
咸丰县楠木种群结构及其群落特征研究
2017-03-28 13:49吴月淼于鹏宇周明芹
湖北林业科技 2017年1期
吴月淼+于鹏宇+周明芹
摘 要: 为保护和合理利用楠木这一珍稀自然资源,采取样方法,调查咸丰县境内5个乡镇的楠木群落,并记录其生境条件、群落特征,再逐一分析各样地楠木落的群落结构特征、各生境因子对其分布的影响。结果表明:调查发现的5处样地中,有2个种群为增长型,3个为衰退型;楠木群落最适宜在阴坡的中坡坡位生长。
关键词: 楠木;群落分析;种群特征;咸丰
中图分类号:S792.24 文獻标识码:A 文章编号:1004-3020(2017)01-0038-06
Abstract: Using the method of plots sampling, we investigated the community of Phoebe zhennan within 5 towns, which are situated in Xianfeng county, Enshi city, Hubei province , in order to protect the natural resource of P.zhennan and make rational use of it. We record and analyzed the effects of ecological fators and community characteristics and the result showed that among the 5 places there were 2 incremental communities and 3 decaying communities. Besides, the middle slope with shady orientation, is the most suitable living environment of P. zhennan communities.
Key words: Phoebe zhennan;community analysis; population characteristic; Xianfeng prefecture
楠属Phoebe和润楠属Machilus植物为樟科常绿乔木,天然分布于我国长江以南各地,在湖北省的主要分布种有楠属7种,润楠属4种[1],其中楠属有国家二级保护植物3种,润楠属有国家二级保护植物1种[2]。楠木P.zhennan素以材质优良而著名,其木材质地坚硬,纹理细密,削面光滑美观,芳香均有光泽;不易开裂起翘,不易腐朽,为上等建筑、家具、雕刻和文玩的优良用材。楠木树形高大秀丽,枝叶繁茂,四季常青,又是理想的园林绿化树种。此外,它也是抗性较强的深根性树种,可作为改善、修复生态环境的重要生态树种。因此,楠木是具备经济价值、美化价值、生态价值能于一体的优良阔叶树种。
然而楠木因其优良的材质,在20 世纪中期被大量砍伐,种群数量锐减,群落斑块缩小,天然大面积的野生楠木林难以见到。不仅其种群数量资源日渐减少,其群落生态环境也因人类的放牧、开垦等活动而受到强烈干扰,变得越来越脆弱。加上楠木种子存在休眠,在自然状态下解除休眠的时间较长,种群更新速度慢等原因,加上人为的采伐和干扰,使得楠木的数量越来越少,尤其是野生资源,目前已处于濒危状态。在现存的楠木资源中,闽楠P. bournei、湘楠P. hunanensis等多作为风水林被保存下来而散生于村边或祠庙旁[3]。对楠木的种群和群落进行调查研究,对于了楠木濒危原因、提出科学合理的保育对策具有重要意义。
1 咸丰县自然地理概况
咸丰县位于湖北省西南部,是恩施州辖县。地域范围在东经108°37′8″~109°20′8″,北纬29°19′28″~30°2′54″之间。县国土总总面积为2 550 km2,其中林地占77.3%,面积19.7×104 hm2;境内山峦众多,沟壑纵横。境内海拔最高点1 911.5 m,最低点445 m,平均海拔在800 m左右[4]。咸丰县地处亚热带季风气候区,四季气候变化分明。年均温为10~16 ℃,极端高低温分别为37.6 ℃和-13 ℃;全县年均降水量1 460 mm,最大降水量2 353.9 mm,雨热同期,降水主要集中在4~9月[5]。土壤类型主要有棕壤、黄棕壤、黄壤和水稻土,多为微酸性,pH值为5.5~6.5;有机质平均含量高[6]。咸丰森林覆盖率为77.2%,已发现并记录的草本植物有160多科900多种,拥有珙桐Davidia involucrata var.involucrata、红豆杉Taxus chinensisi.var.mairei等10多种珍稀树种群落。海拔高于1 400 m的地区,以常绿或落叶硬阔叶林为主,针叶林及软阔叶林为辅;海拔在800 m到1 400 m内的地区,多以针叶林为主,阔叶林为辅;海拔低于1 000 m的黄壤土地带,以柏木林为主[7]。
调查样地有咸丰县辖内的:黄金洞镇、活龙坪乡、尖山乡、清坪镇、小村乡。
2 调查方法
针对湖北省咸丰县的楠木调查,采用样方法[8-9]进行调查。在其所处的群落中选择具有代表性的地段设置主样方。并对生境情况、植株整体形态、植株叶片形态进行拍照存档。楠木群落主样方设置边长L为20 m,面积为20 m×20 m的正方形。在特殊地形下也可设置为长方形,但长方形的最短边长不小于5 m。
2.1 调查内容
2.1.1 目的物种调查
对于调查范围内的楠木,采取“每木记账法”,记录楠木的株数、株高、胸径、健康等级及幼树数量,伴生种的种类。
2.1.2 生境调查
逐项调查楠属植物所处的生境类型:植物群落名称、郁闭度、盖度;海拔高度、坡度、坡向、坡位和土壤类型;人为干扰方式与强度。
3 调查结果
3.1 楠木种群特征
3.1.1 年龄结构
植物种群的年龄结构,指的是种群内各年龄或年龄组的个体数,所占整个种群总个体数量的百分比。年龄结构能够反应林木更新过程的长短,和更新速度的快慢。在分析种群的动态和对种群进行预测时,采用较多的是研究种群年龄结构的方法。由于实际研究条件的限制,难以通过植物的外部特征,准确地确定楠木的树龄,因此,实践中常以立木的胸径结构代替年龄结构,来进行分析研究[10]。
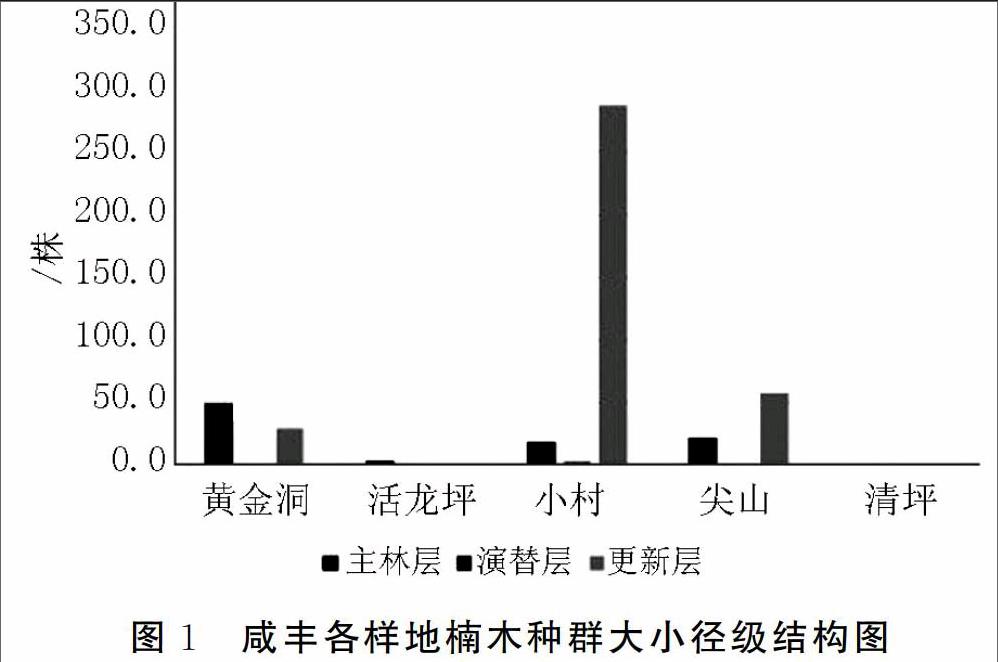
根据楠木胸径的大小,将其划分为5个等级:Ⅰ级幼苗,树高(H)<100 cm;Ⅱ级幼树,胸径(DBH)<3 cm;Ⅲ级立木(3 cm≤DBH<12 cm);Ⅳ级立木(12 cm≤DBH<20 cm);Ⅴ级立木(20 cm≤DBH)。
从图1可以看出,活龙坪(3株)和清坪(古树1株)不具备完整的种群结构,仅有少量成年立木存在,林下无幼树、幼苗,幼年个体无法在未来一定时期内,补充衰老个体的死亡数量,种群为衰退型;位于黄金洞的楠木种群I级幼苗(29株)和V级立木(24株)数量相当,林内无II级幼树(0株),从III级立木(10株)、IV级立木(15株)到V级立木(24株)数量逐渐增加,说明种群的更新能力正在减弱,且自然状态下,I级幼苗向II级幼树转化的过程中,会出现大量的损耗[2],因此,该种群为衰退型;样地位于小村和尖山的种群,虽然II级幼树数量很少,但有充沛的I级幼苗储备,尤其是位于小村的楠木种群,I级幼苗数量达到287株;从径级结构图来看,小村和尖山的楠木种群中,幼年个体明显多于成年和衰老个体,种群更新力旺盛,这两处的楠木种群为增长型。
3.1.2 高度结构
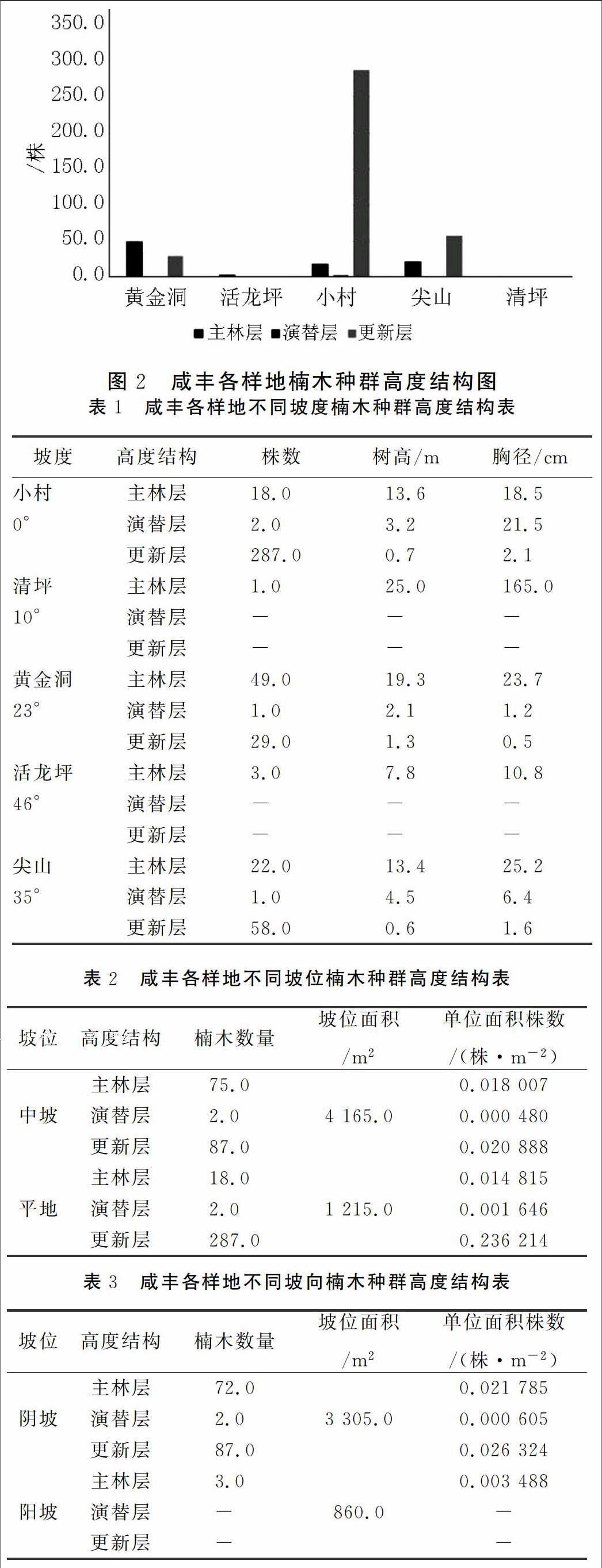
乔木的高度结构,是指在不同高度级内植株所占的数量,该指标也可在一定程度上反映年龄结构。将楠木种群,按照个体高度分为:主林层(植株高度大于5 m)、演替层(植株高度在1~5 m之间)以及更新层(植株高度小于1 m)。
从图2中可以看出,按照高度结构来划分,活龙坪和清坪的楠属植物种群都只具备主林层,为典型的衰退型;黄金洞的主林层数量最高(49株),演替层最少(1株),更新层数量较主林层少(29株),成年树木远多于幼年树木,种群更新能力低下,为衰退型;样地位于小村和尖山的楠木种群,虽然演替层数量较少,但有充沛的更新层储备,尤其是位于小村的楠木种群,更新层数量达到287株,显著多于成年个体(与图1一致),楠木种群为增长型。
3.2 不同生境因子对楠木群落的影响
汇总调查数据,依据不同坡向、坡位、海拔、不同群落单位面积,统计并分析比较不同生境条件下的楠木种群数量,以及不同生境因子与分布数量之间的关系。
3.2.1 坡度
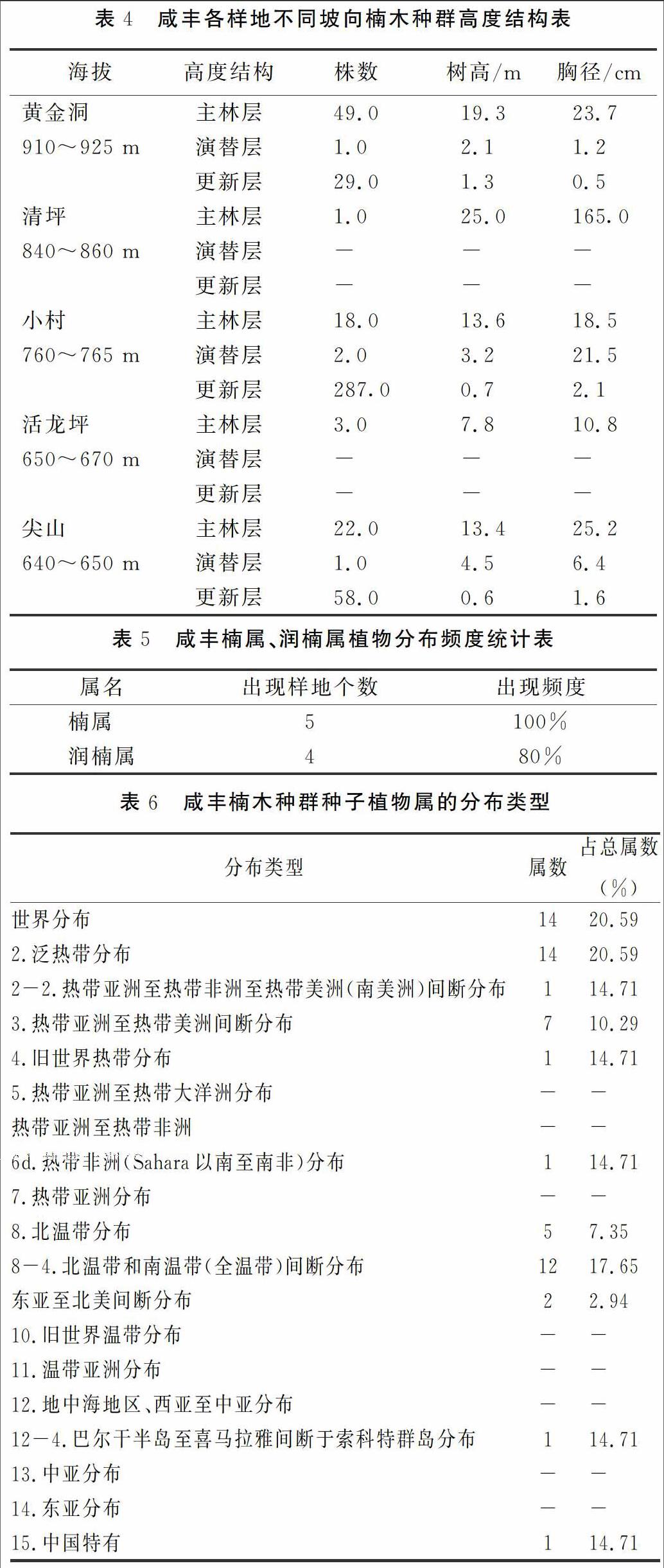
不同坡度对楠木种群的分布会产生一定影响。坡度为0°时,楠木种群具有大量的更新层,但主林层数量不高;坡度在10°和40°时,调查样地内的楠木种群仅有主林层存在,林下无更新层和演替层;坡度在23°和35°时,楠木种群具备完整的主林层、演替层、更新层结构;不同的是,23°时,主林层数量最大,更新层和演替层次之,群落呈现衰退型;坡度为35°时,群落内更新层数量最高,为增长型。
3.2.2 坡位
通过表2可以发现,除尖山样地为平地外,其余楠木种群均分布于中坡坡位。中坡位的更新层单位面积株数(0.020 888株/m2)少于平地的单位面积株数(0.236 214株/m2);演替层在中坡位的分布数量与平地的一致,均为2株,但单位面积株数小;但主林层则差异不大,二者均接近0.016株/m2。
3.2.3 坡向
在调查样方4 165 m2的范围内,共分布楠木633株,其中阳坡3株,阴坡323株(另:平地307株)。 实测单株最大胸径192 cm,树高 23 m,为楠木古树。通过统计在不同的坡向和不同层次(更新层、演替层、主林层)楠木的数量,从表3中可以发现,阳坡的楠木的分布数量很稀少,仅主林层3株;总阴坡环境的单位面积株数为0.048 714株/m2远高于阳坡的单位面积株数0.003 488株/m2。阴坡环境下,楠木群落的结构也较为完整。
3.2.4 海拔
在咸豐县的5处样地中,分布在不同海拔高度的楠木种群之间,相互存在着差异(表4)。更新层在760~765 m分布最多,有287余株;演替层在各海拔高度数量均极少,在调查区域内,随着海拔的降低,演替层楠木的树高和胸径均有增加;主林层则在海拔高度为910~925 m分布最多,有58株,且树体高大粗壮。
3.3 咸丰县楠木群落特征
3.3.1 楠属、润楠属植物出现的频度
调查中,共发现楠属植物4种、润楠属植物2种。从表5可以看出,咸丰县调查范围内的5个样地中,楠属植物出现的频度为100%,其中楠木出现频度为100%,山楠P.chinensis、闽楠、竹叶楠P.faberi均仅出现1次,出现频度为20%;润楠属植物出现频度为80%,其中润楠M.pingii出现在4个样地中,出现频度为80%,但其在各样地中均为零星分布,华润楠M. chinensis仅出现在1个样地中,出现频度为20%。
3.3.2 群落种类组成统计
咸丰楠木群落共有80种,隶属46科68属。通过统计楠木群落维管束植科的属种组成,可看出,樟科Lauraceae物种数最多,有3属7种,占总科的8.75%;其次为百合科Lilaceae、胡桃科Juglandaceae以及壳斗科Fagaceae。其中蓼科Polygonaceae、柿科Enemaceae等的组成,都是单属单种,约共占总科的60.87%,约占总属的41.18%,占总种数的35%,也是重要的组成部分。总体来说,咸丰县楠木群落植物区系成分比较复杂,植物组成多样。
3.3.3 植物区系分析
植物区系既是组成一个地区植被的基础,也是该地区在一定历史条件、自然地理条件下,综合作用促使植物长期演化形成的结果[11]。
咸丰楠木群落有种子植物 59属。根据《中国种子植物属的分布区类型》[12]进行分区。结果从表6可以看出,咸丰楠木群落中植物属的成分只有9个分布区类型和2个变型,且多集中在泛热带分布、泛热带分布和全温带间断分布,共有40属,约占总属数的58.82%。其中楠木所隶属的樟科,为泛热带分布植物。从大类来讲,分布比较多的为热带分布,共有24属,约占总属数的35.29%,其中世界分布和泛热带分布是中最大一类,各占总属数的20.59%。整个群落区系分布,呈现出从亚热带逐渐向温带过渡的趋势,这与咸丰的地理方位一致。
3.4 生境概况统计
通过记录楠木群落的生境情况,发现咸丰县的楠木分布在东经108 °42′~109 °06′,北纬29 °49′~29 °53′的范围内,海拔高度在640 m~925 m之间;土壤类型为山地黄红壤和山地黄棕壤,pH值在主要在4.4~7.5的范围内,活龙坪样地土壤pH值<4.5,整体土壤呈酸性。群落类型均为常绿阔叶混交林,郁闭度45%~88%,盖度63%~93%;分布在房前屋后、农田附近,多在靠近道路、水源的山坡上,以阴坡为主;5处样地的群落受人为干扰严重,主要干扰方式为采集和建设。
4 小结与讨论
4.1 楠属、润楠属植物新旧记录种比较
对比《中国植物志》、《中国植物图鉴》、《湖北植物大全》中楠属、润楠属植物在湖北资源的分布情况,发现在调查区域内发现4种楠属植物、2种润楠属植物中,4种楠属植物均有记载,而调查发现的润楠属植物:华润楠M. chinensis、润楠M. pingii,在已有的文献资料中是没有记载的,属于新记录种。并且,在整理研究中也发现,樟科楠属、润楠属植物系统学研究现状中,仍然存在一定的混乱。本文以《中国植物志》为分类和物种鉴别的主要参考依据,但在《湖北植物志》中,则存在将润楠属Machilus记载为桢楠属Machilus、楠属Phoebe记载为楠木属Phoebe;且属间分类混乱,将小果楠M.microcarpa、宜昌楠M.ichangensis等归类到桢楠属Machilus下。不同出版物之间存在分歧矛盾,尤其是各地方的植物志,说法不一,给研究调查工作,带来了阻力。针对楠属、润楠属植物之间的物种鉴别、属间区分,现在也有大量的研究展开[13]。除了通过传统的形态表征、生理生化指标等进行鉴别,目前较多的是从分子水平层面展开研究,对种源关系的远近进行鉴定,从基因层面上更加可靠地进行区分[1415]。
4.2 咸丰楠木种群特征
咸丰县5个乡镇的调查样地中,总共有3处的楠木种群为衰退型,2处为增长型。该结果与全国范围内,楠属、润楠属植物濒危的现状一致。咸丰县现存的楠木种群,受到较强的人为干扰,种群结构不完整,衰退型楠木种群仅有V级立木存在;增长型的种群也在II级幼树出现断层,种群更新压力大,若持续受到人为干扰破坏,导致现存的大量I级幼苗死亡,则较易转变成为衰退型。
4.3 咸丰楠木群落植物组成与种子植物区系划分
根据调查资料统计,咸丰楠木群落内共有植物80种,隶属46科68属。物种数最多的是樟科,有3属7种,占总科的8.75%;科组成是单属单种的,共占总科的60.87%,占总属的41.18%,占总种数的35%。整体上,咸丰县楠木群落植物区系成分比较复杂,植物组成多样。这与胡蝶[16]等人对恩施南方红豆杉天然林的群落及种群特征研究中,植物区系划分的研究结果一致。
4.4 不同生境条件对咸丰楠木群落分布的影响
从调查数据的分析结果来看,楠木群落在阴坡23°~35°的中坡坡位生长较为良好,而与调查范围内的海拔高度并无显著关系。分析其原因主要是由于楠木的群落主要分布在水源充足的地段,楠木作为深根性树种,在中坡坡位能够更好地获得水分;较低坡位的种子和幼苗,容易因受汛期水位的升高而难以存活或发育成主林层,且无论是在人居环境还是自然环境中,临近水源地带都是动物活动较为频繁的地段,幼苗容易受到踩踏、破坏。总体上,楠木群落在阴坡坡向分布更多,推测是与其幼苗的耐荫性强有关。这與对同为楠属植物的人工林研究结果一致[17]。在光照不充足的情况下,植株依然能够健壮生长,从而在严酷的自然环境中争夺到生存空间。且楠木童期生长缓慢,一旦达到成林高度,则会迅速增大增粗,获得更多的生存资源。而坡度在23°~35°条件下,楠木群落数量和结构上都较优,或许是与种子的保留程度有关。楠木种子量巨大,但存在休眠。掉落在坡度较高的位置,则容易顺势滚落,有的随降雨被流水冲走,难以形成规模分布的楠木林;而咸丰的楠木均分布在房屋前后、水边路旁,平坦的地段人类和家禽家畜的活动较为密集,对Ⅰ级幼苗和Ⅱ级幼树的影响很大,也难以成林出现。总的来说,楠木在阴坡,坡度在23°~35°的中坡坡位环境下,生长良好。这与前人关于楠属植物自然群落和闽楠人工林的研究结果一致[18-21]。
4.5 自然条件下楠木渐危的原因分析
通过以上图表分析和调查,结合其他濒危物种的研究,总结出发现楠木至危因素主要有两方面:
4.5.1 生境破坏导致的种群斑块缩小
其中人为因素为最主要的因素。由于楠属植物抗性强、种子量大,其对自然气候变化导致的生境变化适应力较强,自然因素并不是其生境破碎的主要原因。而人类为追求其木材优良的质地,而进行破坏性的砍伐,导致种群数量极大地减少,群落结构也被破坏;人类为了自身的发展和建设,进行开荒筑路、开垦放牧等活动,也破坏了楠木的生境、缩小了群落斑块,加剧了楠木境破碎化。虽然楠木繁殖力高、种子量大,但种子休眠期长、休眠过程中会因种子失活而产生损耗,幼苗生长缓慢,凭借自身力量难以在短期内修复生境的破碎化,进而导致了遗传漂变等一系问题,进一步加重了植物濒危程度,陷入恶性循环。
4.5.2 楠木从I级幼苗向II级幼树转化损失量大
由于楠木在从I级幼苗向II级幼树的转变过程中,数量损失巨大,造成群落更替障碍。张文辉在分析总结濒危植物的种群生态特征和至危因素中,也提出了这一论点[22]。通过上文的图表分析不难发现,无论何种样地条件,楠木种群的II级幼苗数量都非常稀少,即使在数量较庞大的种群中,也与其他立木级别不成比例;按高度划分来分析,演替层近乎处于断带的状态。这一现象阻碍了楠木种群的更替和发展。然而人类的活动也加剧了这一现象,导致活龙坪和清坪两处样地仅有主林层存在。由于大部分楠木分布在人类活动较为频繁的地段,部分楠木群落或被作为当地风水林而得到保存。但人类主要保护的是已经成林的主林层,进林拾柴、穿行等均会对林下幼苗造成影响,也会影响到促进楠木传粉、种子传播的动物和昆虫的活动;甚至会有村民挑选长势瘦弱的植株当做薪柴进行砍伐,直接阻碍了演替层的形成和发展[2]。
4.6 楠木资源的保育对策
由于楠木童期较长、成树生长缓慢、加之现在社会对经济价值的认知正在逐步提高,所以应加大保护管理力度。定期巡查并记录其生长状况,尽量以避免因病虫害等原因导致的自然损耗。在每年果期,可对野生楠木成熟的种子进行采集,部分优质种子作为野生种质资源进行保存,另一部分健康的楠木种子用于人工培植。培植出的幼苗可以用于开发利用、补充到楠木群落个体数量,以扩大种群数量,提高群落稳定性。同时可通过引种繁育,进行人工培植驯化,判断其是否适应于咸丰的地理气候和生物环境,再选择适宜的生境进行少量的野外种植。以丰富咸丰县的野生植物种群,扩大野生保护植物资源库。
针对较为贫困的地区,缺乏基层技术人员和资金等原因,难以进行濒危树种楠木的保育工作。则可以针对其优良种质资源分布,与林业部门或科研单位联合立项,建立专门的保育项目。
参 考 文 献
[1]中国科学院中植物志编辑委员会.中国植物志[M].北京:科学出版社,1982,(31):714,89116.
[2]中华人民共和国国家环境保护局.中国珍稀濒危保护植物名录[M].北京:科学出版社,1987,(7):2328.
[3]葛永金.闽楠栽培生理生态基础研究.[D].北京:北京林业大学,2014.
[4]咸丰在线.咸丰概况[EB/OL].(20160306)[20160602].http://www.0718xf.com/bendi/info17388.html.
[5]咸丰在线.咸丰县气候特征. [EB/OL].(20160306)[20160602].http://www.0718xf.com/bendi/info19276.html.
[6]咸丰县.咸丰县土壤志[M].咸丰: 咸丰县土壤普查办公室,1984: 224242.
[7]咸丰县.湖北省咸丰县地名志[M].咸丰: 咸丰县地名办公室,1984: 344345.
[8]周纪伦,郑师章,杨持.植物种群生态学[M].北京: 高等教育出版社,1993: 2832.
[9]王伯荪,李鸣光,彭少麟.植物种群学[M].广州: 广东高等教育出版社,1995: 3541.
[10]徐化成,范兆飞,王胜,等.兴安落叶松原始林林木空间格局的研究[J].北京: 生态学报,1994,14(2):155160.
[11]陈灵芝.植物区系与植被地理[M].北京: 科学出版社,2014,12: 3.
[12]吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991(增刊):1139.
[13]陈俊秋.樟科润楠属植物系统学研究.[D].西双版纳: 中国科学院研究生院(西双版纳热带植物园),2008.
[14]沈雪梅.17种樟科润楠属植物的数量分类与分子分类研究.[D].上海:华东师范大学,2015.
[15]陈俊秋,李朗,李捷,等.樟科润楠属植物ITS序列贝叶斯分析及其系统学意义[J].云南植物研究,2009,31(2):117126.
[16]胡蝶,沈瑾花,费永俊.恩施南方红豆杉天然林的群落及种群特征[J].湖北农业科学,2016,55(1): 100107.
[17]江香梅,肖复明,叶金山,等.闽楠天然林与人工林生长特性研究[J].江西农业大学学报,2009,31(6):10491054.
[18]吴大荣,吴永彬.闽楠种群的天然更新[J].植物资源与环境学报,1998,7(3):812.
[19]吴大荣,朱政德.福建省羅卜岩自然保护区闽楠种群结构和空间分布格局初步研究[J].林业科学,2003,39(1): 2330.
[20]王振兴,朱锦懋,王健,等.闽楠幼树光合特性的初步研究[J].安徽农业大学学报2007,07:3234.
[21]吴兴盛.闽楠的生态特性及栽培技术[J].林业勘察设计,2002,02:6769.
[22]张文辉,祖元刚,刘国彬.十种濒危植物的种群生态学特征及致危因素分析[J].生态学报,2002,22(9):15121520.
(责任编辑:夏剑萍)
猜你喜欢
领导文萃(2020年18期)2020-09-29
老友(2020年7期)2020-08-28
湖南农业科学(2020年2期)2020-05-06
少年博览·小学高年级(2018年9期)2018-11-26
当代党员(2018年1期)2018-01-22
领导文萃(2017年19期)2017-10-26
百家讲坛(2016年18期)2016-11-01
故事会(2016年16期)2016-08-23
百家讲坛(2016年7期)2016-08-09
林业与生态(2014年8期)2014-12-23
