
流沙湾海域皮氏叫姑鱼生物学特性的初步研究
2017-05-15 08:22申玉春操玉涛
海南热带海洋学院学报 2017年2期
郭 慧,徐 明,申玉春,叶 宁,操玉涛
(广东海洋大学 水产学院,湛江市海洋生态与养殖环境重点实验室,广东 湛江 524025)
流沙湾海域皮氏叫姑鱼生物学特性的初步研究
郭 慧,徐 明,申玉春,叶 宁,操玉涛
(广东海洋大学 水产学院,湛江市海洋生态与养殖环境重点实验室,广东 湛江 524025)
在2013年1月-12月渔业资源调查数据的基础上,研究了流沙湾海域皮氏叫姑鱼的体长分布、体长(L)-体质量(W)的关系、肥满度、饵料组成、摄食强度和营养级等生物学特性.结果表明:流沙湾海域皮氏叫姑鱼体长80-191 mm,平均139 mm,其中优势体长组为121-140 mm;体长-体质量关系方程为:W=0.0134L3.1718(R2=0.9734);肥满度最大值出现在2月份,最小值出现在4月份;皮氏叫姑鱼的饵料组成以甲壳类为主要类群.皮氏叫姑鱼的空胃率随季节变化显著(P<0.05),在冬季最大,春季达到最小;饱满度指数在秋季最高,冬季最低.充塞度等级显示,在春季,主要以0级和1级为主;在秋季,充塞度在1级以上的比例最高;结果显示营养级以2、3级为主,平均为3.06,随着体长的增加,营养级也增加.
皮氏叫姑鱼;生物学特性;饵料组成;流沙湾
0 引言
皮氏叫姑鱼(Johniusbelangerii)是分布于印度-西太平洋热带海域的暖水性中下层鱼类,隶属鲈形目(Perciformes),石首鱼科(Sciaenidae),叫姑鱼属(Juhnius) ,在我国沿海、日本和韩国等地区均有分布(http://fishdb.sinica.edu.tw/chi/species.php?id=382469).是许多海域鱼类群落的优势种[1-3].皮氏叫姑鱼是一种肉食性鱼类,以底栖的虾类、贝类和小型鱼类为食[4],同时又是大黄鱼、带鱼、牙鲆等鱼类的生物饵料.在捕捞渔业上,皮氏叫姑鱼是主捕的经济种类之一,是海洋食物网的一个重要环节,在海洋生态系统中也占据了重要的地位.然而,随着人们对海洋资源的过度索取,尤其是近海的过度捕捞,导致了生物资源种类与数量上的迅速减少[5],再加上近年来人为活动的影响导致水环境恶化,都使皮氏叫姑鱼资源已开始衰退[6].
国内外关于皮氏叫姑鱼的研究,主要集中在体内重金属含量[4]、系统进化分析[7]、遗传多样性分析[8]等方面,对皮氏叫姑鱼基础生物学特性的研究报道有:国外学者研究了阿拉伯海域皮氏叫姑鱼体长体质量的关系与氟化物浓度的相关性[9];王家樵等[10]初步研究了福建沿岸海域皮氏叫姑鱼的渔业生物学特征和种群参数;王凯等[6]对马鞍列岛海域皮氏叫姑鱼的的体长组成、体长与体质量关系、性比、繁殖习性和摄食等渔业生物学进行了初步研究.流沙湾是典型的亚热带海湾,位于广东省雷州半岛西南部,是北部湾支流,海水水质环境较好,蕴藏着丰富的海洋鱼类资源.课题组前期对流沙湾渔业资源的调查表明,在流沙湾海域的鱼类渔获物中,皮氏叫姑鱼在数量和重量上都具有优势.目前,对流沙湾海域皮氏叫姑鱼的生物学研究还未见报道.本文利用流沙湾海域鱼类资源调查时收集的皮氏叫姑鱼实验材料,分析了该鱼种的体长分布、体长-体质量关系、肥满度月变化、饵料组成、摄食强度和营养级等生物学特性,为流沙湾海域皮氏叫姑鱼资源的开发利用和保护奠定理论基础.
1 材料与方法
1.1 样品采集
课题组从2013年1月至12月期间,每月下旬到湛江市流沙湾覃斗镇鱼港码头收集皮氏叫姑鱼样品,共采集鱼类样品541尾.
1.2 实验室分析
样品采集当天在实验室完成皮氏叫姑鱼的生物学测量分析.全长和体长的测定用游标卡尺进行,体质量的测定用电子天平(精度0.001 g)进行.
测量相关生物学数据后,解剖,取出胃和消化道内的食物,用吸水纸吸去饵料生物上的水分后,根据残体确定每种饵料生物的个数,然后用电子天平(精度0.001 g)对每种饵料生物进行称重.
稳定同位素碳、氮比值的测定使用Delta V Advantgae质谱仪(Thermo Scientific, USA)进行,每组样品测3个重复,碳、氮稳定同位素比值精密度为±0.2‰.
1.3 数据处理
统计结果以平均值±标准差来表示(mean±SD),所有数据采用Microsoft Excel 2003 和SPSS18.0软件进行分析[11].用幂函数回归法拟合体长与体质量的关系.并计算肥满度,计算公式为CF=W/L3×100%,其中:CF为肥满度(%),W为体重(g),L为体长(mm).用来描述鱼类饵料组成的指标主要有个数百分比(N%)、质量百分比(W%)和出现频率(F%),这些指标可以用一个综合性指数来表示,即相对重要性指数(IRI),本文用相对重要性指数来评估皮氏叫姑鱼不同饵料生物的重要性[6-12],计算公式为:
IRI=(W%+N%)×F%×104.
研究摄食强度的变化情况时,采用饱满指数、空胃率和充塞度三个指标,所用到的计算公式如下:
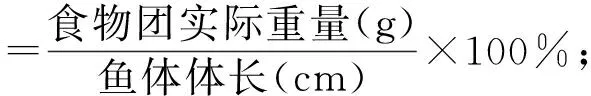
2 结果
2.1 体长分布
本研究中,共测量皮氏叫姑鱼541尾,全长为128-223 mm,平均164 mm,体长80-191 mm,平均139 mm.总体上,皮氏叫姑鱼体长分布呈现6个体长范围,其中体长在121-140 mm之间的数目最多,141-160 mm的次之(图1).
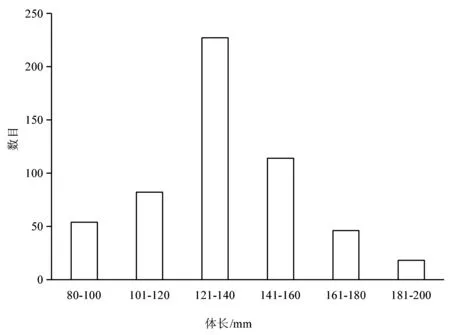
图1 皮氏叫姑鱼的体长分布(n=541)
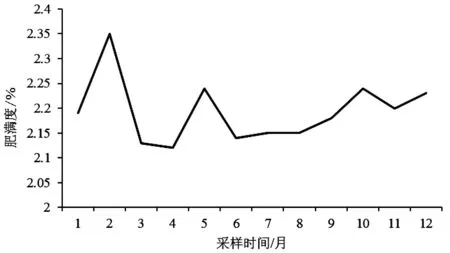
图2 皮氏叫姑鱼的肥满度月变化
2.2 体长-体质量关系
数据分析表明,皮氏叫姑鱼体长和体重有密切的相关性,根据回归分析,其体长-体重的关系方程为:W=0.0134L3.1718,R2=0.9734.生长指数为b值>3,说明皮氏叫姑鱼的生长特性呈正异速生长的特征[12].
2.3 肥满度月变化
衡量鱼类营养状况的指标是鱼体的肥满程度.如图2所示,皮氏叫姑鱼的肥满度在2月份和4月份达到最高和最低值,分别为2.35%和2.12%.6月份开始肥满度呈现逐渐上升的趋势.
2.4 饵料组成
皮氏叫姑鱼的摄食饵料中出现频率(F%)最高的是甲壳类,为81.22%,重量百分比(W%)和个数百分比(N%)均为最高.甲壳类中又以虾类的F%为最高,为33.01%,其次是磷虾类28.75%,蟹类8.77%.鱼类的F%为13.28%,W%为28.95%.多毛类F%为5.63%,W%为3.17%,头足类的F%为0.71%,W%为0.45%(表1).
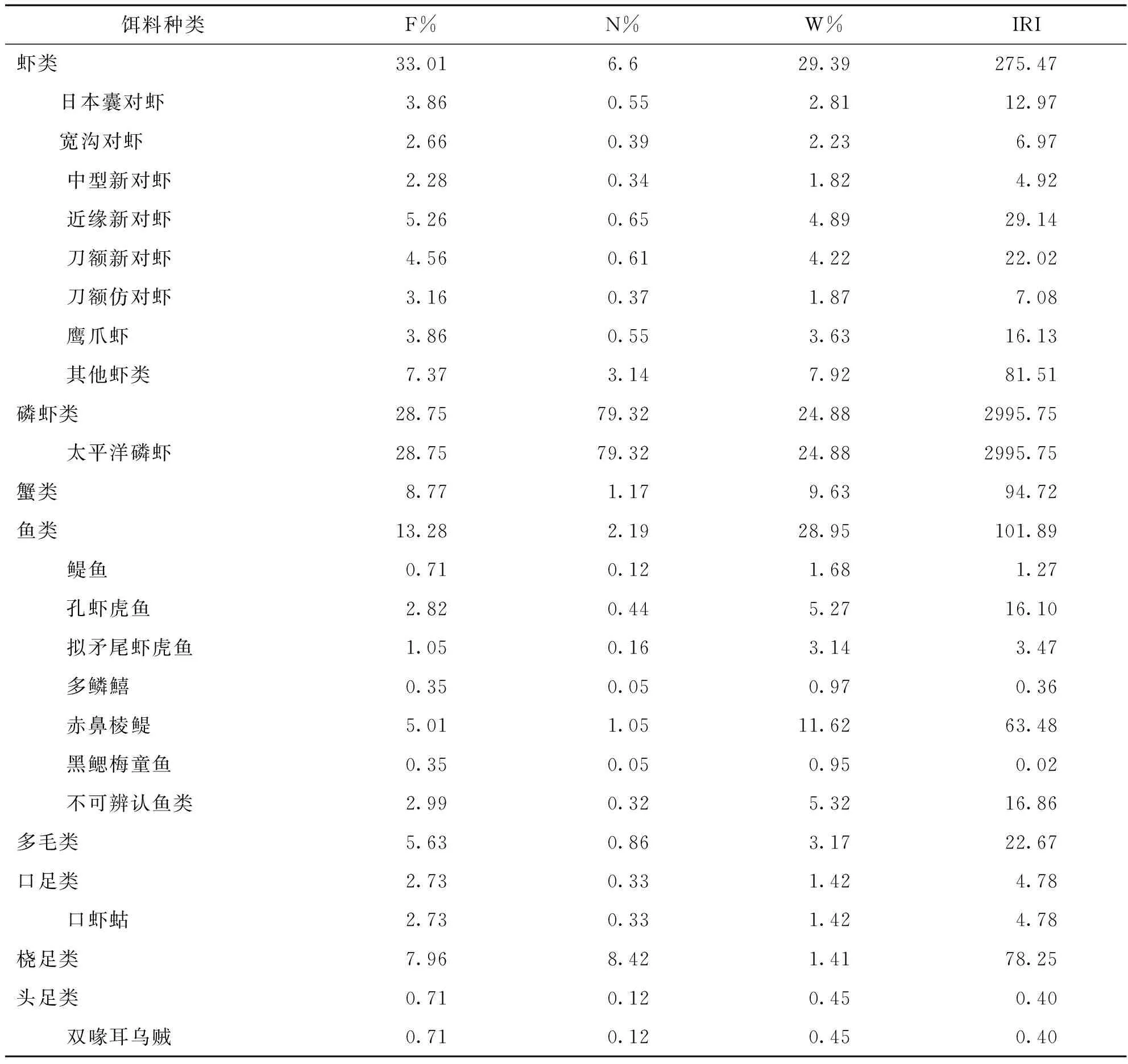
表1 皮氏叫姑鱼的饵料组成
2.5 摄食强度
本文用平均饱满指数、空胃率和充塞度来研究皮氏叫姑鱼的摄食强度.结果显示,皮氏叫姑鱼的空胃率最高值出现在冬季,其次是夏季,春季最小,空胃率的变化具有显著的季节性(X2=14.91,P<0.05).其平均饱满度指数在秋季最高,春季次之,冬季最低(图3).充塞度等级显示,在春季,主要以0级和1级为主;在秋季,充塞度在1级以上的比例最高(图4).
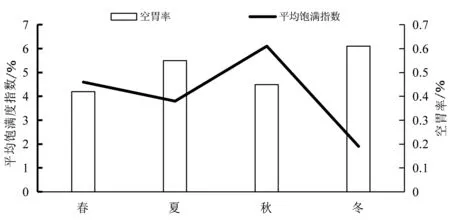
图3 皮氏叫姑鱼的空胃率和平均饱满度指数的季节变化
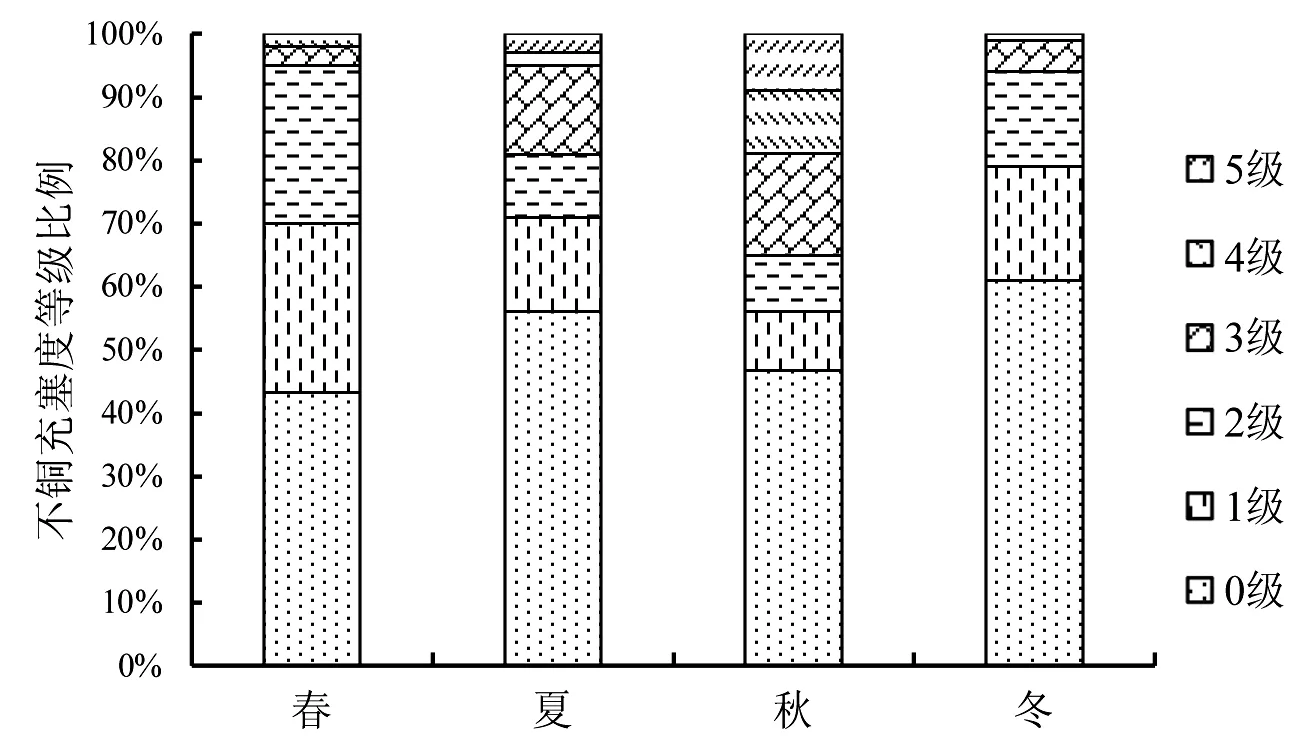
图4 皮氏叫姑鱼不同季节各个充塞度等级所占的比例
2.6 营养级
利用同位素法计算皮氏叫姑鱼营养级,结果显示营养级以2、3级为主,对样本营养级的平均体长进行分析发现,营养级随着体长的增加而增加,其变化范围在2.72-3.36(表2),平均营养等级为3.06.

表2 皮氏叫姑鱼不同体长组的营养级
3 讨论与结论
流沙湾海域皮氏叫姑鱼的体长范围为80-191 mm,其中优势体长组为121-140 mm,而马鞍列岛海域的皮氏叫姑鱼的体长范围为30-264 mm,优势体长组为80-100 mm和130-180 mm[6],这和本文的结果差距较大,这种差异是由于不同地理种群差异所致,还是由于外界环境差异的影响,尚需进一步的研究.皮氏叫姑鱼体长-体重拟合方程中的生长指数b为3.1718,说明流沙湾海域皮氏叫姑鱼为正异速生长的鱼类(b>3),即该鱼体重的增长速度要快于体长的生长速度.流沙湾海域皮氏叫姑鱼的生长指数与珠江口海域的皮氏叫姑鱼的生长指数[14]相比,存在的差异较大,但与福建近海海域的皮氏叫姑鱼的生长指数[10]相似.
流沙湾海域皮氏叫姑鱼的肥满度最大值出现在2月份,最小值在4月份,分别为2.35%和2.12%.6月份开始肥满度呈现上升趋势,直至上升到次年繁殖期前的2月份达到顶峰.这可能是受到性腺发育和摄食量的影响.在秋季时,性成熟的个体开始积累营养以度过饵料生物数量很少的越冬期,因此肥满度在6-10月间是呈上升趋势.冬季时,温度降低,饵料生物很少,肥满度也随之下降.春季是皮氏叫姑鱼的繁殖期,性腺发育成熟,对肥满度造成影响,同时,皮氏叫姑鱼也需要囤积营养以满足繁殖期的营养需求.因此在冬末春初的2月份时肥满度达到最高值.4月份时肥满度下降,可能与此时皮氏叫姑鱼处于性成熟期,摄食强度下降有关[15],也可能是受到捕捞压力的影响[16],具体原因尚需进一步的研究.对于未成熟的个体来说,在生长过程中除了冬季以外,其肥满度基本保持着不断升高的趋势.
饵料组成的分析结果表明,皮氏叫姑鱼的生物饵料种类有19中,主要分为4大类,分别是甲壳类、鱼类、多毛类和头足类.从各饵料生物的相对重要性指数来看,甲壳类是最主要的生物饵料,优势饵料生物种类为磷虾类和虾类;其次是鱼类,主要以赤鼻棱鳀为主;而多毛类和头足类所占比例都较小.从饵料重量百分比来看,皮氏叫姑鱼摄食的主要饵料是虾类、鱼类和磷虾类;从摄食的饵料类群来看,该种鱼的食谱中有出现了底栖动物、浮游生物和游泳动物,因此属混合动物食性鱼类.皮氏叫姑鱼的生物饵料均为流沙湾海域常见的种类,甲壳类是皮氏叫姑鱼的饵料中最重要的类群,出现频率为70.53%.以往的研究表明,南黄海皮氏叫姑鱼是主要摄食底栖动物的肉食性鱼类,主要以虾类居多,其次是鱼类和多毛类[17],马鞍列岛海域的皮氏叫姑鱼成鱼主要摄食虾类和鱼类,繁殖期间主要摄食端足类[6],这和本文的研究结果基本是一致的.
皮氏叫姑鱼的摄食强度有显著的季节性变化.饱和指数和充塞度在1级以上的胃的个数在秋季均达到最高值,而空胃率达到最小值,说明此季节的摄食强度最高.这是由于9月份之后,皮氏叫姑鱼的繁殖期逐渐结束,鱼类个体会在繁殖期结束后补充损耗的能量,同时鱼类需要在冬季之前贮存大量的脂肪度过越冬期,这与其他的研究结果类似的[18-19].春季是繁殖的高峰期,皮氏叫姑鱼此时的摄食强度仅次于秋季,高于夏季和冬季,即处于繁殖旺盛期的皮氏叫姑鱼依旧保持了较高的摄食强度水平.这可能是由于虾类在流沙湾有明显的数量优势,每年3月份到达了虾类的繁殖高峰期,虾类密度上升到顶峰,同时,流沙湾浮游动物的丰富度指数在春季也达到了全年的高峰期[20],因此皮氏叫姑鱼能够较为容易获得饵料.但是通过图4可以看出,在繁殖高峰期春季时,皮氏叫姑鱼充塞度等级在3级以上的个体比例少,仅高于冬季,从图2可以看出,4月份时,肥满度也最低,这可能是卵巢发育影响了摄食行为.林楠等[21]通过对象山港黄姑鱼的食性分析得出类似的观点,认为可能是卵巢发育到4期以后会占据腹腔大量的位置,从而影响了鱼类的摄食量.冬季时,皮氏叫姑鱼的平均胃饱满度指数最低,空胃率最高.可能是低温减低了鱼类的摄食能力,也是饵料生物的数量减少导致的.研究表明,一般鱼类在冬季的摄食量会有所减少[14],这与本文的结果是一致的.
营养级是海洋食物网研究的重要内容,能够反映海洋生态系统动态变化的多种信息[22].鱼类营养级取决于其食性.同一种类不同阶段及个体大小间由于食性的变化,从而导致营养级的变化[23].与胃含物分析相比,氮稳定同位素测定能够更加客观地反应鱼类等消费者的营养级和食性特征[24].本文利用氮稳定同位素计算得到的皮氏叫姑鱼的营养级以2、3级为主,平均营养等级是3.06,属于低营养级的鱼类[25].从摄食种类上来说,低营养级鱼类主要摄食浮游生物和小型底栖动物,这和本文的研究结果是一致的.张波等[25]对崂山湾鱼类摄食生态的研究中发现,崂山湾皮氏叫姑鱼群体的平均营养级为4.11.薛莹等对黄海中南部主要鱼种的摄食生态研究发现,黄海区域的皮氏叫姑鱼营养级在3.5-3.8之间[26].不同群落相同鱼种之间营养级的差别,可能是由于个体大小和摄食饵料生物的不同所导致的[25-26].
[1]孙典荣,李渊,林昭进,等.海南岛近岸海域鱼类群落结构研究[J].中国海洋大学学报(自然科学版),2011,41(4): 33-38.
[2]乔延龙,林昭进,邱永松.北部湾秋、冬季渔业生物群落结构特征的变化[J].广西师范大学学报(自然科学版),2008,26(1): 100-104.
[3]徐兆礼.兴化湾和闽江口海域春夏季鱼类区系特征[J].生物多样性,2011,19(1): 79-84.
[4]Fard N J H,Ravanbakhsh M,Ramezani Z,et al.Determination of mercury and vanadium concentration inJohniusbelangerii(C) fish in Musa estuary in Persian Gulf[J].Marine Pollution Bulletin,2015,97(1/2): 499-505.
[5]贺立静,周述波,李晓梅,等.海南近岸海域污染现状及对生物多样性影响[J].琼州学院学报,2014,21(5): 99-103.
[6]王凯,章守宇,汪振华,等.马鞍列岛海域皮氏叫姑鱼渔业生物学初步研究[J].水产学报,2012,36(2): 228-237.
[7]Jiang L,Su Y,Wu C,et al.Phylogenetic estimation of Sciaenidae in the East China Sea inferred from nuclear EPIC DNA sequence variation[J].Biochemical Systematics & Ecology,2014,53: 69-75.
[8]谢子强,刘雪媚,郭昱嵩,等.北部湾皮氏叫姑鱼的遗传多样性分析[J].南方农业学报,2013,44(1): 140-144.
[9]Azmat R, Talat R, Ahmed K.The Length-weight Relationship, Condition Factor and Impact of Fluoride Concentration inJohniusbelangeriiof Arabian Sea[J].Research Journal of Environmental Toxicology, 2007, 1(3): 138-143.
[10]王家樵,张雅芝,黄良敏,等.福建沿岸海域主要经济鱼类生物学研究[J].集美大学学报(自然科学版),2011,16(3): 161-166.
[11]潘志,冯廷廷,顾志峰.马来西亚吉罗鱼幼鱼生长及形态性状分析[J].海南热带海洋学院学报,2016,23(5):1-4.
[12]Cortés E.A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes[J].Canadian Journal of Fisheries & Aquatic Sciences, 1997, 54(3): 726-738.
[13]Salerno D J, Burnett J, Ibara R M.Age, growth, maturity, and spatial distribution of bluefish, Pomatomus saltatrix (Linnaeus), off the northeast coast of the United States, 1985-96[J].Journal of Northwest Atlantic Fishery Science, 2001, 29: 31-40.
[14]李永振,陈国宝,孙典荣.珠江口鱼类组成分析[J].水产学报,2000,24(4): 312-317.
[15]朱清澄,陈丙见,金鑫,等.缅甸安达曼海浅色黄姑鱼生物学特性的初步研究[J].大连海洋大学学报,2013,28(5):487-491.
[16]李忠炉,金显仕,单秀娟,等.小黄鱼体长-体质量关系和肥满度的年际变化[J].中国水产科学,2011,18(3):602-610.
[17]薛莹,金显仕,张波,等.南黄海三种石首鱼类的食性[J].水产学报,2005,29(2): 178-187.
[18]Xue Y, Jin X B, Liang Z.Seasonal, diel and ontogenetic variation in feeding patterns of smallyellow croaker in the Central Yellow Sea[J].Journal of Fish Biology, 2005, 67(1): 33-50.
[19]Letourneur Y, Galzin R, Harmelin-Vivien M.Temporal variations in the diet of the damselfish Stegastes nigricans (Lacepède) on a Réunion fringing reef[J].Journal of Experimental Marine Biology & Ecology, 2015, 217: 1-18.
[20]王彦.流沙湾浮游生物群落结构与物种多样性研究[D].湛江:广东海洋大学,2010.
[21]林楠,姜亚洲,袁兴伟,等.象山港黄姑鱼的食物组成与摄食习性[J].中国水产科学,2013,20(6): 1284-1292.
[22]Pauly D, Watson R.Background and interpretation of the “Marine Trohpic Index” as a measure of biodiversity[J].Philosophical Transactions of the Royal Society of London B: Biological Sciences, 2005, 360(1454): 415-423.
[23]廖建基,郑新庆,杜建国,等.基于氮稳定同位素的九龙江口鱼类的营养级研究[J].海洋学报,2015,37(2):93-103.
[24]肖协文,王玉玉,张欢,等.饶河枯水期主要鱼类营养级位置及其影响因素[J].生态学报,2015,35(18): 6216-6223.
[25]张波,袁伟,戴芳群.应用稳定同位素技术研究崂山湾夏季鱼类群落的摄食生态[J].水产学报,2016,40(4): 585-594.
[26]薛莹.黄海中南部主要鱼种摄食生态和鱼类食物网研究[D].青岛:中国海洋大学,2005.
(编校:李由明)
Preliminary Study on the Biological CharacteristicsofJohniusbelangeriiin Liusha Bay
GUO Hui, XU Ming, SHEN Yu-chun, YE Ning, CAO Yu-tao
(Key Laboratory of Marine Ecology and Aquaculture Environment of Zhanjiang,College of Fisheries, Guangdong Ocean University, Zhanjiang Guangdong 524025, China)
The biological characteristics ofJohniusbelangerii, including size distribution, relation of body length and weight, condition factor, food composition, feeding intensity and trophic level, were analyzed based on the fisheries data collected in Liusha Bay from January to December 2013.The results showed that the body length ofJohniusbelangeriiranged from 80 to 191 mm, with an average of 139 mm, and the dominant body length group was 121-140 mm.The relationship between body length (L) and weight (W) was expressed as W=0.0134L3.1718(R2=0.9734).The maximum of condition factor was observed in February and minimum in April.Crustacean was the main group in food composition.The percentage of empty stomachs was significantly changed with seasonal variations (P<0.05), and the maximum was observed in winter and minimum in spring.Most of the fish had saturation level of stage 0 and 1 in spring, and rate which above stage 1 reached the peak in autumn.The main trophic levels were 2 and 3, with an average of 3.06, and trophic levels increased with body length.
Johniusbelangerii; biological characteristics; food composition; Liusha Bay
格式:郭慧, 徐明, 申玉春,等.流沙湾海域皮氏叫姑鱼生物学特性的初步研究[J].海南热带海洋学院学报,2017,24(2):7-12.
2017-02-22
广东省普通高校省级重大科研项目(GDOU2013050219);湛江市海洋生态与养殖环境重点实验室(2015A06006)
郭慧(1986-),女,安徽阜阳人,广东海洋大学水产学院讲师,博士,研究方向为水产动物健康养殖生态学. 通信作者:申玉春(1964-)男,内蒙古赤峰人 ,广东海洋大学水产学院教授,博士,研究方向为海洋环境与生物资源保护.
S931.1
A
2096-3122(2017) 02-0007-06
10.13307/j.issn.2096-3122.2017.02.02
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
ViVi美眉(2021年10期)2021-01-25
ViVi美眉(2021年10期)2021-01-25
当代水产(2019年9期)2019-10-08
江西建材(2018年1期)2018-04-04
猪业科学(2016年1期)2016-03-30
新疆地质(2015年3期)2015-12-10
