
泡桐林网系统内小麦产量对光合有效辐射分布的响应*
2017-05-09 08:23王炜炜王保平周海江崔令军苏凌燕
中国生态农业学报(中英文) 2017年5期
赵 阳, 乔 杰, 王炜炜, 王保平**, 周海江, 崔令军, 段 伟, 苏凌燕
泡桐林网系统内小麦产量对光合有效辐射分布的响应*
赵 阳1, 乔 杰1, 王炜炜2, 王保平1**, 周海江1, 崔令军1, 段 伟1, 苏凌燕3
(1. 国家林业局泡桐研究开发中心 郑州 450003; 2. 北京林淼生态环境技术有限公司 北京 100000; 3. 长葛市林业技术推广站 长葛 461500)
本研究以黄淮海平原地区重要的农林复合经营模式泡桐-小麦林网复合生态系统为对象, 通过对2013—2015年系统内光合有效辐射(PAR)的连续定位观测及小麦产量的调查, 结合对小麦不同生育期内的PAR与小麦产量、千粒重、粒数的相关性分析, 研究了系统内PAR的分布状况及小麦产量对其的响应。结果表明: PAR、透光率均随着与林带距离的增加而增加, 且在10 m(约1倍树高)范围内变化显著, 10 m之后增加缓慢。在所有生育期、所有测点中, 透光率的最小值出现在灌浆成熟期2 m处观测点。单位面积小麦产量与小麦全生育期内的PAR、粒数和小麦扬花期内的PAR、千粒重及小麦灌浆成熟期内的PAR的相关关系均达极显著水平(=0.918,=0.000;=0.926,=0.000;=0.922,=0.000)。扬花期林带对小麦的遮荫直接影响小麦的粒数, 灌浆成熟期林带对小麦的遮荫直接影响小麦的千粒重, 系统内小麦产量的空间差异性可以通过小麦粒数和千粒重的差异来解释。小麦产量()与全生育期内PAR()的线性回归方程为:=0.121 3+95.117 (2=0.842)。经检验, 方程的模拟值和实测值无显著差异(=0.609), 预测精度达91.8%。可以根据此方程, 结合PAR观测值对系统内各点的小麦产量进行预测。本研究结果为建立泡桐-小麦林网复合生态系统整体生产力的预测模型奠定了基础, 为优化泡桐林网复合生态系统结构提供理论依据。
泡桐林网; 小麦; 生育期; 光合有效辐射; 透光率; 产量
农林复合系统是有目的地把多年生木本植物与农作物进行合理的时空搭配, 形成的共存互助的生态系统[1]。该系统在协调农林争地矛盾、高效利用各种自然资源、保护生态环境等方面起到了积极作用[2]。有研究表明, 草牧场防护林为牧草的生长创造了良好的小气候环境, 牧草产量高于无林草牧场, 非洲草原上孤立木下的牧草产量也远高于空旷草地[3-4]。但是, 由于木本植物和农作物经常利用同一时空序列上的资源, 导致出现了生态位重叠现象, 产生强烈的竞争作用[5], 二者对水分、营养和光照的竞争也引起了广泛关注[6]。有观点认为, 在复合系统中, 相对于地上部分光竞争而言, 地下部分的水分竞争对作物产量的影响更大, 营养的竞争次之[7-9]。Singh等[10]通过利用根障排除地下部分的竞争作用来研究遮荫是否是导致农作物产量降低的主导因子, 结果发现, 银合欢()对相邻农作物的遮荫率达85%时, 农作物产量仍然没有显著降低, 说明适度的遮荫对农作物产量没有明显影响, 这也进一步说明复合系统中地下部分对水分和养分的竞争是影响农作物产量的主导因子。但美国夏威夷的一项根障试验则显示, 大叶千斤拔[(Willd.) Merr]和玉米(L.)之间的光竞争对玉米产量的影响较为严重[11]。可见根系分布状况在很大程度上影响了水分和养分的竞争状况。浅根性木本植物的根系与农作物的根系基本分布在相同土层, 因此与农作物对水分和养分的竞争就比较激烈[12]。深根性木本植物则不然, 一些深根性木本植物甚至可以将从深层土壤吸收来的水分释放到干旱的上层土壤, 供相邻农作物利用[13-14]。因此, 在设计农林复合系统时, 要考虑到木本植物和农作物在生理或者物候特性上的差异, 减少资源利用的竞争, 实现资源利用的互补[5]。
在黄淮海平原地区, 泡桐[(Seem.) Hemsl.]在每年的4月下旬开始展叶, 至5月中旬叶完全展开, 历时1个月左右; 冬季和春季, 叶面积指数几乎为零, 主要是树冠枝桠的遮荫[15], 对夏作物的影响较小。泡桐以其发叶迟、树冠稀疏、主根深、须根少等生物学特性, 在很大程度上减少了与农作物对光照、水分和养分的竞争[16], 与夏季主要作物小麦形成了农桐复合生态系统经营模式。相关专家针对该系统开展了一系列研究, 基本阐明了系统对小麦(L.)产量的影响[17-20]。但是, 这些研究主要集中在桐麦间作系统, 鲜见泡桐林网内小麦产量、复合系统整体生产力及综合经济效益的报道。
泡桐林网作为农田防护林体系的重要组成部分, 在改善小气候、抵御自然灾害及确保农作物高产、稳产方面起到了重要作用, 是农桐复合系统主要的经营模式。本研究以黄淮海高产农区广泛采用的泡桐-小麦林网复合生态系统为研究对象, 研究了林带对系统内光合有效辐射(PAR)分布及对小麦产量的影响, 建立了林网内PAR与小麦产量的关系, 旨在为建立小麦产量乃至整个系统生产力的预测模型奠定基础, 为优化泡桐林网的结构提供参考依据。
1 材料与方法
1.1 研究区域概况
研究区域位于河南省长葛市南席镇马武村(34.23°N, 114.06°E)。年均气温14.5 ℃, 1月平均气温最低, 为0.1 ℃。7月平均气温最高, 达27.3 ℃。年均无霜期214 d, 降雨量711 mm, 日照时数1 883 h, 日照百分率为54%。土壤为沙壤土, pH、有机质、有效磷、速效钾和全氮含量见表1。
1.2 定位观测林网介绍
定位观测林网网格东西长160 m, 南北宽320 m。结构为东林带1行, 西、北林带1路4行, 南林带为1路2行。2011年3月采用1年生‘泡桐9501’造林, 平均胸径和苗高分别为4.6 cm和3.4 m, 株距4 m。林网网格内种植小麦, 品种为‘豫麦13号’, 播种量为150 kg×hm-2, 统一田间管理。2014年12月调查林木生长状况, 结果见表2。
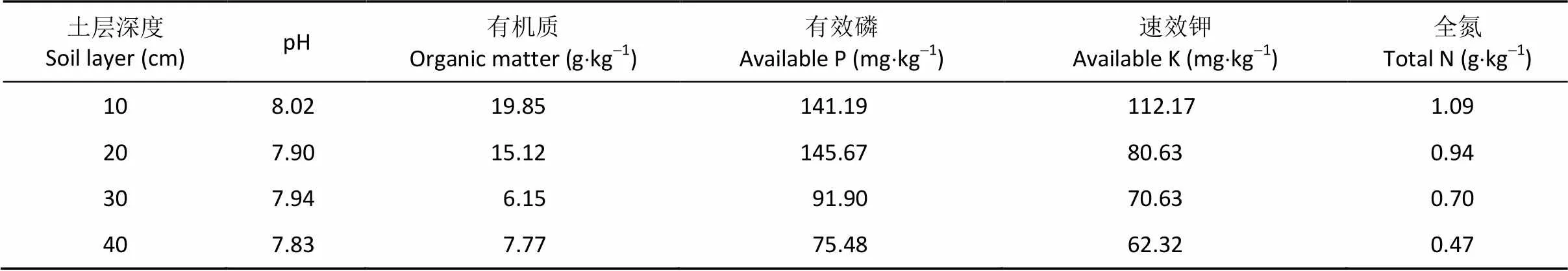
表1 试验地土壤化学基本性状
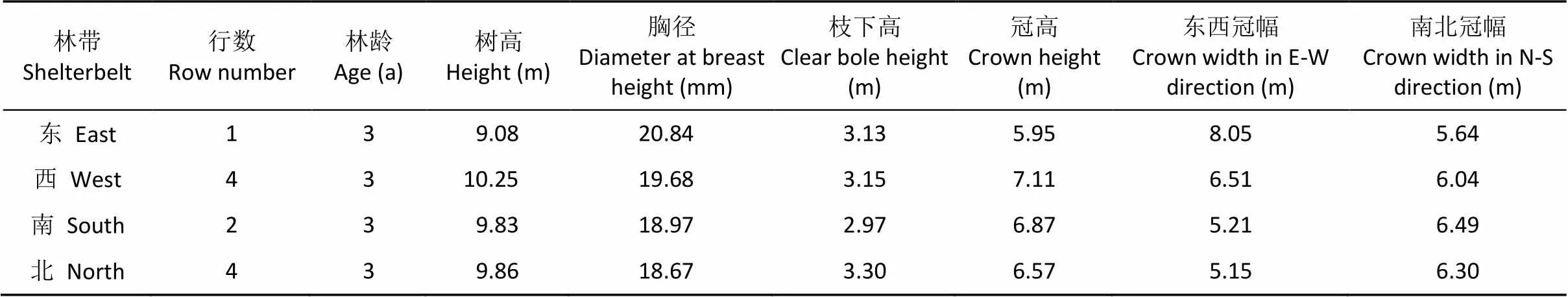
表2 泡桐林网林带基本情况
1.3 观测点的设置
2013年10月1日自东林带中间点开始向西和西林带中间点开始向东2 m、5 m、10 m、15 m、25 m、40 m处各设置1个观测点。自南林带中间点向北和北林带中间点向南2 m、5 m、10 m、15 m、25 m处各设置1个观测点。在林网中间设置2个观测点作为对照, 总计24个观测点。在各观测点分别设置1个高2.5 m, 大小为2.38 Dia.×2.54 cm H的光量子传感器(LI-COR, 美国)。所有传感器均被连接到CR-1000数据采集器上(Campbell, 美国), 每分钟扫描并记录1组数据。每月从数据采集器中导出数据。连续观测3年。
1.4 小麦产量调查
在林网内, 按照距北林带80 m、150 m、230 m和距东林带40 m、70 m、110 m布设6条样带。分别在每条样带与PAR观测点相应的位置设置1个1 m×1 m的样方, 总计72个样方。于小麦蜡熟期采用全收获法收割各样方内的小麦, 人工脱粒之后, 风干。每个样方按照占风干重20%的抽样强度抽取样品在80 ℃条件下连续烘干72 h后称重, 计算各样方内的小麦产量、粒数和千粒重[19]。
1.5 数据处理
依据段伟等[21]日PAR的处理方法, 计算各观测点1 d的PAR, 进而计算影响小麦产量的3个关键生育期[19](扬花期、灌浆成熟期、全生育期)内的PAR和透光率。采用单因素方差分析(ANOVA)和Duncan’s多重比较分析不同观测点之间单位面积的小麦产量、粒数和千粒重的差异。
将2015年各观测点小麦平均产量、粒数和千粒重分别与3个关键生育期内的PAR进行Pearson相关性分析并进行双尾显著性检验, 筛选出相关关系显著且相关系数高的组合分别建立线性回归关系, 研究PAR对小麦产量及其构成因子的影响。将2014年小麦全生育期内的PAR代入建立的回归关系中计算小麦产量的模拟值, 通过与小麦产量实测值进行K-W秩和检验和线性拟合, 检验已经建立的回归关系。
采用Microsoft Excel 2010和SPSS 20.0软件进行数据分析, 采用Origin 9.0软件制图。
2 结果与分析
2.1 泡桐林网对小麦不同生育期光量分布的影响
2.1.1 对小麦不同生育期PAR的影响
位置、季节和时刻的共同作用, 导致太阳高度角不同, 引起了光量子通量密度的变化[22]。图1表明, 在泡桐林网复合系统中, 林带的光竞争作用导致作物所接收的光量子数随着与林带距离的增加而逐渐增加至平稳, 所接收的PAR呈现出相同的变化规律。小麦扬花期、灌浆成熟期和全生育期, 东、西、南和北4条林带结构下, PAR均随着与林带距离的增加而增加, 且增加的速率均逐渐降低, 至10 m之后逐渐趋于稳定。即10 m(约1倍树高)之后, PAR几乎不再受林带遮荫的影响。
东、南、西、北指东林带、西林带、南林带和北林带。E, S, W and N infer east shelterbelt, south shelterbelt, west shelterbelt and north shelterbelt.
2.1.2 对小麦不同生育期透光率的影响
从图2可以看出, 同一生育期内, 透光率随着与林带距离的增加呈增加趋势。距林带10 m范围内对透光率的影响较大。距林带10 m之后, 透光率增加速率逐渐降低。在小麦的扬花期、灌浆成熟期和全生育期, 透光率的最小值均出现在距南林带2 m处观测点, 而且以灌浆成熟期的透光率0.43为最小, 这与此时期泡桐已经进入全叶期有密切关系。
东、南、西、北指东林带、西林带、南林带和北林带。E, S, W and N infer east shelterbelt, south shelterbelt, west shelterbelt and north shelterbelt.
2.2 泡桐林网对小麦产量分布及其构成因子的影响
由表3可以看出, 各观测点单位面积内小麦产量随着与林带距离的变化而变化。方差分析结果显示, 距东林带2 m、5 m处的小麦产量无显著差异, 5 m、10 m、15 m、25 m处的小麦产量无显著差异, 但2 m和5 m处的小麦产量均显著低于40 m处的小麦产量。距西林带5 m、10 m、15 m、25 m 、40 m处的小麦产量无显著差异, 且均显著高于2 m处的小麦产量。距南林带不同距离观测点小麦产量的变化规律与之相同。距北林带2 m、5 m处的小麦产量无显著差异, 5 m、10 m、15 m、25 m处的小麦产量无显著差异, 但2 m处的小麦产量显著低于10 m、15 m、25 m处的小麦产量。
单位面积小麦的粒数和千粒重是小麦产量的2个重要构成因子[19]。距东林带2 m、5 m、10 m、15 m处的小麦粒数无显著差异, 10 m、15 m、25 m、40 m处的小麦粒数无显著差异, 2 m处的小麦粒数显著低于25 m和40 m处的小麦粒数。距西林带2 m、5 m处的小麦粒数无显著差异, 10 m、15 m、25 m、40 m处的小麦粒数与5 m处的小麦粒数无显著差异, 但显著高于2 m处的小麦粒数。距南林带5 m、10 m、15 m、25 m处的小麦粒数无显著差异, 但显著高于2 m处的小麦粒数。距北林带不同距离的5个观测点的小麦粒数无显著差异。
距东林带10 m、15 m、25 m处的小麦千粒重无显著差异, 但均显著高于5 m和2 m处的小麦千粒重。距北林带不同距离小麦千粒重的变化规律与之相同。距西林带2 m、5 m处的小麦千粒重无显著差异, 但均显著低于15 m、25 m、40 m处的小麦千粒重, 10 m与5 m处的小麦千粒重无显著差异, 但10 m处小麦千粒重显著高于2 m处的小麦千粒重。距南林带2 m处的小麦千粒重显著小于5 m、10 m、15 m、25 m处的小麦千粒重, 10 m、15 m、25 m处的小麦千粒重无显著差异。
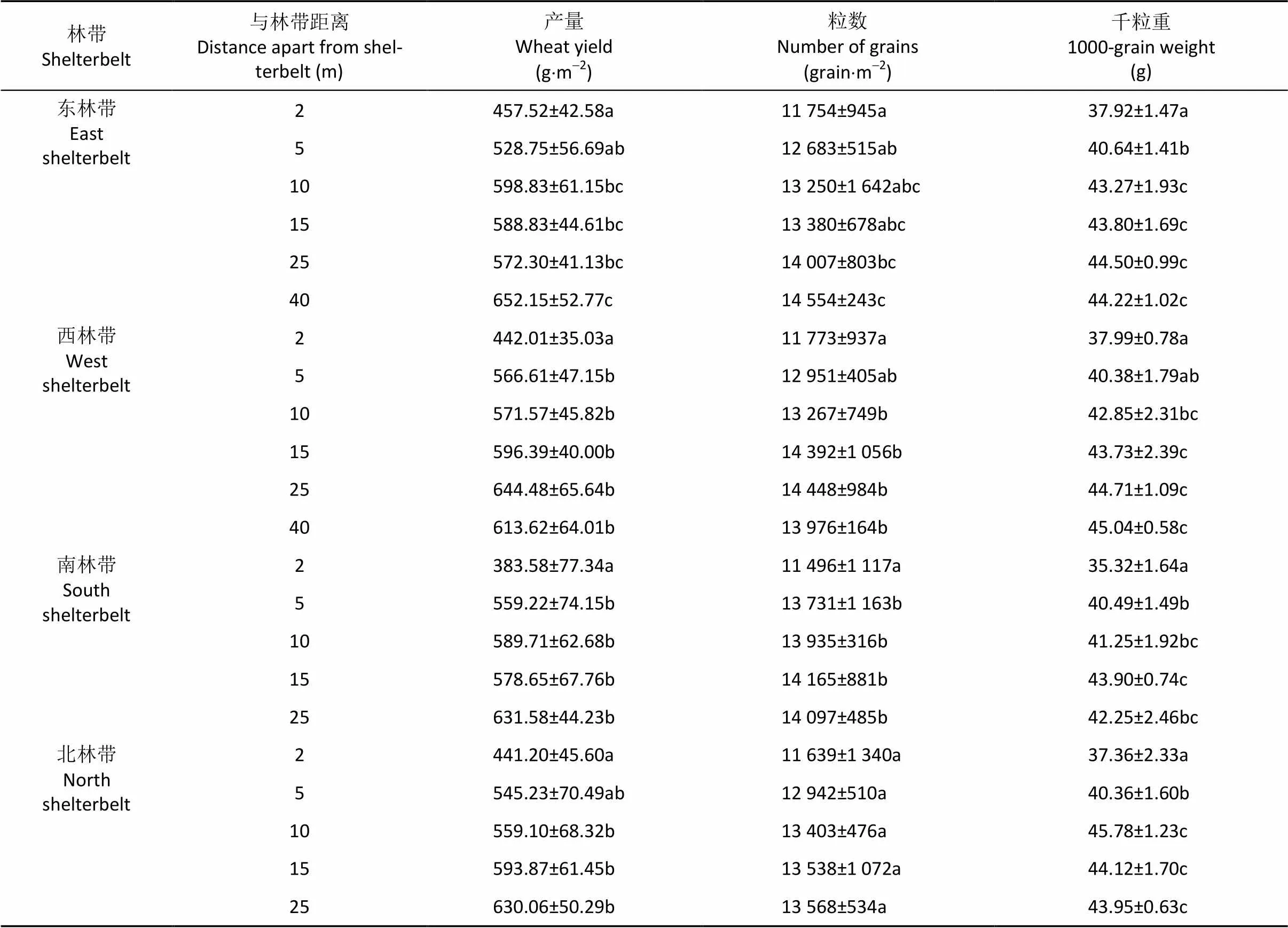
表3 泡桐林带对系统内不同观测点小麦产量、粒数和千粒重的影响
表中数据为平均值±标准差, 同列数字后不同字母表示在0.05水平上差异显著。Data are means±S.E. Values followed by different lowercases in the same column are significantly different at 5% level according to ANOVA analysis and Duncan’s multiple comparisons.
2.3 小麦产量及其构成因子对PAR的响应
由于林带的遮荫, 系统内小麦所接收的PAR发生了规律性变化[23]。泡桐林网系统内各观测点小麦产量之间的差异可以用该点在小麦整个生育期内所接收PAR的差异来解释。即距林带距离越近, 受林带遮荫的影响越严重, 小麦接收的PAR也越少, 产量受到的影响也越严重。小麦在整个生育期内的PAR与小麦产量之间的相关关系达极显著水平(=0.917 4,=0.000)(图3c), 且相关系数高于扬花期内PAR与小麦产量的相关系数(=0.887 3,=0.001)(图3a)、灌浆成熟期内PAR与小麦产量的相关系数(=0.912 1,=0.000)(图3b)。
单位面积小麦粒数与扬花期PAR呈极显著相关关系(=0.926 2,=0.000)(图4a), 且相关系数高于灌浆成熟期内PAR与小麦粒数的相关系数(=0.921 4,=0.000)(图4b)、全生育期内PAR与小麦粒数的相关系数(=0.894 2,=0.000)(图4c)。
小麦千粒重与灌浆成熟期内PAR呈现极显著相关关系(=0.921 7,=0.000)(图5b), 且相关系数高于扬花期内PAR与小麦千粒重的相关系数(=0.917,=0.000)(图5a)、全生育期内PAR与小麦产量的相关系数(=0.920 6,=0.000)(图5c)。
基于上述结果, 分别建立小麦产量()与全生育期PAR()之间的关系(=0.121 3+95.117,2=0.842), 小麦粒数()与扬花期PAR()之间的关系(=11.861+9 295.8,2=0.857 8), 小麦千粒重()与灌浆成熟期PAR()之间的关系(=0.015 5+30.821,2=0.849 6)。PAR与小麦产量线性回归关系方程斜率为0.121 3, 说明PAR每减少1 mol∙m-2, 单位面积小麦产量减少0.121 3 g∙m-2。扬花期间系统内PAR通过影响单位面积内小麦的粒数从而对产量产生影响(PAR每减少1 mol∙m-2, 单位面积小麦粒数减少12粒∙m-2)。灌浆成熟期系统内PAR通过影响小麦的千粒重从而对产量产生影响(PAR每减少1 mol∙m-2, 千粒重减少0.015 5 g∙m-2)
2.4 全生育期PAR与小麦产量之间关系的验证
将2014年各个观测点小麦全生育期内的PAR代入到建立的线性回归方程中, 得到小麦产量的模拟值。将其与实测值进行K-W秩和检验, 结果显示二者之间差异不显著(=0.609)。将各观测点的模拟值与实测值进行线性回归分析, 发现方程的斜率接近于1, 截距接近于0, 说明模拟值和实测值非常接近, 预测精度达91.8%。因此, 可以根据建立的关系, 结合光合有效辐射观测值对系统内各点的小麦产量进行预测(图6)。
3 讨论与结论
造成农林复合生态系统中农作物生产力下降的原因众多, 但在某些情况下, 光能竞争或许是起主导作用的限制因素[24]。本研究表明, 在小麦扬花期、灌浆成熟期和全生育期内, 随着与林带距离的增加, 各观测点的PAR不断增加, 且在10 m(1倍树高)内, 变化幅度较大。10 m之后, PAR增加缓慢。说明林网系统内林带对1倍树高之外光强的影响不再显著, 这与陈振江等[25]的研究结果一致。通过进一步研究透光率的变化发现, 在所有生育期、所有测点, 透光率的最小值出现在灌浆成熟期2 m处观测点。这说明在灌浆成熟期林带对光照的影响最大。这是由于在此期间, 泡桐进入全叶期, 林冠的体积和表面积变大, 对太阳的反射、吸收和遮挡作用增强而导致了这一结果[26]。
林网系统内小麦产量、粒数、千粒重均随着与林带距离的增加而发生变化。小麦产量与其全生育期内的PAR呈极显著正相关关系, 说明泡桐林网复合生态系统中光能的竞争是导致系统内部小麦产量存在空间差异性的主要原因, 这与前期相关的研究结果一致[27-28]。小麦扬花期的PAR与粒数, 灌浆成熟期的PAR与千粒重的正相关关系也均达极显著水平。可以推测, 扬花期林带对小麦的遮荫直接影响小麦的粒数, 灌浆成熟期林带对小麦的遮荫直接影响小麦的千粒重, 系统内小麦产量的空间差异性可以通过小麦粒数和千粒重的差异来解释。
泡桐林网复合生态系统作为黄淮海地区重要的农林业复合经营模式, 不仅可改善农作物的生态环境条件, 还能为产区提供大量的桐材, 在一定程度上缓解农林用地的矛盾。但由于传统的复合生态系统在结构模式等方面缺乏系统研究, 使得复合生态系统的综合效益未能充分发挥。本研究针对泡桐林网复合生态系统建设中存在的关键技术问题, 选择系统内光合有效辐射分布对小麦产量的影响作为切入点, 建立并验证了其与小麦产量之间的关系。这将对建立预测泡桐林网复合生态系统整体生产力的模型提供参考, 为进一步评价泡桐林网复合生态系统的综合效益和优化泡桐林网的结构模式奠定基础。
在泡桐林网复合系统内, 由于林带的遮荫, 系统内的PAR降低, 导致小麦出现一定程度的减产, 泡桐林网未表现出明显的防护效益。这与研究区域在高产农区, 且研究期间未遭受明显的干热风、强对流等灾害性天气有密切关系。如若出现极端灾害性天气, 林网系统就可能会表现出改善农田小气候、保障农作物稳产、高产的功能[4,29]。
优化泡桐林网复合生态系统的结构模式, 需要在发挥林网生态防护效益的同时, 降低林网对系统内农作物产量的影响。减少林带与农作物之间对光的竞争是解决这一问题的有效途径。例如, 通过修枝, 使枝下高升高, 林冠的表面积降低, 可以减少对PAR的截留[30]。合理的株行距与林带宽度也能增加林带的疏透度, 降低林带对农作物的遮荫影响[31]。
[1] 谢京湘, 于汝元, 胡涌. 农林复合生态系统研究概述[J]. 北京林业大学学报, 1988, 10(1): 104–108 Xie J X, Yu R Y, Hu Y. A brief review of agroforestry[J]. Journal of Beijing Forestry University, 1988, 10(1): 104–108
[2] 杨修, 吴刚, 黄冬梅, 等. 农桐复合系统林带结构优化模式的研究[J]. 应用生态学报, 1999, 10(3): 286–288 Yang X, Wu G, Huang D M, et al. Structure optimization of-belt in-crop integrated system[J]. Chinese Journal of Applied Ecology, 1999, 10(3): 286–288
[3] 刘淑玲, 吴德东, 周景荣, 等. 草牧场防护林放牧功能[J]. 东北林业大学学报, 1997, 25(3): 38–44 Liu S L, Wu D D, Zhou J R, et al. Grazing function of the protective forests on grassland[J]. Journal of Northeast Forestry University, 1997, 25(3): 38–44
[4] Belsky A J, Mwonga S M, Amundson R G, et al. Comparative effects of isolated trees on their undercanopy environments in high-and low-rainfall savannas[J]. Journal of Applied Ecology, 1993, 30(1): 143–155
[5] 毛瑢, 曾德慧. 农林复合系统植物竞争研究进展[J]. 中国生态农业学报, 2009, 17(2): 379–386 Mao R, Zeng D H. Research advances in plant competition in agroforestry systems[J]. Chinese Journal of Eco-Agriculture, 2009, 17(2): 379–386
[6] 董宛麟, 张立祯, 于洋, 等. 农林间作生态系统的资源利用研究进展[J]. 中国农学通报, 2011, 27(28): 1–8 Dong W L, Zhang L Z, Yu Y, et al. Resource utilization in agro-forestry intercropping ecosystems[J]. Chinese Agricultural Science Bulletin, 2011, 27(28): 1–8
[7] Jose S, Gillespie A R, Seifert J R, et al. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 2. Competition for water[J]. Agroforestry Systems, 2000, 48(1): 41–59
[8] Jose S, Gillespie A R, Seifert J R, et al. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 3. Competition for nitrogen and litter decomposition dynamics[J]. Agroforestry Systems, 2000, 48(1): 61–77
[9] Hou Q J, Brandle J, Hubbard K, et al. Alteration of soil water content consequent to root-pruning at a windbreak/crop interface in Nebraska, USA[J]. Agroforestry Systems, 2003, 57(2): 137–147
[10] Singh R P, Saharan N, Ong C K. Above and below ground interactions in alley-cropping in semi-arid India[J]. Agroforestry Systems, 1989, 9(3): 259–274
[11] Friday J B, Fownes J H. Competition for light between hedgerows and maize in an alley cropping system in Hawaii, USA[J]. Agroforestry Systems, 2002, 55(2): 125–137
[12] 刘兴宇, 曾德慧. 农林复合系统种间关系研究进展[J]. 生态学杂志, 2007, 26(9): 1464–1470 Liu X Y, Zeng D H. Research advances in interspecific interactions in agroforestry system[J]. Chinese Journal of Ecology, 2007, 26(9): 1464–1470
[13] Richards J H, Caldwell M M. Hydraulic lift: Substantial nocturnal water transport between soil layers byroots[J]. Oecologia, 1987, 73(4): 486–489
[14] Emerman S H, Dawson T E. Hydraulic lift and its influence on the water content of the rhizosphere: An example from sugar maple,[J]. Oecologia, 1996, 108(2): 273–278
[15] 蒋建平, 朱建军, 刘廷志, 等. 桐麦间作系统中小麦产量与光合有效辐射条件的相关变化[J]. 华北农学报, 1994, 9(S1): 133–137 Jiang J P, Zhu J J, Liu T Z, et al. Related changes of wheat yield and photosynthetically active radiation in/wheat intercropping system[J]. Acta Agriculturae Boreali-Sinica, 1994, 9(S1): 133–137
[16] 麻婷婷. 泡桐生长性状及在苏北平原林农间作系统中的应用研究[D]. 南京: 南京林业大学, 2007 Ma T T. Study of pawlownia growth character and the using about pawlownia of interplanting system in Northern Jiangsu Plain[D]. Nanjing: Nanjing Forestry University, 2007
[17] 卢琦, 阳含熙, 慈龙骏, 等. 农桐间作系统辐射传输对农作物产量和品质的影响[J]. 生态学报, 1997, 17(1): 36–44 Lu Q, Yang H X, Ci L J, et al. Effect of radiation transmission on crop yield and quality[J]. Acta Ecologica Sinica, 1997, 17(1): 36–44
[18] 李芳东, 傅大立, 王保平. 桐麦间作系统小麦群体光合量及其与产量的关系[J]. 北京林业大学学报, 1998, 20(3): 108–114 Li F D, Fu D L, Wang B P. Canopy photosynthesis of wheat crops and its relationship with yield in the inter-cropping system of-wheat[J]. Journal of Beijing Forestry University, 1998, 20(3): 108–114
[19] 李芳东, 王保平, 傅大立. 桐麦间作系统内光量分布及其对小麦产量的影响[J]. 北京林业大学学报, 1998, 20(3): 101–107 Li F D, Wang B P, Fu D L. Light distribution within the inter-cropping system of paulownia-wheat and its influences on wheat yield[J]. Journal of Beijing Forestry University, 1998, 20(3): 101–107
[20] 陈振江, 李芳东, 王保平, 等. 平原区农田林网内光量分布及对小麦生物量的影响[J]. 山东农业科学, 2012, 44(10): 40–43 Chen Z J, Li F D, Wang B P, et al. Light distribution in farmland shelterbelt of plain area and its effect on wheat biomass[J]. Shandong Agricultural Sciences, 2012, 44(10): 40–43
[21] 段伟, 王保平, 孟伟, 等. 杨树-玉米林网系统内光强空间变化特征的研究[J]. 中南林业科技大学学报, 2013, 33(12): 113–118 Duan W, Wang B P, Meng W, et al. Characteristics of light intensity in different age poplar forest network system[J]. Journal of Central South University of Forestry & Technology, 2013, 33(12): 113–118
[22] 廖文超, 毕华兴, 高路博, 等. 苹果-大豆间作系统光照分布及其对作物的影响[J]. 西北林学院学报, 2014, 29(1): 25–29Liao W C, Bi H X, Gao L B, et al. Light distribution in apple-soybean intercropping and its impact on the crops[J]. Journal of Northwest Forestry University, 2014, 29(1): 25–29
[23] Li F D, Meng P, Fu D L, et al. Light distribution, photosynthetic rate and yield in a Paulownia-wheat intercropping system in China[J]. Agroforestry Systems, 2008, 74(2): 163–172
[24] 彭晓邦. 光能竞争对商洛低山丘陵区农林复合系统林下作物生产力的影响[J]. 西北林学院学报, 2012, 27(5): 70–75 Peng X B. Impacts of light competition on crop productivity in agroforestry ecosystem in the Hilly Regions in Shangluo[J]. Journal of Northwest Forestry University, 2012, 27(5): 70–75
[25] 陈振江, 李芳东, 王保平, 等. 平原区农田林网光强变化及其对小麦产量的影响[J]. 河南农业科学, 2013, 42(2): 20–25 Chen Z J, Li F D, Wang B P, et al. PAR Change of farmland shelterbelt and its effect on wheat yield in plain area[J]. Journal of Henan Agricultural Sciences, 2013, 42(2): 20–25
[26] 赵忠宝, 万福绪, 刘奕琳, 等. 杨粮复合系统内生态因子的变化及对小麦产量的影响[J]. 南京林业大学学报: 自然科学版, 2008, 32(1): 136–138
Zhao Z B, Wan F X, Liu Y L, et al. Ecological factor changes and wheat output in poplar-crop system[J]. Journal of Nanjing Forestry University: Natural Sciences Edition, 2008, 32(1): 136–138
[27] 彭晓邦, 张硕新. 商洛低山丘陵区农林复合生态系统光能竞争与生产力[J]. 生态学报, 2012, 32(9): 2692–2698Peng X B, Zhang S X. Competition for light and crop productivity in an agro-forestry system in the Hilly Region, Shangluo, China[J]. Acta Ecologica Sinica, 2012, 32(9): 2692–2698
[28] Newman S M, Bennett K, Wu Y. Performance of maize, beans and ginger as intercrops in Paulownia plantations in China[J]. Agroforestry Systems, 1997, 39(1): 23–30
[29] 张改文, 高麦玲, 袁洪振, 等. 黄泛沙地小网格林网对小麦增产效益的研究[J]. 山东林业科技, 2006, (4): 16–17 Zhang G W, Gao M L, Yuan H Z, et al. Study on effects of small network farm-shelter forest in Sandy Area of Yellow River[J]. Shandong Forestry Science and Technology, 2006, (4): 16–17
[30] Ding S S, Su P X. Effects of tree shading on maize crop within a poplar-maize compound system in Hexi Corridor oasis, northwestern China[J]. Agroforestry Systems, 2010, 80(1): 117–129
[31] 赵东. 不同宽度杨树农田防护林带树冠结构特征[D]. 郑州: 河南农业大学, 2011Zhao D. Tree crown structure in relation to width of poplar field shelterbelts[D]. Zhengzhou: Henan Agricultural University, 2011
Response of wheat yield to light distribution in intercroppedshelterbelt and wheat*
ZHAO Yang1, QIAO Jie1, WANG Weiwei2, WANG Baoping1**, ZHOU Haijiang1, CUI Lingjun1, DUAN Wei1, SU Lingyan3
(1. China Paulownia Research Center, Zhengzhou 450003, China; 2. Beijing Linmiao Eco-Environment Technology CO. LTD, Beijing 100000, China; 3. Changge General Station for Forest Technology, Changge 461500, China)
-wheat intercropping system is one of the most important agro-forestry ecosystems in Yellow-Huai-Hai River Flooding Plain in China. In this paper, ashelterbelt intercropped with wheat was studied in order to determine light distribution and its impact on wheat yield. A total of 24 observation points with different differences to.shelterbelt were set to monitor photosynthetically active radiation (PAR) and 72 quadrats investigated to measure wheat yield, 1000-grain weight and grains per unit area for the period 2013–2015. Correlations between PAR at 3 key stages (including flowering stage, grain-filling stage and whole growth stage) and wheat yield, 1000-grain weight and grains per unit area were then analyzed. The results suggested that PAR and transmittance increased along with increasing distance fromshelterbelt and there were significant changes within 10 m (approximately 1-fold tree stem height) in theshelterbelt. PAR and transmittance changed slightly above the distance of 10 m apart. The transmittance at 2 m apart from the south of the shelterbelt at grain-filling stage was smallest among all the observation points at 3 growth stages. The correlations were significant between wheat yield and PAR for whole growth period (= 0.918,= 0.000), the number of grains and PAR at flowering stage (= 0.926,= 0.000), the 1000-grain weight and PAR at grain-filling stage (= 0.922,= 0.000). The number of grains per unit area and 1000-grain weight were directly affected by shelterbelt overshadow both at flowering stage and grain-filling stage. The spatial difference in wheat yield in the intercropping system was explained by the difference in number of grains per unit area. The linear regression equation for wheat yield () and PAR () was= 0.121 3+ 95.117 (2= 0.842). The K-W rank test (= 0.609) at equation precision of 91.8% showed no significant difference between predicted yield and actual yield. The equation (in combination with observed PAR) was suitable for predicting wheat yield in the intercropping system. This study provided theoretical basis for establishing predictive model of ecosystem productivity and for designing optimal structure of-wheat intercropping system.
shelterbelt; Wheat; Growth period; Photosynthetically active radiation; Transmittance; Wheat yield
S344.2
A
1671-3990(2017)05-0647-09
10.13930/j.cnki.cjea.160887
* 国家科技支撑计划项目(2015BAD07B00)资助
**通讯作者:王保平, 主要从事高效森林培育及农林复合经营技术研究。E-mail: bpingwang@163.com
赵阳, 主要从事农林复合经营技术研究。E-mail: zhao5980@126.com
2016-10-09
2017-01-09
* This work was funded by the National Key Technologies R&D Program of China (2015BAD07B00).
** Corresponding author, E-mail: bpingwang@163.com
Oct. 9, 2016; accepted Jan. 9, 2017
赵阳, 乔杰, 王炜炜, 王保平, 周海江, 崔令军, 段伟, 苏凌燕. 泡桐林网系统内小麦产量对光合有效辐射分布的响应[J]. 中国生态农业学报, 2017, 25(5): 647-655
Zhao Y, Qiao J, Wang W W, Wang B P, Zhou H J, Cui L J, Duan W, Su L Y. Response of wheat yield to light distribution in intercroppedshelterbelt and wheat[J]. Chinese Journal of Eco-Agriculture, 2017, 25(5): 647-655
猜你喜欢
现代农村科技(2021年2期)2021-03-15
农业工程学报(2020年11期)2020-07-22
绿色科技(2019年13期)2019-08-31
绿色科技(2019年12期)2019-07-15
现代园艺(2017年13期)2018-01-19
浙江林业(2017年8期)2017-11-13
科技视界(2016年19期)2017-05-18
安徽林业科技(2015年6期)2015-09-25
中小企业管理与科技·中旬刊(2014年10期)2015-02-03
吐鲁番(2014年2期)2014-02-28
