
白菜型冬油菜叶片结构和光合特性对冬前低温的响应
2017-03-16 09:11许耀照曾秀存张芬琴孙佳孙万仓武军艳方彦刘自刚孙柏林
作物学报 2017年3期
许耀照曾秀存张芬琴孙 佳孙万仓武军艳方 彦刘自刚孙柏林
1甘肃农业大学农学院 / 甘肃省油菜工程技术研究中心, 甘肃兰州 730070;2河西学院 / 甘肃省河西走廊特色资源利用重点实验室,甘肃张掖 734000;3Department of Plant Sciences, University of Manitoba, Winnipeg, Manitoba, R3T2N2, Canada
白菜型冬油菜叶片结构和光合特性对冬前低温的响应
许耀照1,2,**曾秀存2,**张芬琴2孙 佳3孙万仓1,*武军艳1方 彦1刘自刚1孙柏林2
1甘肃农业大学农学院 / 甘肃省油菜工程技术研究中心, 甘肃兰州 730070;2河西学院 / 甘肃省河西走廊特色资源利用重点实验室,甘肃张掖 734000;3Department of Plant Sciences, University of Manitoba, Winnipeg, Manitoba, R3T2N2, Canada
为明确白菜型冬油菜在冬前低温下叶片结构特征、光合作用特性及其抗寒性, 本研究在0℃和–7.6℃自然低温条件下, 选用白菜型冬油菜品种陇油7号(超强抗寒)和天油4号(弱抗寒), 测定并比较其叶片气孔性状、解剖结构和光合、荧光参数的日变化等指标。结果表明, 随着冬前温度下降, 2个白菜型冬油菜叶片气孔密度、气孔面积、气孔周长、栅栏组织和海绵组织厚度均变小, 细胞间隙变大, 叶片变薄; Pn日变化呈“单峰”型曲线, 无光合“午休”现象;叶片的Pn、Gs、Tr和CE均降低, 而Ci均升高, 说明是非气孔限制引起Pn降低。白菜型冬油菜在冬前低温条件下发生了光抑制现象, 表现为Fm和Fv/Fm下降, Fo上升。与超强抗寒品种陇油7号相比, 弱抗寒品种天油4号叶片气孔密度和气孔面积均较大, 气孔总周长较长, 叶片较厚, Pn、Fm和Fv/Fm均较高, 说明冬前低温条件下, 天油4号光合能力较强, 光抑制程度较弱。白菜型冬油菜在冬前低温条件下的叶片气孔密度越大、气孔面积越大、气孔周长越长、叶片及栅栏组织和海绵组织越厚, 光合能力越强, 地上部生长越旺盛, 品种抗寒性越差。本研究为冬油菜抗寒种质创新和育种提供了部分理论支撑。
白菜型冬油菜; 叶片结构; 光合特性; 叶绿素荧光参数; 低温; 抗寒性
白菜型冬油菜是我国西北寒旱区主要的油料作物, 其整个生育期经历较长的低温环境。冬前低温驯化阶段光合产物的累积是冬油菜能否安全越冬以及返青后地上部营养体重建的关键[1], 而光合作用是作物对低温最敏感的过程[2], 低温光合响应机制对于农业生产及低温育种具有重要的指导意义。因此, 冬前低温是影响冬油菜安全越冬的重要环境因子[3]。目前关于冬油菜抗寒性研究多集中于外部形态[4]、抗氧化和渗透调节能力[5-6]以及抗性基因的克隆[7-8]等方面, 而较少报道从叶片解剖结构与光合作用相结合揭示冬前持续低温对其抗寒性的影响。
叶片作为植物光合作用的主要器官, 其形态结构的变化是植物对环境适应的基础[9]。已有研究认为, 叶片上下表皮厚度、栅栏组织和海绵组织厚度以及叶片总厚度的变化是植物适应低温环境的反应[10],叶片栅栏组织细胞层数及细胞大小、气孔大小、栅栏组织与海绵组织的厚度比、海绵组织与叶片厚度比等都与植株的抗寒性有一定相关性, 而叶片结构的变化往往又导致光合作用的变化[11]。低温胁迫下,植物光合机构受损, PSII反应中心失活, 引起其光能原初捕捉能力和光能同化率减弱, 光合作用能力下降[12]。樊怀福等[13]认为, 低温条件下, 光合速率(Pn)的下降易导致同化物积累量下降, 从而使植株生长势减弱, 而气孔导度(Gs)的下降使光合底物运输能力降低, 进而影响光合作用的正常进行。植物光合变化的信息可以通过叶绿素荧光动力学参数较为准确地表达[14], 故在抗逆性研究中, 常将叶绿素荧光参数作为植物耐寒性评价的重要生理指标。近年来已在菠菜[15]、甘蓝型油菜[16]、白菜[17]、花椰菜[18]等作物的抗寒性鉴定上得到应用。
本研究分析了 2个抗寒性不同的白菜型冬油菜叶片解剖结构特征、气孔结构等叶片组织结构参数和光合、荧光参数日变化等生理参数对冬前低温的响应与抗寒性的关系, 旨在明确低温条件下冬油菜叶片光合作用的运转和调节适应机制, 为我国西北寒旱区白菜型冬油菜抗寒种质创新、抗寒育种及抗寒评价提供理论依据。
1 材料与方法
1.1 试验材料
白菜型冬油菜品种陇油 7号(超强抗寒)和天油 4号(弱抗寒), 均由甘肃省油菜工程技术研究中心提供。
1.2 试验设计
试验材料于2013年8月中下旬播种于河西学院农业与生物技术学院教学科研示范基地, 试验点位于河西走廊中部张掖市, 试验地平均海拔 1474 m,地处东经 100°26′21′, 北纬 38°57′09′之间, 年平均日照时间 3065.6 h, 年平均气温 7.3℃, 最高气温38.6℃, 最低气温–28.7℃, 年平均降水量为 130.4 mm。试验地环境条件一致, 肥力中等, 前茬为春小麦, 试验小区面积为20 m2(4 m ×5 m), 3次重复, 随机区组排列。
在张掖市白菜型冬油菜的枯叶期为当年的 11月下旬至12月上旬, 此后进入越冬期, 为此本试验在越冬前设置2个自然低温观测点T1和T2, 即于10月20日(T1, 晴天)和11月9日(T2, 晴天)观察白菜型冬油菜叶片气孔性状和显微解剖结构, 测定光合特性和叶绿素荧光参数日变化。试验期间(2013年10月至11月)试验点环境温度呈下降趋势(图1)。根据朱慧霞等[19]方法, 以试验前 3 d和测定当天试验地的最低平均气温为低温条件, 本研究观测点 T1 (10月20日)前3 d平均最低气温为0℃, 观测点T2 (11月9日)为–7.6℃。
1.3 试验方法
1.3.1 冬油菜叶片气孔和解剖结构的观察 随机选 5株健康、生长势一致的冬油菜植株, 摘取由顶端向下数的第4片功能叶, 参照唐道城等[20]的方法,将功能叶表皮制作成临时装片, 在 Olympus数码显微镜 40×物镜下观察和照相, 并利用 Motic Images Advanced图象分析软件测量气孔的纵横径(μm)、气孔面积(μm2)、气孔密度数(mm2)和单位面积气孔总长度(μm)。
气孔面积(μm2)=0.25×3.14×横径长×纵径长
气孔密度(mm2)=单位面积的气孔数/单位面积
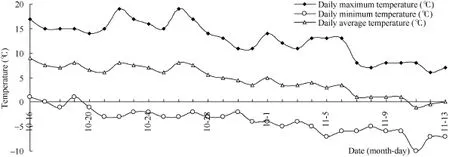
图1 张掖市2013年10月至11月温度Fig. 1 Temperature from October to November in 2013 in Zhangye, Gansu province
单位面积气孔总长度(μm)=气孔总长度/气孔密度
以功能叶的叶脉为中心, 切取1.0 cm × 0.5 cm叶片, 参照李正理[21]的方法制成石蜡切片, 在Olympus光学显微镜下观察, 测量叶片厚度(μm)、表皮厚度(μm)、栅栏组织和海绵组织厚度(μm)。
叶片组织细胞结构疏松度 = 疏松海绵组织厚度/叶片总厚度×100%
叶片组织细胞结构紧密度 = 栅栏组织厚度/叶片总厚度×100%
栅/海比(栅栏组织厚度与海绵组织厚度比值)用P/S表示。
1.3.2 光合特性和叶绿素荧光参数的测定 采用美国PP Systems生产的CIRAS-2型便携式光合测定系统在自然条件下测定 2个白菜型冬油菜功能叶净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)等光合参数的日进程, 参照Fischer等[22]的方法计算瞬时羧化效率(CE=Pn/Ci), 同时记录叶室内光合有效辐射(PAR)、环境CO2浓度(Ca)、环境温度(Ta)和环境相对湿度(RH)等环境参数, 测定从当天10:00开始到18:00结束, 每2 h测量1次标记叶片的光合参数, 重复读取3个观测值。
用英国Hansatech公司生产的FMS-2便携式脉冲调制荧光仪在自然条件下测定2个白菜型冬油菜连体叶片的叶绿素荧光参数, 冬油菜功能叶充分暗适应20 min后, 用弱测量光(0.1 μmol m–2s–1)测定初始荧光(Fo), 随后给一个强闪光(3000 μmol m–2s–1,脉冲时间 0.7 s)测得最大荧光 Fm, 并由该荧光仪读取PSII原初光能转化效率(Fv/Fm), 可变荧光强度Fv= Fm–Fo。测定从当天8:00开始到18:00结束, 每2 h测量1次标记叶片的叶绿素荧光参数。测定光合特性和叶绿素荧光参数时, 选取观测组的每个冬油菜品种5株, 以每株从顶端向下数第4片健康功能叶作为测量叶, 并挂牌标记, 保证测量时叶位无变化,取其平均值作为该种该品时刻的测定值。
1.4 统计分析
用 Microsoft Excel 2003整理数据和作图, 用DPS 9.50软件对数据进行Duncan’s新复极差法差异显著性分析。
2 结果与分析
2.1 冬油菜叶片气孔性状和结构特征
冬油菜叶片气孔是由形状规则、排列紧密的单层细胞组成, 气孔器呈卵圆形, 由 2个呈肾形的保卫细胞对称排列而成(图2)。由表1可以看出, 冬油菜功能叶的气孔密度、气孔面积和单位面积气孔总周长在不同观测温度点以及不同品种间均存在显著差异(P<0.05), 表现为T1>T2, 天油4号>陇油7号。
冬油菜叶肉组织较为发达, 分化为栅栏组织和海绵组织, 栅栏组织由多层细胞构成, 排列紧密规则,约占叶肉组织的 1/2~2/3, 海绵组织细胞较大, 排列疏松, 尤其在T2低温下, 栅栏组织和海绵组织的细胞间隙更大(图3)。由表1可以看出, 冬油菜叶片厚度、栅栏组织和海绵组织厚度均随温度降低而明显变薄(P<0.05), 且在T1和T2观测温度下, 表现为天油4号>陇油7号。叶片疏松度、紧密度和栅/海比在T1和T2观测温度点以及2个品种间无显著差异(P>0.05)。
2.2 环境温度和光合有效辐射日变化
环境温度和光合有效辐射日变化在测定时间内均呈“单峰”曲线, 且T1 (10月20日)的环境温度和光合有效辐射均高于T2 (11月9日)(图4)。T1和T2环境温度从早晨 10:00的 11.8℃和 7.6℃逐渐升高,并于下午 14:00时达到峰值, 分别为 23.9℃和 20.9℃, 而后逐渐下降。

图2 不同品种白菜型冬油菜叶片的气孔分布Fig. 2 Stomatal distribution of leaves in winter rape cultivarsA和C分别为陇油7号在T1和T2温度下的气孔; B和D分别为天油4号在T1和T2温度下的气孔。A and C are stomata of Longyou 7 at T1 and T2 temperatures; B and D are stomata of Tianyou4 at T1 and T2 temperatures.
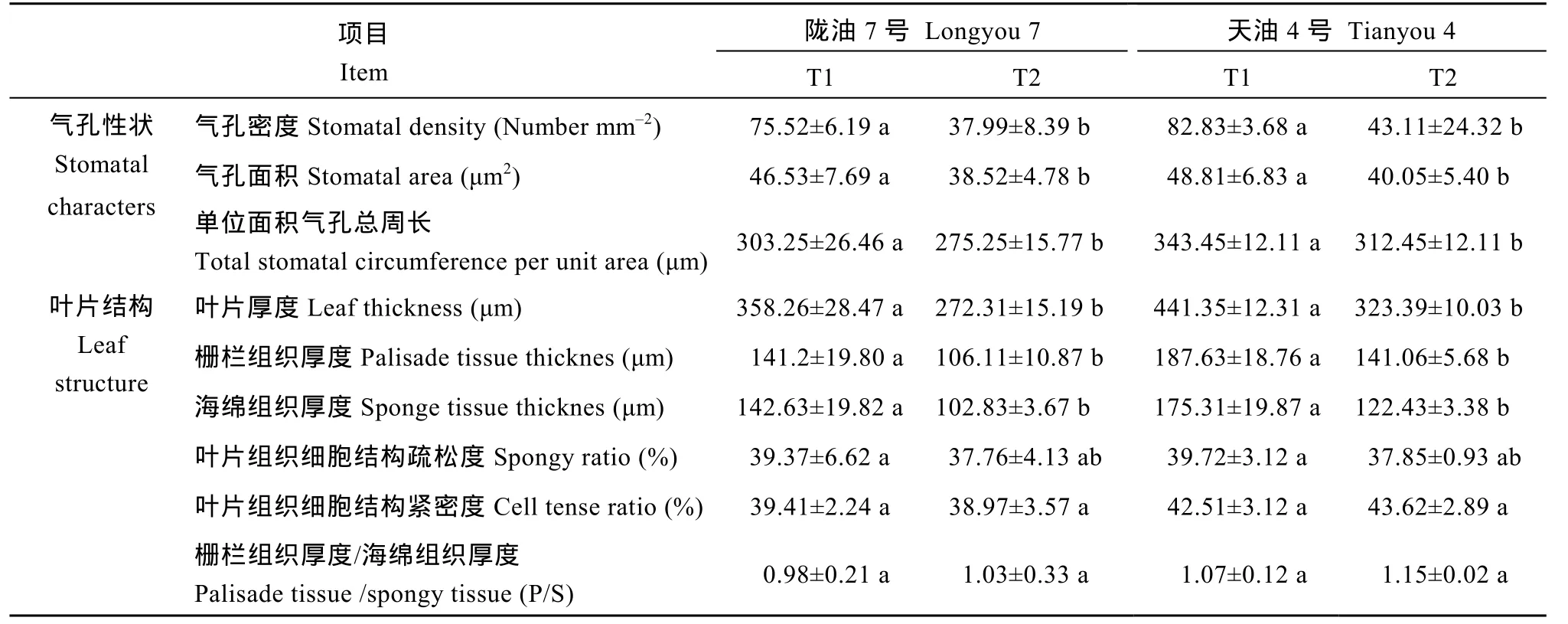
表1 白菜型冬油菜叶片气孔性状及叶片结构的比较Table 1 Comparison of stomatal characters and leaf anatomical structure of winter rape cultivars
光合有效辐射日变化与环境温度的变化趋势一致, 也呈先升高后降低的趋势, 但光合有效辐射明显较环境温度的下降快。在 12:00–14:00时, T1和T2的光合有效辐射最强, 分别为1699.42 μmol m–2s–1和1515.43 μmol m–2s–1, 当18:00时, T1的光合有效辐射降至819.12 μmol m–2s–1, 而T2于16:00时光合有效辐射降至455.14 μmol m–2s–1, 由此表明T1和T2环境温度的变化受光合辐射的影响较大, 环境温度随光合有效辐射增强而升高, 光合有效辐射逐渐减弱, 环境温度逐渐降低。
2.3 冬油菜叶片Pn、Gs、Tr、CE和Ci日变化对冬前低温的响应
白菜型冬油菜的净光合速率(Pn)日变化呈“单峰”型曲线, 无光合“午休”现象(图5)。在10:00时, Pn较低, 而后随光强和温度的增加逐渐升高, 并于14:00时达到峰值。T1观测温度点时, 陇油7号和天

图3 冬前低温条件下白菜型冬油菜叶片的解剖结构Fig. 3 Anatomical structure of winter rape leaves under low temperatureA和C分别为陇油7号在T1和T2温度下叶的解剖结构; B和D分别为天油4号在T1和T2温度下叶的解剖结构。A and C are leaf anatomical structure of Longyou 7 at T1 and T2 temperatures; B and D are leaf anatomical structure of Tianyou 4 at T1 and T2 temperatures.
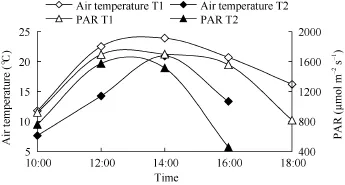
图4 环境温度和光合有效辐射的日变化Fig. 4 Diurnal variation of air temperature and photosynthetic active radiation (PAR)
2.4 冬油菜叶片叶绿素荧光参数Fo、Fm和Fv/Fm日变化对低温的响应
由图7可以看出, 白菜型冬油菜Fo和Fm的日变化呈“V”型曲线, 上午8:00时相对较高, 后随着光强的增加而逐渐下降, T1于12:00时达到最小值,而T2较T1推迟2 h, 于14:00时达到最小值, 此后又回升, 并于 16:00时再次降低, 在同一温度(T1或T2)条件下, 陇油7号的Fo较天油4号大, 而Fm较小。
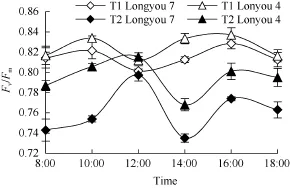
Fv/Fm日变化呈“双峰”曲线(图 8), 并随温度的下降而降低。T1温度下, 陇油 7号和天油 4号的Fv/Fm值于上午10:00和下午 16:00达到峰值, 分别为0.82、0.83和0.83、0.83, 而T2温度下于中午12:00 (较T1推迟2 h)和下午16:00达到峰值, 分别为0.80、0.77和0.82, 0.80。同一温度条件下, 弱抗寒品种天油4号的Fv/Fm值始终高于强抗寒品种陇油7号。
3 讨论
叶片的结构对植物光合作用有明显的影响。已有研究表明, 植物叶片的光合速率取决于叶肉组织、栅栏组织和海绵组织的厚度[23]; 叶片厚度决定了单位叶面积光合细胞的数目, 叶片越厚, 光合细胞数目越多, 光合能力越强, 反之, 光合能力越弱[24-25]。叶片对环境变化比较敏感, 其形态结构的改变是植物对环境的适应性的表现。低温条件下, 植物叶片变薄, 栅栏组织和海绵组织变得疏松, 细胞间隙增大, 细胞间 CO2浓度上升, 光合速率下降, 有向阴生结构变化的趋势[26-27], 这一结果在本研究也得到验证。由此说明冬油菜可通过自身的叶片结构改变而主动地适应冬前低温条件, 也表明冬前低温使其光合作用受到了严重的抑制。

图5 白菜型冬油菜叶片光合速率、气孔导度、蒸腾速率和瞬时羧化效率的日变化Fig. 5 Diurnal variation of Pn, Gs, Tr, and CE in two winter rape leavesA: 净光合速率(Pn)日变化; B: 气孔导度(Gs)日变化; C: 蒸腾速率(Tr)日变化; D: 瞬时羧化效率(CE)日变化。A: diurnal variation of Pn; B: diurnal variation of Gs; C: diurnal variation of Tr; D: diurnal variation of CE.
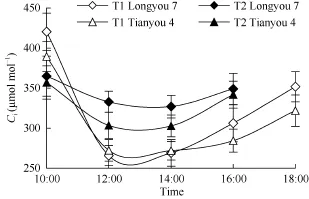
图6 白菜型冬油菜叶片胞间CO2浓度的日变化Fig. 6 Diurnal variation of Ciof two winter rapes leaves
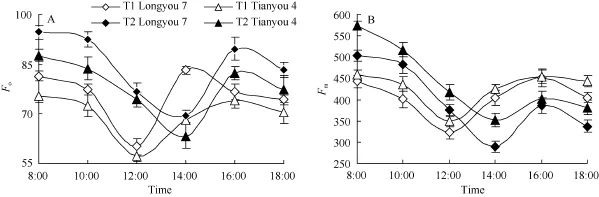
图7 白菜型冬油菜叶片初始荧光和最大荧光的日变化Fig. 7 Diurnal variation of Foand Fmof two winter rapes leavesA: 初始荧光(Fo)日变化; B: 最大荧光(Fm)日变化。A: diurnal variation of Fo; B: diurnal variation of Fm.
气孔在植物适应环境和在光合作用中起着关键作用。低温胁迫下, 许多植物都表现出Pn明显下降。Pn的下降是由气孔限制因素和非气孔因素引起的[28]。本研究中发现, 当温度由0℃下降至–7.6℃时, 2个白菜型冬油菜品种的叶片Pn均下降, 同时Gs、Tr和CE也下降, 而Ci升高, 说明Pn的下降是由非气孔因素引起的, 即因光化学反应和 RuBP羧化反应的限制而阻碍了CO2的利用, 致使Ci积累, 叶肉细胞光合活性下降[29]。植物光合作用日变化曲线分为“单峰”型和“双峰”型2种[30]。白菜型冬油菜净光合速率日变化呈“单峰”型曲线, 无光合“午休”现象, 这与环境温度和光合有效辐射日变化是一致, 说明在一定范围内, 环境温度和光合效速率呈正相关。叶绿素荧光参数最先对植物低温胁迫作出反应, 能够评价其光合系统功能以及环境胁迫的影响。初始荧光(Fo)是判断 PSII反应中心运转情况的重要指标[31-32]。PSII天线的热耗散增加常导致Fo的降低, 而Fo的增加则表明 PSII反应中心受到了破坏或可逆的失活,因此可根据Fo的变化推测反应中心的状况和可能的光保护机制[33]。Fv/Fm和 Fm降低则是判断光抑制的最重要的特征[34]。本研究发现, T2低温下Fo值较T1升高, 而 Fv/Fm和 Fm则下降, 说明冬前低温加剧降低了冬油菜原初光能转化效率, 引起了光抑制, 造成PSII反应中心失活或伤害。冬油菜叶片 Fm的日变化和Fo变化趋势一致, 在中午12:00至14:00时处于较低的水平, 表明中午的强光下冬油菜叶片 PSII受到严重的光抑制, 其吸收的光能中只有极少量用于光化学电子传递和参与 CO2固定, 但下午由于光强的减弱, Fm逐渐恢复。冬油菜Fv/Fm日变化呈“双峰”型曲线, 在中午较高的光强和温度下, 冬油菜光合作用器官的光合活性受到暂时抑制, 但并未受损,后由于光强和温度的下降, 光合器官功能得到恢复, Fv/Fm上升。
已有的研究发现植物抗寒性与叶片气孔密度有密切关系, 气孔密度越小, 抗寒性越强, 反之抗寒性越弱[35-36]。本研究中弱抗寒品种天油 4号气孔密度大于强抗寒品种陇油 7号的结果一致, 因此叶片气孔密度可以作为鉴定冬油菜抗寒性强弱的指标。叶片气孔大小与植物抗寒性的关系, 已有的研究说法不一。吴林等[37]研究认为气孔大小等单一性状与抗寒性无明显的关系, 而李映晖等[38]认为气孔越大的品种抗寒性越强, 但钟克友等[39]研究认为气孔面积和单位叶面积气孔总周长越小水稻耐冷性越强,反之越弱, 这与本研究的结果一致。杨宁宁等[40]研究认为, 抗寒性强的油菜叶片薄、栅栏组织厚度和栅/海小, 这一结果在本研究中也得到证实。前人研究认为植物抗寒性与叶片结构紧密度(CTR值)呈正相关, 与疏松度(SR值)呈负相关[41], 但也有的研究认为SR和CTR与植物抗寒性关系不大[42], 在本研究中2个品种的SR和CTR差异不明显。植物Fv/Fm的正常值为0.83左右, 低于正常值则认为发生了光抑制现象[43]。本研究中2个白菜型冬油菜品种冬前低温下均不同程度地发生光抑制, 且抗寒性强的陇油7号光抑制程度更大, 这与天油4号具有较高的净光合速率的结果是一致的, 同时也与其叶片气孔性状和解剖结构特征相符合。
越冬植物秋冬季温度降低后, 逐渐减弱其生长强度是保证良好锻炼的重要条件[44], 因为冬前植物生长速率过快, 植物组织比较疏松, 呼吸旺盛, 抗寒准备不足, 对逆境适应性差, 抗寒能力弱[45]。本研究中 2个白菜型冬油菜品种叶片气孔性状、解剖结构以及光合特性和叶绿素荧光参数日变化对冬前低温的响应存在差异, 说明白菜型冬油菜在越冬前抗寒性的生理机制存在差异, 而天油 4号叶片气孔性状和结构特征更有利于其在冬前低温下进行光合作用, 致使生长速率过快, 对逆境适应性差, 因此天油4号抗寒性较陇油7号弱, 这与田间观察到抗寒性弱的冬油菜品种冬前地上部生长快, 枯叶期晚的结果是一致的[46]。
4 结论
随着冬前气温降低, 白菜型冬油菜叶片气孔密度、气孔面积、气孔周长、栅栏组织和海绵组织厚度均变小, 细胞间隙变大, 叶片变薄, Pn下降, 并产生光抑制; 白菜型冬油菜Pn日变化呈“单峰”型曲线,无光合“午休”现象, 其 Pn的下降是由非气孔限制引起的; 与超强抗寒品种陇油 7号相比, 冬前低温条件下, 弱抗寒品种天油 4号叶片气孔密度和气孔面积较大, 气孔总周长较长, 叶片、栅栏组织和海绵组织较厚, 光合能力较强, 光抑制程度较轻。
References
[1] 刘自刚, 孙万仓, 杨宁宁, 王月, 何丽, 赵彩霞, 史鹏飞, 杨刚,李学才, 武军艳, 方彦, 曾秀存. 冬前低温胁迫下白菜型冬油菜抗寒性的形态及生理特征. 中国农业科学, 2013, 46: 4679–4687 Liu Z G, Sun W C, Yang N N, Wang Y, He L, Zhao C X, Shi P F, Yang G, Li X C, Wu J Y, Fang Y, Zeng X C. Morphology and physiological characteristics of cultivars with different levels of cold-resistance in winter rapeseed (Brassica campestris L.) during cold acclimation. Sci Agric Sin, 2013, 46: 4679–4687 (in Chinese with English abstract)
[2] 陶宏征, 赵昶灵, 李唯奇. 植物对低温的光合响应. 中国生物化学与分子生物学报, 2012, 28: 501–508 Tao H Z, Zhao C L, Li W Q. Photosynthetic response to low temperature in plant. Chin J Biochem Mol Biol, 2012, 28: 501–508 (in Chinese with English abstract)
[3] 孙万仓, 武军艳, 方彦, 刘秦, 杨仁义, 马维国, 李学才, 张俊杰, 张鹏飞, 雷建明, 孙佳. 北方旱寒区北移冬油菜生长发育特性. 作物学报, 2010, 36: 2124–2134 Sun W C, Wu J Y, Fang Y, Liu Q, Yang R Y, Ma W G, Li X C, Zhang J J, Zhang P F, Lei J M, Sun J. Growth and development characteristics of winter rapeseed northern-extended from the cold and arid regions in China. Acta Agron Sin, 2010, 36: 2124–2134 (in Chinese with English abstract)
[4] 孙万仓, 马卫国, 雷建民, 刘秦, 杨仁义, 吴军艳, 王学芳, 叶剑, 曾军, 张亚宏, 康艳丽, 郭秀娟, 魏文惠, 杨杰, 蒲媛媛,曾潮武, 刘红霞. 冬油菜在西北旱寒区的适应性和北移的可行性研究. 中国农业科学, 2007, 40: 2716–2726 Sun W C, Ma W G, Lei J M, Liu Q, Yang R Y, Wu J Y, Wang X F, Ye J, Zeng J, Zhang Y H, Kang Y L, Guo X J, Wei W H, Yang J, Pu Y Y, Zeng C W, Liu H X. Study on adaptation and introduction possibility of winter rapeseed to dry and cold areas in northwest China. Sci Agric Sin, 2007, 40: 2716–2726 (in Chinese with English abstract)
[5] 蒲媛媛, 孙万仓. 白菜型冬油菜抗寒性与生理生化特性关系.分子植物育种, 2010, 8: 335–339 Pu Y Y, Sun W C. The relationship between cold resistance of winter turnip rape varieties and its physiological characteristics. Mol Plant Breed, 2010, 8: 335–339 (in Chinese with English abstract)
[6] 孔德晶, 王月, 孙万仓, 曾秀存, 方彦, 鲁美红, 杨宁宁. 北方白菜型冬油菜F2主要生理生化特性的变异与抗寒性相关分析.草业学报, 2014, 23(4): 79–86 Kong D J, Wang Y, Sun W C, Zeng X C, Fang Y, Lu M H, Yang N N. Analysis of variation in physio-biochemical characteristics and cod resistance in winter rapeseed F2populations. Acta Pratac Sin, 2014, 23(4): 79–86 (in Chinese with English abstract)
[7] 曾秀存, 孙万仓, 方彦, 刘自刚, 董云, 孙佳, 武军艳, 张鹏飞,史鹏辉, 孔德晶, 张腾国, 何丽, 赵彩霞. 白菜型冬油菜抗坏血酸过氧化物酶(APX)基因的克隆、表达及其活性分析. 作物学报, 2013, 39: 1400–1408 Zeng X C, Sun W C, Fang Y, Liu Z G, Dong Y, Sun J, Wu J Y, Zhang P F, Shi P H, Kong D J, Zhang T G, He L, Zhao C X. Cloning, expression, and activity analysis of ascorbate peroxidase (APX) gene from winter turnip rape (Brassica campestris L.). Acta Agron Sin, 2013, 39: 1400–1408 (in Chinese with English abstract)
[8] 曾秀存, 刘自刚, 史鹏辉, 许耀照, 孙佳, 方彦, 杨刚, 武军艳,孔德晶, 孙万仓. 白菜型冬油菜铜锌超氧化物歧化酶(Cu/Zn-SOD)基因的克隆及其在低温条件下的表达分析. 作物学报, 2014, 40: 636–643 Zeng X C, Liu Z G, Shi P H, Xu Y Z, Sun J, Fang Y, Yang G, Wu J Y, Kong D J, Sun W C. Cloning and expression analysis of copper and zinc superoxide dismutase (Cu/Zn-SOD) gene from Brassica campestris L. Acta Agron Sin, 2014, 40: 636–643 (in Chinese with English abstract)
[9] 邱佳妹, 王康才, 段云晶, 管晖, 魏慧玲. 麦冬、山麦冬叶片解剖结构与光合特性研究. 西北植物学报, 2014, 34: 727–732 Qiu J M, Wang K C, Duan Y J, Guan H, Wei H L. Study on leaf morphology and photosynthetic characteristics of Ophiogon japonicus and Liriope spicata. Acta Bot Boreali-Occident Sin, 2014, 34: 727–732 (in Chinese with English abstract)
[10] 蔡永立, 宋永昌. 浙江天童常绿阔叶林藤本植物的适应生态学: I. 叶片解剖特征的比较. 植物生态学报, 2001, 25(1): 90–98 Cai Y L, Song Y C. Adaptive ecology of lianas in tiantong evergreen broad-leaved forest, Zhejiang, China: I. leaf anatomical characters. Acta Phytoecol Sin, 2001, 25(1): 90–98 (in Chinese with English abstract)
[11] Kloeppe J W, Ryu C M, Zhang S. Induced systemic resistance and promotion of plant growth by Bacillus sp. Phytopathology, 2004, 94: 1259–1266
[12] 刘辉, 郭延平, 胡美君. 杨梅光合作用的低温光抑制. 热带亚热带植物学报, 2005, 13: 338–342 Liu H, Guo Y P, Hu M J. Chilling-induced photoinhibition of photosynthesis in Myrica rubra leaves. J Trop Subtrop Bot, 2005, 13: 338–342 (in Chinese with English abstract)
[13] 樊怀福, 杜长霞, 朱祝军. 外源NO对低温胁迫下黄瓜幼苗生长、叶片膜脂过氧化和光合作用的影响. 浙江农业学报, 2011, 23: 538–542 Fan H F, Du C X, Zhu Z J. Effects of exogenous nitric oxide on plant growth membrane lipid peroxidation and photosynthesis in cucumber seedling leaves under low temperature. Acta Agric Zhejiangensis, 2011, 23: 538–542 (in Chinese with English abstract)
[14] Horton P. Non-photochemical quenching of chlorophyll fluorescence. In: Jennigs R C, ed. Light as an Energy Source and Information Carrier in Plant Physiology. New York: Plenum Press, 1996. pp 99–112
[15] Peterson R B, Sivak M N, Walker D A. Relationship between steady-state fluorescence yield and photosynthetic efficiency in spinach leaf tissue. Plant Physiol, 1998, 88: 158–163
[16] 胡胜武, 于澄宇, 王绥璋, 李红兵, 黄继英. 甘蓝型油菜抗寒性的鉴定及相关性状的研究. 中国油料作物学报, 1999, 21: 33–35 Hu S W, Yu C Y, Wang S Z, Li H B, Huang J Y. Determination of winter hardiness and its correlated characters in Brassica napus. Chin J Oil Crop Sci, 1999, 21: 33–35 (in Chinese with English abstract)
[17] 胡春梅, 侯喜林, 王敏. 低温胁迫对不结球白菜光合及叶绿素荧光特性的影响. 西北植物学报, 2008, 28: 2478–2484Hu C M, Hou X L, Wang M. Effects of low temperature on photosynthetic and fluorescent parameters of non-heading Chinese cabbage. Acta Bot Boreali-Occident Sin, 2008, 28: 2478–2484
[18] 李光庆, 谢祝捷, 姚雪琴, 陈学好. 花椰菜叶绿素荧光参数与耐寒性的关系研究. 园艺学报, 2010, 37: 2001–2006 Li G Q, Xie Z J, Yao X Q, Chen X H. Studies on the relationship between chlorophyll fluorescence parameters and cold tolerance of cauliflower. Acta Hort Sin, 2010, 37: 2001–2006 (in Chinese with English abstract)
[19] 朱惠霞, 孙万仓, 邓斌, 燕妮, 武军燕, 范惠玲, 叶剑, 曾军,刘雅丽, 张亚宏. 白菜型冬油菜品种的抗寒性及其生理生化特性. 西北农业学报, 2007, 16(4): 34–38 Zhu H X, Sun W C, Deng B, Yan N, Wu J Y, Fan H L, Ye J, Zeng J, Liu Y L, Zhang Y H. Study on cold hardiness and its physiological and biochemical characteristics of winter turnip rape (Brassica campetris). Acta Agric Boreali-Occident Sin, 2007, 16(4): 34–38 (in Chinese with English abstract)
[20] 唐道城, 李宗仁, 王艳萍. 白芥及三大类型油菜叶片的气孔特征比较. 青海大学学报(自然科学版), 1999, 17(1): 1–4 Tang D C, Li Z R, Wang Y P. Comparison about stomatal characteristic among leaf types on white mustaral (Sinapis alba) and three types of rope. J Qinghai Univ, 1999, 17(1): 1–4 (in Chinese with English abstract)
[21] 李正理. 植物制片技术(第2版). 北京: 科学出版社, 1987. pp 129–137 Li Z L. Plant Production Technology, 2nd edn. Beijing: Science Press, 1987. pp 129–137 (in Chinese)
[22] Fiseher R A, Tumer N C. Plant produetivity in the arid and semi-arid zones. Annu Rev Plant Physiol, 1978, 29: 277–317
[23] 付雅丽. 大白菜随体单体、三体及其二倍体的光合作用研究.河北农业大学硕士学位论文, 河北保定, 2004. pp 1–2 Fu Y L. Studies on the Photosynthesis of Sat-monosome, Sat-trisome and Its Diploid in Chinese Cabbage. MS Thesis of Hebei Agricultural University, Baoding, China, 2004. pp 1–2 (in Chinese with English abstract)
[24] Araus J L, Alegre L, Tapia L, Calafell R, Serret M D. Relationships between photosynthetic capacity and leaf structure in several shade plants. Am J Bot, 1986, 73: 1760–1770
[25] Gamier E, Salager J L, Laurent G, Sonie L. Relationgships between photosynthesis, nitrogen and leaf structure in 14 grass species and their dependence on the basis of expression. New Phytol, 1999, 143: 119–129
[26] 曹红星, 黄汉驹, 雷新涛, 张大鹏, 张如莲. 低温胁迫下椰子叶片解剖结构差异研究. 热带作物学报, 2014, 35: 2420–2425 Cao H X, Huang H J, Lei X T, Zhang D P, Zhang R L. Difference of the leaf anatomical structure of coconut varieties under low temperature treatments. Chin J Trop Crops, 2014, 35: 2420–2425 (in Chinese with English abstract)
[27] 李章田, 段承俐, 萧凤回. 人工光照对一年生三七形态和光合特性的影响. 云南农业大学学报, 2009, 24: 677–683 Li Z T, Duan C L, Xiao F H. Effects of artificial light treatments on morphological structure and photosynthetic indices of one-year-old panax notoginseng plants. J Yunnan Agric Univ, 2009, 24: 677–683 (in Chinese with English abstract)
[28] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annu Rev Plant Physiol, 1982, 33: 317–345
[29] 眭晓蕾, 毛胜利, 王立浩, 李伟, 张宝玺, 张振贤. 低温对弱光影响甜椒光合作用的胁迫效应. 核农学报, 2008, 22: 880–886 Sui X L, Mao S L, Wang L H, Li W, Zhang B X, Zhang Z X. Effects of low temperature on photosynthesis of sweet pepper under low light. J Nucl Agric Sci, 2008, 22: 880–886 (in Chinese with English abstract)
[30] 杨甲定, 刘志民. 引种于青藏高原的大田玉米研究: 光合作用日变化的特点. 作物学报, 2002, 28: 475–479 Yang J D, Liu Z M. Study oil field-grown maize introduced into Tibetan plateau L: some characteristics of diurnal variation of photosynthesis. Acta Agron Sin, 2002, 28: 475–479
[31] Genty B, Brintais J M, Baker N R. The relationship between the quantum yield of photosyntetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta, 1989, 990: 87–92
[32] 缴丽莉, 路丙社, 周如久, 白志英, 梁海永, 甄红伟. 遮光对青榨槭光合速率及叶绿素荧光参数的影响. 园艺学报, 2007, 34: 173–178 Jiao L L, Lu B S, Zhou R J, Bai Z Y, Liang H Y, Zhen H W. Effects of shading on net photosynthetic rate and chlorophyll fluorescence parameters of leaf in David maple (Acer davidii Franch.). Acta Hort Sin, 2007, 34: 173–178 (in Chinese with English abstract)
[33] 吕芳德, 徐德聪, 侯红波, 刘云龙, 郑良康. 5种红山茶叶绿素荧光特性的比较研究. 经济林研究, 2003, 21(4): 4–7 Lyu F D, Xu D C, Hou H B, Liu Y L, Zheng L K. Comparative study on chlorophyll fluorescence character of five kinds of camellia. Econ For Res, 2003, 21(4): 4–7 (in Chinese with English abstract)
[34] Razinger J, Dermastia M, Drinovec L, Drobne D, Zrimec A, Koce J D. Antioxidative responses of duckweed (Lemna minor L.) to short-term copper exposure. Environ Sci Poll Res Internat, 2007, 14: 194–201
[35] 李国华, 徐涛, 陈国云, 岳海, 梁国平. 10个品种澳洲坚果叶片解剖学的比较研究. 热带作物学报, 2009, 30: 1437–1441 Li G H, Xu T, Chen G Y, Yue H, Liang G P. Anatomical structure of leaves of 10 macadamia cultivars. Chin J Trop Crops, 2009, 30: 1437–1441 (in Chinese with English abstract)
[36] 韦晓霞, 吴如健, 胡菡青. 橄榄种质资源叶片解剖结构的研究.福建农业学报, 2006, 21: 370–374 Wei X X, Wu R J, Hu H Q. Study on natomical leaf structure of Chinese olive germplasm. Fujian J Agric Sci, 2006, 21: 370–374 (in Chinese with English abstract)
[37] 吴林, 刘海广, 刘雅娟. 越橘叶片组织结构及其与抗寒性的关系. 吉林农业大学学报, 2005, 27(1): 48–50 Wu L, Liu H G, Liu Y J. Studies on leaf tissue structure and its relations to cold resistance of blueberry. J Jilin Agric Univ, 2005, 27(1): 48–54
[38] 李映晖, 李润唐, 吴钿, 邹雪娟, 叶昌辉. 3个菠萝品种叶片气孔特征及其与抗寒性的关系, 安徽农业科学, 2013, 41: 1–3 Li Y Y, Li R T, Wu D, Zou X J, Ye C H. Studies on relationship between stomata and cold resistance of three pineapple cultivars. J Anhui Agric Sci, 2013, 41: 1–3 (in Chinese with English abstract)
[39] 钟克友, 唐湘如, 胡飞, 黎国喜, 肖立中, 段美洋. 水稻剑叶气孔性状与孕穗期耐冷性的关系研究. 西北植物学报, 2010,30(1): 98–103 Zhong K Y, Tang X R, Hu F, Li G X, Xiao L Z, Duan M Y. Correlations between the cold tolerance during the booting stage and the stomatal traits of flag leaf surface in rice. Acta Bot Boreali-Occident Sin, 2010, 30(1): 98–103 (in Chinese with English abstract)
[40] 杨宁宁, 孙万仓, 刘自刚, 史鹏辉, 方彦, 武军艳, 曾秀存, 孔德晶, 鲁美宏, 王月. 北方冬油菜抗寒性的形态与生理机制.中国农业科学, 2014, 47: 452–461 Yang N N, Sun W C, Liu Z G, Shi P H, Fang Y, Wu J Y, Zeng X C, Kong D J, Lu M H, Wang Y. Morphological characters and physiological mechanisms of cold resistance of winter rapeseed in northern China. Sci Agric Sin, 2014, 47: 452–461 (in Chinese with English abstract)
[41] 彭伟秀, 杨建民, 张芹, 蒋品. 不同抗寒性的杏品种叶片组织结构比较. 河北林果研究, 2001, 16(2): 145–147 Peng W X, Yang J M, Zhang Q, Jiang P. A comparison among apricot varieties with different cold hardiness in leaf tissue structure. Hebei J For Orchard Res, 2001, 16(2): 145–147 (in Chinese with English abstract)
[42] 梁李宏, 梅新, 林锋, 夏军, 刘术金, 王金辉. 低温胁迫对腰果幼苗叶片组织结构和生理指标的影响. 生态环境学报, 2009, 18: 317–32 Liang L H, Mei X, Lin F, Xia J, Liu S J, Wang J H. Effect of low temperature stress on tissue structure and physiological index of cashew young leaves. Ecol Environ Sci, 2009, 18: 317–321 (in Chinese with English abstract)
[43] Johnson G N, Young A J, Scholes J D, Horton P. The dissipation of excess excitation energy in British plant species. Plant Cell Environ, 1993, 16: 673–679
[44] 张爱民, 孙其信. 有关小麦抗寒生理的研究概况. 国外农学——麦类作物, 1985, (6): 32–35 Zhang A M, Sun Q X. The research situation of wheat cold hardiness physiology. Foreign Agric—Triticeae Crops, 1985, (6): 32–35 (in Chinese with English abstract)
[45] 潘瑞炽. 植物生理学(第6版). 北京: 高等教育出版社, 2008. p 289 Pan R C. Plant Physiology, 6th edn. Beijing: Higher Education Press, 2008. p 289 (in Chinese )
[46] 孙万仓, 武军艳, 曾军, 朱慧霞, 刘雅丽, 张亚宏. 8个白菜型冬油菜品种抗寒性的初步评价. 湖南农业大学学报(自然科学版), 2007, 33(8): 151–155 Sun W C, Wu J Y, Zeng J, Zhu H X, Liu Y L, Zhang Y H. Primary evaluation of cold tolerance among eight winter Brassica rapa. J Hunan Agric Univ (Nat Sci Edn), 2007, 33(8): 151–155 (in Chinese with English abstract)
Response of Leaf Anatomical Structure and Photosynthetic Characteristics of Winter Turnip Rape (Brassica rapa L.) to Low Temperature before Winter
XU Yao-Zhao1,2,**, ZENG Xiu-Cun1,2,**, ZHANG Fen-Qin2, SUN Jia3, SUN Wan-Cang1,*, WU Jun-Yan1, FANG Yan1, LIU Zi-Gang1, and SUN Bo-Lin2
1College of Agronomy, Gansu Agricultural University / Gansu Province Rapeseed Engineering Research Center, Lanzhou 730070 China;2Hexi University / Key Laboratory of Hexi Corridor Resources Utilization of Gansu, Zhangye 734000, China;3Department of Plant Sciences, University of Manitoba, Winnipeg, Manitoba, R3T2N2, Canada
Stomatal traits, leaf anatomic structure characteristic and diurnal variation of photosynthesis and fluorescence parameters, in two winter turnip rape (B. rapa L.) cultivars Longyou 7 (strong cold-tolerant) and Tianyou 4 (weak cold-tolerant) were determined under 0°C and –7.6°C natural low temperature conditions to clearify the effects of low temperature on leaf anatomical structure, mechanism of photosynthesis and cold resistance. With decreasing the temperature before winter, stomatal density, stomatal area, and total stomatal circumference per unit area in two cultivars decreased. Moreover, leaf thickness, palisade tissue and sponge tissue thickness became thinner and cellular space grew bigger. The diurnal variation of net photosynthetic rate of two cultivars presented a single-peak curve, and no typical mid-day depression occurred. Under the lower temperature, Pndeclined with a non-stomatal limitation in two cultivars. Photo inhibition in the two cultivars occurred under low temperature and increased with the temperature decreased. Compared with Longyou 7, Tianyou 4 had bigger stomatal density, stomatal area, total stomatalcircumference per unit area and leaf thickness, as well as higher photosynthetic capacity and weaker photo inhibition. In conclusion, higher photosynthesis and faster growth in winter turnip rape under lower temperature may result from its bigger stomatal density, stomatal area and total stomatal circumference per unit area as well as thicker leaf, palisade tissue and sponge tissue, which results in a poorer resistance to cold stress. The result may be useful for mining new germplasm and facilitating winter turnip rape breeding for cold resistance.
Winter turnip rape (Brassica rapa L.); Leaf anatomical structure; Photosynthesis characteristic; Fluorescence parameters; Low temperature; Cold resistance
10.3724/SP.J.1006.2017.00432
本研究由国家自然科学基金项目(31560397, 31460356, 31660401, 31560072), 甘肃省自然科学基金项目(145RJZG050, 1506RJZG051)和甘肃省河西走廊特色资源利用重点实验室面上项目(XZ1403)资助。
This study was supported by the National Natural Science Foundation of China (31560397, 31460356, 31660401, 31560072), the Gansu Provincial Natural Science Foundation (145RJZG050, 1506RJZG051), and Gansu Province Key Laboratory Program of Hexi Corridor Resources Utilization (XZ1403).
*通讯作者(Corresponding author): 孙万仓, E-mail: 18293121851@163.com
联系方式: E-mail: xuyaozhao@126.com**同等贡献(Contributed equally to this work)
稿日期): 2016-06-13; Accepted(接受日期): 2016-11-02; Published online(
日期): 2016-11-15.
URL: http://www.cnki.net/kcms/detail/11.1809.S.20161115.1618.008.html
猜你喜欢
今日农业(2022年1期)2022-11-16
蔬菜(2022年10期)2022-11-10
电脑迷(2022年10期)2022-11-08
农业工程学报(2022年5期)2022-06-22
热带作物学报(2022年5期)2022-06-01
今日农业(2021年20期)2021-11-26
农技服务(2021年6期)2021-09-22
河北果树(2020年4期)2020-11-26
河北果树(2020年1期)2020-02-09
中国果菜(2016年9期)2016-03-01
