴定出这些功能等位基因的位点.本研究利用向日葵自交系为材料,利用随机筛选的向日葵分子标记,分析向日葵资源材料自交系的遗传结构.在此向日葵遗传结构的基础上,对向日葵多头分枝和株高性状进行关联分析定位.该研究结果将为今后向日葵亲本选育提供可靠的分子标记,加速向日葵自交系的培育,节约育种时间.同时也可以为今后该基因的图位克隆打下良好的基础.
1 材料与方法
1.1 试验材料
向日葵不同自交系种子材料由新疆农垦科学院作物所提供.为了尽可能使研究的向日葵品系具有较高的遗传多样性,我们收集了128份向日葵油葵和食葵各种自交系(见表1),包括油葵保持系30份和恢复系43份;食葵保持系20份和恢复系35份.田间种植试验株距 25 cm,行距 70 cm,栽培密度 1.4×104株/hm2.小区行长 6 m,4 行区,采用完全随机区组设计,3 次重复,生育期田间管理同当地大田生产.开花期调查材料的分枝情况,收获期测量株高.
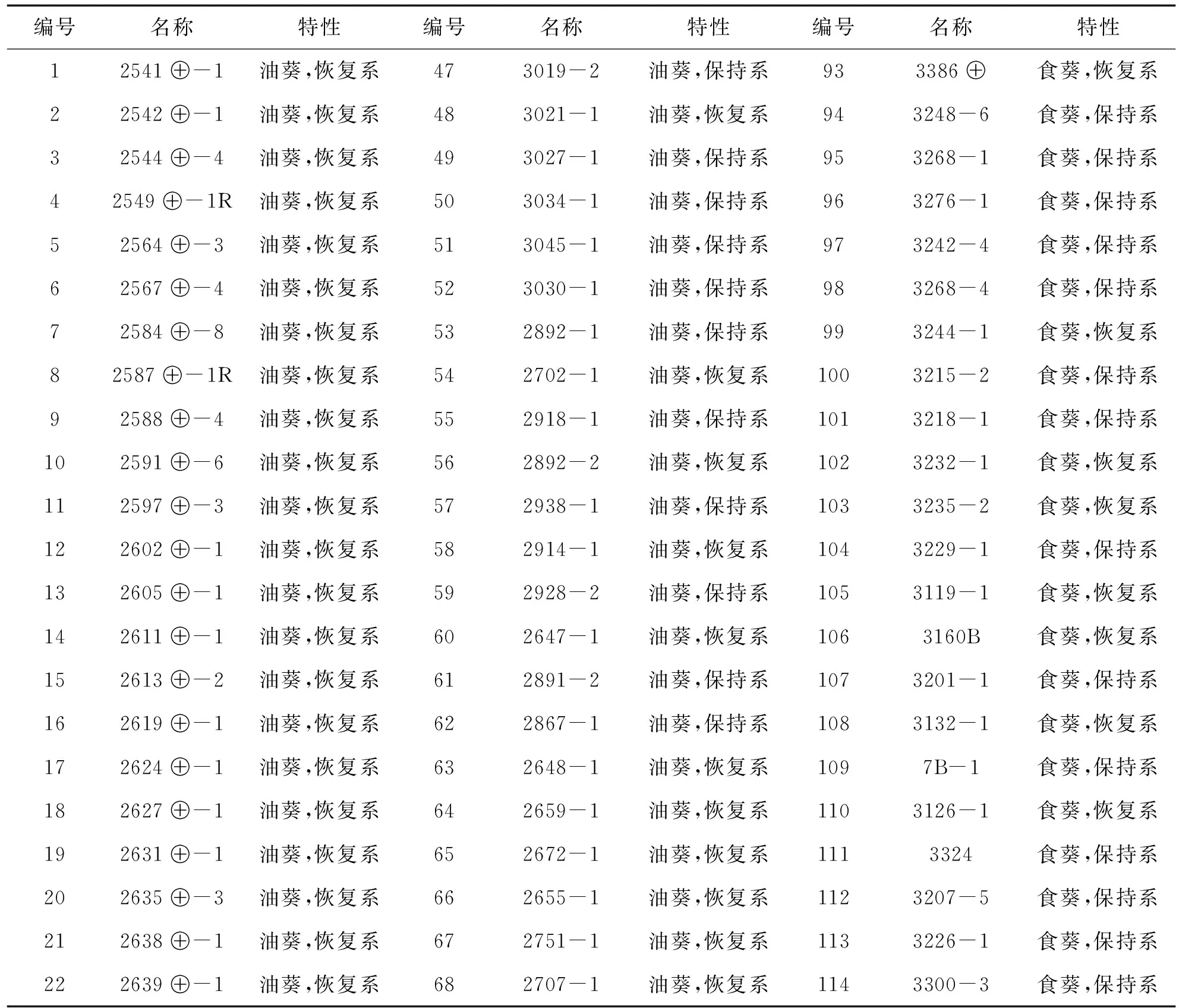
表1 向日葵自交系材料
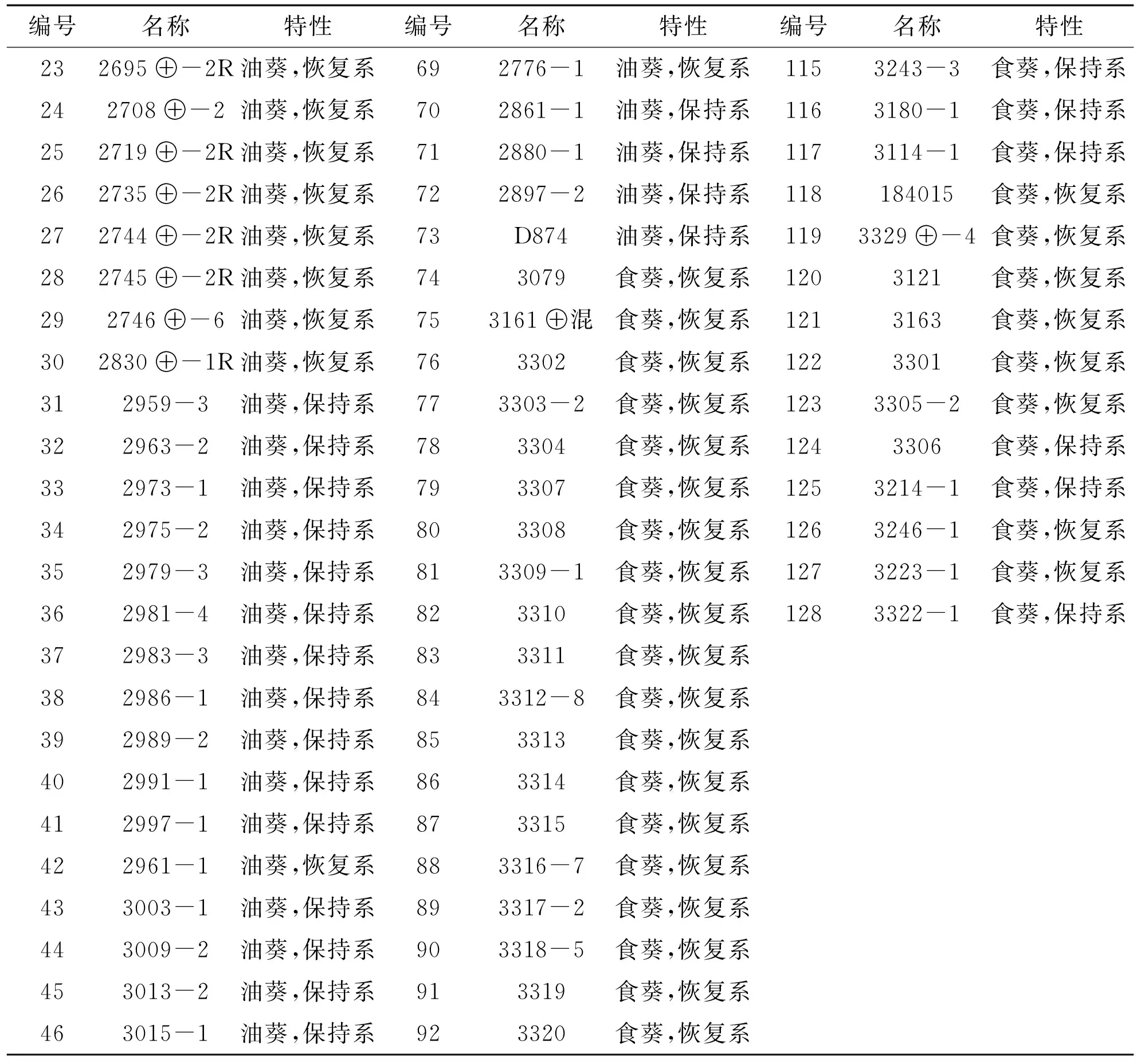
续表1
1.2 DNA提取、PCR扩增和聚丙烯酰胺凝胶电泳
DNA提取:供试材料种植于海南三亚农垦科学院试验田.取新鲜叶片,当天取样后直接提取基因组DNA.DNA提取采用百泰克新型快速植物基因组DNA提取试剂盒(离心柱型),最后得到的DNA样品用TE(Tris-EDTA)缓冲液溶解,放在-20℃的冰箱内保存.
TaqDNA聚合酶、dNTPs、Marker ladder、溴酚蓝、丙烯酰胺、甲叉双丙烯酰胺和TEMED等主要生化试剂购自海南相关生物销售商,为国产分析纯.
SSR引物:SSR引物见文献[12].在该文报道的向日葵序列标签位点(sequence tagged site)序列中,随机选取500对SSR引物,由上海生物工程公司合成引物,首先对16份向日葵自交系材料进行扩增筛选多态.
PCR 扩增、PCR反应程序以及变性聚丙烯酰胺凝胶参照有关文献[13].
1.3 群体结构分析
应用STRUCTRE软件,对供试群体进行基于数学模型的类群划分.先设定群体数目(K)为2-10,5次重复,将MCMC设为100000次,其参数设定参见已发表文献[14].
1.4 关联分析
在预估群体的 K 值范围,获得稳定可靠的群体分类结果.在此基础上以各个体 Q 值作为协变量进行群体矫正,将向日葵分枝和株高性状的表型数据分别对标记变异进行回归分析,评价标记等位变异的平均效应,依据显著水平(p<0.05和p< 0.01) ,选取贡献率高的主要位点[15].
2 结果分析
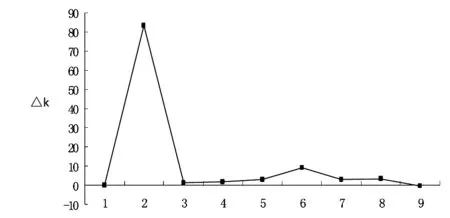
图1 利用△K对向日葵群体结构的优化的双边图表
2.1 向日葵资源材料的群体结构分析
128份向日葵资源材料群体基因型经过structure软件运行后,结果表明供试群体的等位变异频率特征类型数K=2时ΔK值最大(见图1),而且差异显著.在K为3-9之间,其△K基本降低到0附近,近似一条直线.因此依据△K的变化,向日葵自交系材料群体可以分为2大类群.
2.2 向日葵群体标记间的连锁不平衡分析
向日葵群体标记间的连锁不平衡分析显示,对于全部群体SSR标记间的r2值的范围在0-1之间,平均为0.02;而D′值在0-1之间,D′平均值达0.26.在选用D′来计算标记位点间的连锁不平衡时,共有1320个SSR标记成对位点组合中存在连锁不平衡,占所有标记可能成对位点组合的99%.其中25%的标记成对位点组合中存在较高水平的连锁不平衡(D′>0.4),其中统计概率p<0.01的组合有67对,占5%.
2.3 向日葵群体分枝和株高性状关联分析及其等位变异表型效应
向日葵开花期调查统计结果表明,所研究的向日葵群体共有84份材料表现为不分枝,44份材料具有分枝性状.收获期向日株高性状统计表明,株高为0.4米的有8份材料,0.5米的有6份材料,0.6米的有5份材料,0.7米的有2份材料, 0.8米的有26份材料, 0.9米的有6份材料, 1米的有49份材料, 1.1米的有15份材料, 1.2米的有7份材料, 1.3米的有4份材料,所有株高性状呈现极端株高类型数目少,中等株高类型的材料占多数.
利用structure软件中得到的各向日葵个体相应的Q值作为协变量,分别对向日葵分枝和株高性状的表型值和标记变异进行回归性分析,计算其相关联的位点及其等位变异.结果显示:在所检测的利用62对ORS标记位点中,共有9个位点与性状相关(见表2).其中8个位点与向日葵分枝性状相关联,其中有6个位点显著关联(p<0.001),分别是ORS679、ORS 227-1、ORS16、 ORS22、 ORS33和ORS38;解释表型变异8%-16%不等,6个位点显著关联(p<0.01),分别是ORS803、ORS 227-2;在株高性状状中,有3个位点显著关联(p<0.01),分别是ORS679,ORS9、ORS22,解释表型变异4%-6%不等.
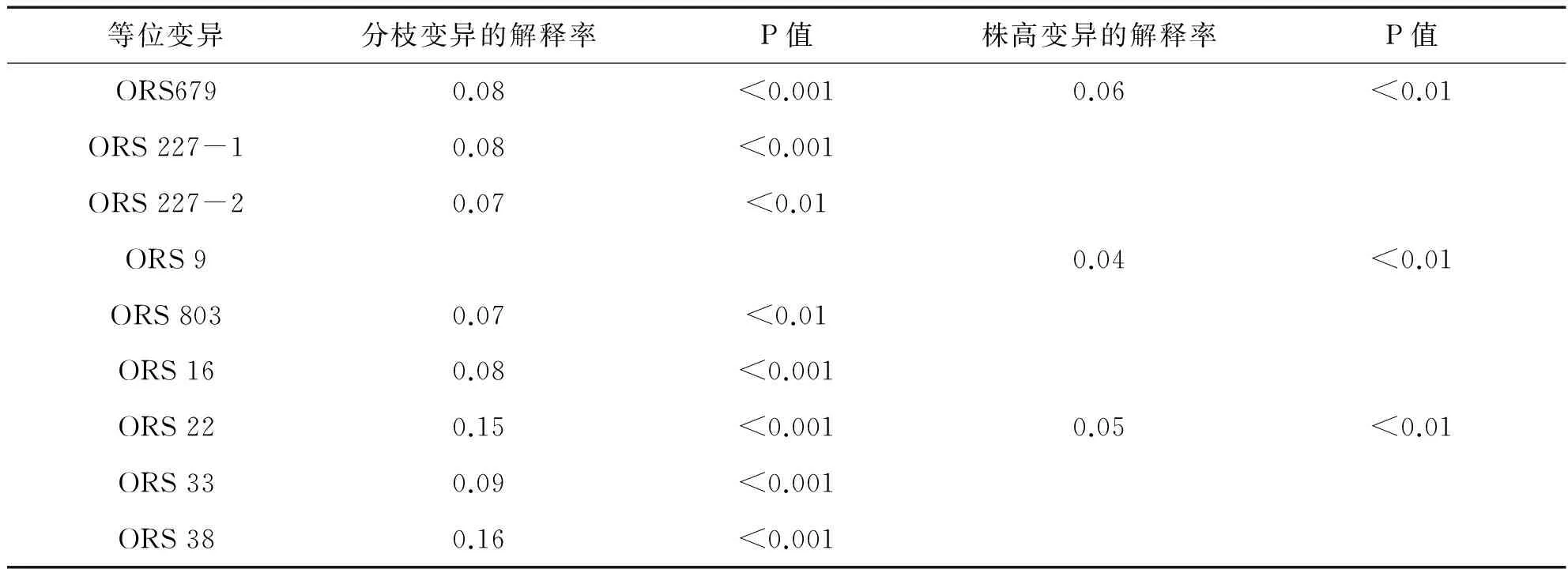
表2 与性状相关联(p<0.01和p<0.001)的标记位点及其表型变异解释率
3 讨论
向日葵多头植株花期较长,在向日葵杂交种制种过程中常作为父本,可以解决单头亲本间花期难于相遇的问题,显著提高向日葵杂交制种的产量.因此在向日葵种质资源研究中向日葵多头分枝性状是很重要的农艺性状.文献[9]将多头性状定位到第10号染色体上的ORS1088和ORS908之间.本研究定位到8个位点与向日葵分枝性状相关联,分别是ORS679、ORS 227-1、ORS16、ORS22、ORS33、ORS38、ORS803、ORS 227-2,解释表型变异8%-16%不等,和已发表的向日葵图谱比对只有ORS679位于17号染色体,ORS803定位于16号染色体,其它标记未能与向日葵已知的连锁群比对上.本研究中的关联分析表明在所研究的向日葵资源材料中,向日葵分枝性状由多数基因位点控制,一些定位到的向日葵分枝位点可能是以前文献所没报道过.向日葵茎干倒伏给向日葵生产造成严重的损失.然而降低植株高度将有助于增强茎干强度,减少倒伏,从而提高产量,因此现在育种的策略就是开发矮杆的种质资源,以利于其在育种中应用.本研究在株高性状状态中,有3个位点显著关联(p<0.01),分别是标记ORS679,ORS9、ORS22,解释表型变异4%-6%不等.本研究检测到的株高关联位点不多,主要原因可能还是株高是由复杂数量性状基因控制,同时所研究的分子标记数量还不够多,因此在今后研究工作当中应继续增加标记数量.
[1]崔良基,刘悦,王德兴.我国发展向日葵生产潜力及对策 [J].杂粮作物,2008,28(5):336-338.
[2]罗伟强.气相色谱法测定葵花籽油的脂肪酸[J].食品工业科技, 2003, 24(6):79-80.
[3]张明.国内外向日葵育种概况及动向[J].黑龙江农业科学, 2010(6):149-151.
[4]Hockett E A, Knowles P F.Inheritance of branching in sunflowers,HelianthusannuusL[J].Crop Sci .1970,10:432-436.
[5]王贵,刘学文,韩英.向日葵不育系恢复系主要性状配合力分析[J].黑龙江农业科技,1989(4):12-16.
[6]Chandra B S, Kumar S S, Ranganadha A R G, Dudhe M Y.Combining ability studies for development of new hybrids over environments in sunflower(HelianthusannuusL.)[J].Journal of Agricultural Science, 2011,3(2):230-237.
[7]Andarkhor S A, Mastibege N, Rameeh V.Combining ability of agronomic traits in sunflower (HelianthusannuusL.) using LinexTester analysis[J].International Journal of Biology,2011,4(1):89-95.
[8]Gentzbittel L, Mestries E, Mouzeyar S, Mazeyrat F, Badaoui S, Vear F, de Labrouhe DT, Nicolas P.A Composite map of expressed sequences and phenotypic traits of the sunflower (HelianthusannuusL.) genome[J].Theoretical and Applied Genetics, 1999,99(99):218-234.
[9]Tang S, Leon A, Bridges WC, Knapp SJ.Quantitative trait loci for genetically correlated seed traits are tightly linked to branching and pericarp pigment loci in sunflower.Crop Science, 2006, 46(2):721-734.
[10]Rojas-Barros P, Hu J, Jan C.Molecular mapping of an apical branching gene of cultivated sunflower (HelianthusannuusL.) [J].Theoretical and Applied Genetics, 2008, 117(1):19-28.
[11]Ramos M L, Altieri E, Bulos M, Sala C A.Phenotypic characterization, genetic mapping and candidate gene analysis of a source conferring reduced plant height in sunflower[J].Theoretical and Applied Genetics,2013, 126(1): 251-263.
[12]Tang S, Yu J K, Slabaugh B, et al.Simple sequence repeat map of the sunflower genome[J].Theoretical and Applied Genetics.2002,105 (8):1124-1136.
[13]夏春兰,卫泽,张雪钰,等.改良测序聚丙烯酰胺凝胶和普通聚丙烯酰胺凝胶技术在玉米分子标记中的应用研究[J].琼州学院学报, 2014, 21(5): 64-68.
[14]张俊,王静毅,陈友,等.基于模型的香蕉种质资源群体结构聚类分析及其亲缘关系分析[J].热带作物学报, 2014,35(2):232-238.
[15]文自翔,赵团结,郑永战,等.中国栽培和野生大豆农艺及品质性状与 SSR 标记的关联分析II优异等位变异的发掘[J].作物学报, 2008,34(8):1339-1349.
(编校:李由明)
Correlation Analysis on the Branches and Plant Height of the Sunflower
LIU Sheng-li1, DUAN Wei1, WANG Peng1,LIU Yan-Tao1,WANG Pei-zheng2
(1.Xinjiang Academy of Agricultural Reclamation Science, Shihezi Xinjiang 832000, China;2.School of Life Science and Ecology, Hainan Tropical Ocean University, Sanya Hainan 572022, China;3.Xinjiang Kangdi Seed Science & Technology Co.Ltd, Urumqi 830011, China)
According to the variety of △K, 128 sunflower inbred lines, analyzed by using the structure software, were divided into 2 groups.The Linkage disequilibrium between the markers shows that r2value is in the range of 0-1 with an average of 0.02.When D′ was applied to calculate the linkage disequilibrium between loci, 25% of the marker paired point combination has a high level of linkage disequilibrium (D′>0.4).Result of the correlation analysis showed that 8 marker loci were found to correlate with branch traits, of which, 6 loci—ORS679, ORS 227-1,ORS16, ORS22, ORS33 and ORS38 with phenotypic variation ranging from 8%-16%—arep<0.001, 2 loci—ORS 803 and 227-2 arep<0.01, 3 loci—ORS679, ORS9, ORS22—are signif-icantly associated with the traits of plant height withp<0.01, with their phenotypic variation ranging from 4%-6%.The collection of the identified candidate loci provides a pool of promising for the future application of molecular marker technology in the sunflower breeding.
sunflower; population structure; correlation analysis
格式:刘胜利,段维,王鹏,等.向日葵分枝和株高性状的关联分析定位研究[J].海南热带海洋学院学报,2017,24(2):80-84+95.
2016-09-15
新疆兵团现代农业科技攻关与成果转化项目(2016AC024,2016AC027);三亚市院地科技合作项目(2013YD41)
刘胜利(1967-),男,山东宁津人, 新疆农垦科学院研究员,研究方向为向日葵新品种选育与栽培推广.
王沛政(1972-),男,陕西长安人,海南热带海洋学院热带生物与农学院副教授,博士,研究方向为分子遗传育种.
S565.5
A
2096-3122(2017) 02-0080-05
10.13307/j.issn.2096-3122.2017.02.16
猜你喜欢
语文周报·教研版(2021年28期)2021-08-19 02:14:42
小天使·一年级语数英综合(2020年8期)2020-12-16 02:56:49
农村百事通(2019年17期)2019-10-08 02:24:55
数学理论与应用(2017年2期)2017-06-27 07:38:56
数学理论与应用(2017年2期)2017-06-27 07:38:54
现代农业科技(2017年1期)2017-03-06 23:08:18
江苏农业科学(2016年8期)2017-02-15 19:19:29
江苏农业科学(2016年8期)2017-02-15 19:19:15
文理导航·科普童话(2016年5期)2016-07-14 15:38:41
数学理论与应用(2016年1期)2016-02-28 09:25:55
