
栽培措施对酿酒葡萄赤霞珠和霞多丽果皮脂肪酸组分的影响
2017-03-03 03:29鞠延仑赵现方房玉林
食品科学 2017年3期
鞠延仑,刘 敏,赵现方,曾 婕,闵 卓,房玉林,2,*
(1.西北农林科技大学葡萄酒学院,陕西 杨陵 712100;2.陕西省葡萄与葡萄酒工程技术研究中心,陕西 杨陵 712100)
栽培措施对酿酒葡萄赤霞珠和霞多丽果皮脂肪酸组分的影响
鞠延仑1,刘 敏1,赵现方1,曾 婕1,闵 卓1,房玉林1,2,*
(1.西北农林科技大学葡萄酒学院,陕西 杨陵 712100;2.陕西省葡萄与葡萄酒工程技术研究中心,陕西 杨陵 712100)
为研究栽培措施对酿酒葡萄脂肪酸组分的影响,以欧亚种(Vitis vinifera L.)酿酒葡萄品种赤霞珠(Cabernet Sauvignon)、霞多丽(Chardonnay)为实验材料,通过植物生长调节剂、摘叶、机械损伤、套袋、不同采收时间等方式进行处理分析。结果表明:葡萄果皮不饱和脂肪酸主要由亚油酸、油酸、棕榈油酸组成;亚麻酸未检测出;饱和脂肪酸主要由棕榈酸、硬脂酸、山嵛酸、花生酸组成。脱落酸、茉莉酸甲酯、光照强度、采收时间、机械损伤处理能够影响亚油酸的含量,并且这种影响与品种有关。在两个品种葡萄果皮中,都发现了呈显著正相关的脂肪酸组分,但品种间差异较大;在霞多丽葡萄果皮中,呈显著正相关的脂肪酸组分明显多于赤霞珠;并且,饱和脂肪酸之间的相关性强于不饱和脂肪酸之间的相关性。结果将为改善葡萄果皮脂肪酸组分,进而提高果实品质提供理论依据。
栽培措施;酿酒葡萄;脂肪酸组分;相关性
葡萄作为世界种植范围最广的果树之一,具有重要的经济价值和营养价值。葡萄果实品质的形成受温度、光照、水分等因素的影响,而葡萄原料的品质又直接关系到葡萄酒品质。葡萄园的栽培措施严重影响了葡萄原料的品质,进而对葡萄产业产生很大影响。酿酒葡萄原料的品质取决于其本身的香气品质,绿叶挥发性物质(green leaf volatiles,GLVs)是葡萄果实香气的重要来源,脂氧合酶(lipoxygenase,LOX)是GLVs形成过程中的关键酶[1-5]。LOX能专一地催化含有顺、顺-1,4戊二烯结构的多不饱和脂肪酸及其相应酯类物质的加氧反应,生成具有共轭双键的脂氢过氧化物[4,6],再经过不同酶的作用发生一系列反应,产物直接参与果实香气的形成,决定着果实品质[7-8]。
脂肪酸组分是GLVs形成的直接底物,是形成葡萄香气的主要前体物质,LOX催化脂肪酸得到的产物氢过氧化物再被氢过氧化物裂解酶催化后得到的挥发性的小分子醇、醛、酯类物质[4,9-12],这些小分子是葡萄浆果、葡萄汁和葡萄酒香气的主要来源[13]。
脱落酸(abscisic acid,ABA)对植物的生长发育,对抗环境胁迫等具有积极的调节作用[14],外源ABA处理能引起植物多种代谢活动的改变,从多方面对植物产生影响[15],研究发现ABA对促进果实成熟、提高果实品质有重要作用[16]。茉莉酸与ABA在植物体内有相似的生理作用,两者的合成均受到LOX的调控。研究表明,植物受到损伤后,ABA、茉莉酸含量在短时间内明显升高,说明ABA、茉莉酸在逆境胁迫适应性反应的上游起作用,而ABA、茉莉酸积累量明显受到LOX活性的影响[17]。研究两者对植株生长发育及果实品质的影响具有重要意义。
目前,栽培措施对葡萄品质影响研究主要集中在鲜食葡萄上,对酿酒葡萄品质的影响鲜有报道。尤其是果皮脂肪酸组分影响方面的研究较少,本课题组研究了植物生长调节剂、光照、采收期、机械损伤等对葡萄果皮香气物质形成的前体脂肪酸组分的影响,旨在考察栽培措施对酿酒葡萄GLV代谢途径中直接底物脂肪酸组分的影响,为提高酿酒葡萄品质、葡萄园生产管理提供理论依据。
1 材料与方法
1.1 材料与试剂
1.1.1 材料
实验于陕西省杨陵农业示范区曹辛庄葡萄园进行,实验用葡萄于2008年扦插定植,欧亚种赤霞珠和霞多丽,均采用单杆双臂架型,南北行,株行距0.8 m×1.5 m,常规管理。
1.1.2 试剂
茉莉酸甲酯、脱落酸 美国Sigma-Aldrich公司;十七烷酸甲酯 美国AccuStandard公司;NaOH(色谱级,下同)、聚乙二醇辛基苯基醚、吐温20、交联聚乙烯吡咯烷酮、石油醚、乙醚、甲醇、甲酸 天津科密欧化学试剂有限公司。
1.2 仪器与设备
centrifuge 5424R离心机 德国Eppendorf公司;Mili-Q超纯水系统 上海摩速科学器材有限公司;101-2AB型烘箱 上海百典仪器厂;FA2004型电子天平、RE-52C旋转蒸发仪 上海精科实业有限公司;TGL-16M高速台式冷冻离心机 上海卢湘仪离心机仪器有限公司;热电TRACE DSQ气相色谱-质谱联用仪 美国Thermo Finnigan公司。
1.3 方法
1.3.1 外源激素处理
ABA处理:配制质量浓度为200、400、600、800、1 000 mg/L的ABA溶液,在果实转色初期,于晴朗无风的傍晚,分别喷洒在长势中庸一致的葡萄植株的葡萄果粒表面,直到液体不成滴流下,每个质量浓度为一个处理,每个处理重复在5 株葡萄植株上进行,分别记为A1~A5。对照组为蒸馏水处理。
茉莉酸甲酯(methyl jasmonate,MeJA)处理:配制浓度为50、100、200、400、800 μmol/L的MeJA溶液,在果实转色初期,于晴朗无风的傍晚,分别喷洒在长势中庸一致的葡萄植株的葡萄果粒表面,直到液体不成滴流下,每个处理重复在5 株葡萄植株上进行,分别记为J1~J5。对照组为蒸馏水处理。
1.3.2 果实受光处理
增加光照的措施为摘除老叶,在果实转色初期,将新梢基部起第1~5片叶片摘除,使75%的果穗表面暴露在阳光下,处理重复在5 株葡萄植株上进行。减少光照的措施为套袋,采用专用葡萄果袋,在葡萄转色初期进行套袋,处理重复在5 株葡萄植株上进行。对照组为正常光照处理。
1.3.3 采收时间
提早采收的采收时间比正常采收的时间提早一周,延迟采收的采收时间比正常采收的时间延迟一周,处理重复在5 株葡萄植株上进行。对照组为正常时间采收。
1.3.4 机械损伤处理
葡萄转色初期,在葡萄植株上对浆果进行创伤处理,使用6 枚×6 枚的大头针创伤板对整穗葡萄实施创伤处理,每粒葡萄果实平均刺入1~2 次,该处理重复在5 株葡萄植株上进行。对照组为无机械损伤处理。
1.3.5 样品采集与处理
经过上述方法处理后,对葡萄植株进行正常管理,待浆果成熟时(霞多丽的采收时间为2013年8月12日,赤霞珠的采收时间为2013年9月12日,提早采收和延迟采收处理的样品除外)随机取无病健康、大小均一果穗10 穗,样品采下后立即放入冰盒中,1 h内带回陕西省葡萄与葡萄酒工程技术研究中心实验室,用不锈钢镊子小心剥下葡萄皮,将葡萄皮内部附着的果肉清除干净,以蒸馏水冲洗、沥干后,置于10 mL离心管内,液氮速冻后保存在-80 ℃冰箱内。
1.3.6 脂肪酸的提取和测定
脂肪酸的提取:参考文献[18-19]的研究方法,略微改动。取葡萄皮5 g在液氮下充分研磨,加入10 mL石油醚-乙醚(4∶3,V/V)混合液于0~4 ℃条件下提取24 h。然后加入0.4 mol/L 氢氧化钾-甲醇溶液10 mL,在室温条件下反应2 h,室温4 000 r/min离心10 min,取上层有机相溶液,置于10 mL蒸馏烧瓶中,减压蒸馏,定容至5 mL。取1 mL样品溶液于进样瓶中,并加入质量浓度为100 mg/mL 1 μL的十七烷酸甲酯作为内标物,待用。
色谱条件:参考文献[20-21]。DB5MS石英毛细管柱(30 m×0.25 mm,0.25 μm);进样1 μL;分流进样(分流比20∶l);进样口温度为240 ℃;检测器温度为250 ℃;升温程序;100 ℃保持1 min,以8 ℃/min的升温速率升至205 ℃,保留0.l min,再以3 ℃/min的升温速率升至240 ℃,保留10 min;载气(N2)压力为0.60 kg/cm2,燃气(H2)压力为0.65 kg/cm2;助燃气(空气)压力为0.50 kg/cm2。
质谱条件:电子轰击离子源;电子能量为70 eV;激活电压为1.5 V;传输线温度为275 ℃;离子源温度为250 ℃;质量扫描范围m/z为40~500。
脂肪酸的定性方法参考文献[22],样品质谱图与标准谱图对比,相似度≥95%时确认为该种化合物。采用内标法计算每种脂肪酸含量。
1.4 数据分析
2 结果与分析
2.1 酿酒葡萄果皮中脂肪酸组分的测定
由表1~3可知,在酿酒葡萄果皮中共检测出16 种脂肪酸,葡萄果皮中不饱和脂肪酸主要由亚油酸、油酸、棕榈油酸组成,以亚油酸含量最多,亚麻酸未检测出,且多不饱和脂肪酸总量占脂肪酸总量的70%以上;主要的4 种饱和脂肪酸为(含量由高到低):棕榈酸、硬脂酸、山嵛酸、花生酸,且棕榈酸含量远远高于其他3 种酸,且霞多丽葡萄果皮中的脂肪酸含量多于赤霞珠(表2、3)。不同处理对两个葡萄品种脂肪酸组分含量的影响也不同,激素处理会降低霞多丽亚油酸和油酸含量,但提高棕榈油酸含量。低质量浓度的激素处理会提高赤霞珠亚油酸含量,高质量浓度激素处理降低赤霞珠亚油酸含量,且不同栽培措施处理会提高赤霞珠油酸和棕榈油酸含量。不同栽培措施处理会同时提高霞多丽和赤霞珠果皮内不饱和脂肪酸的含量。栽培措施对不同品种葡萄脂肪酸组分含量的不同影响的具体机理尚不明确,还需进一步研究。由于在检测出的脂肪酸中亚油酸含量最多,且亚油酸是果皮香气物质的的主要前体之一,所以主要研究不同处理对葡萄亚油酸含量的影响,进一步揭示各栽培措施对葡萄脂肪酸组分的影响。
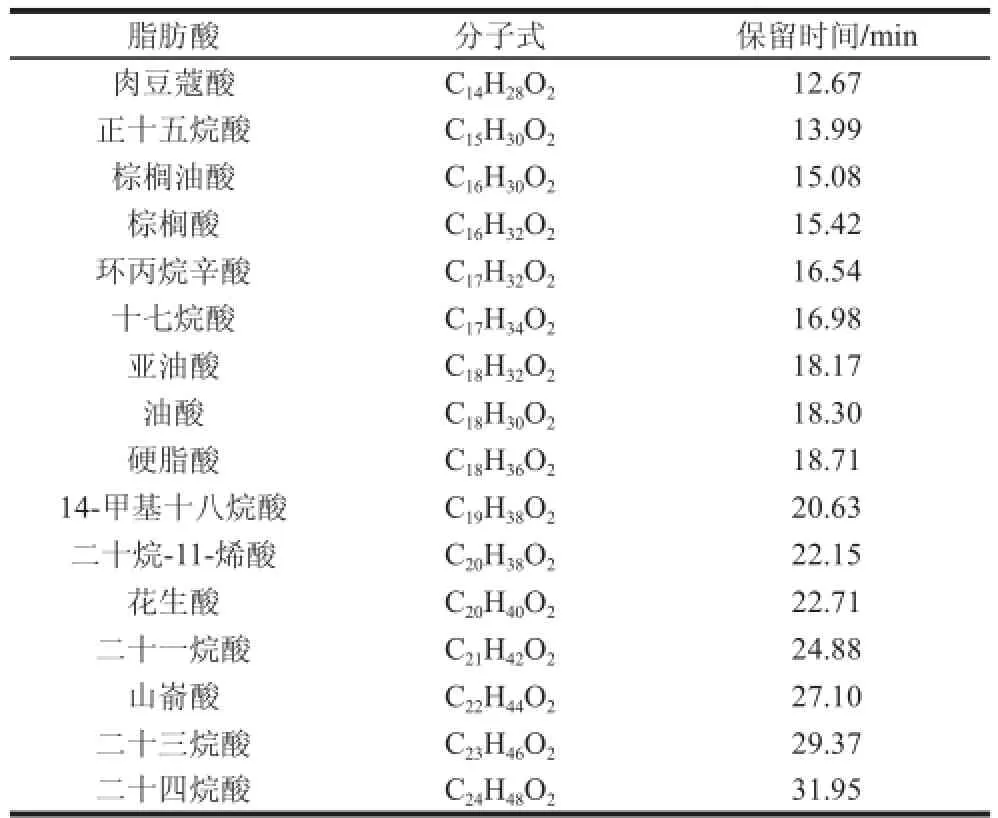
表1 葡萄果皮中检测出的脂肪酸组分Table1 Fatty acid composition of grape berry skins and their retention tiimmeess
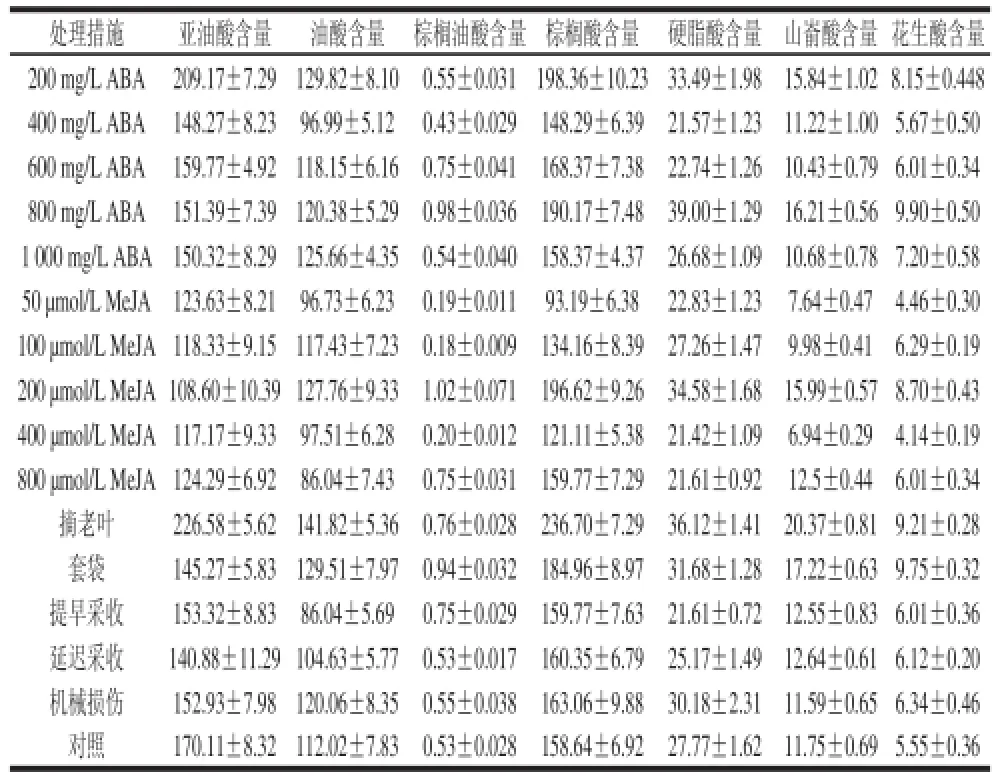
表2 栽培措施对霞多丽葡萄果皮中脂肪酸组分含量的影响Table2 Effects of various experimental treatments on fatty acids in skins of Chardonnay berries skins mg/kg
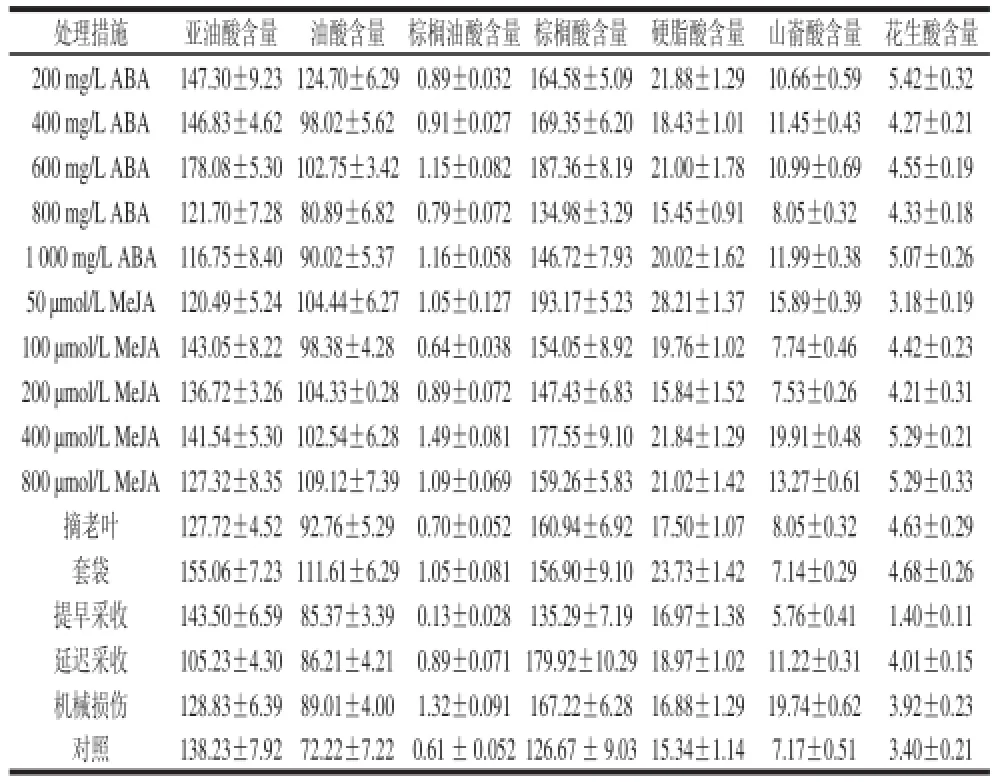
表3 不同处理对赤霞珠葡萄果皮中脂肪酸组分含量的影响Table3 Effects of various experimental treatments on fatty acids in skins of Cabernet Sauvignon berries mg/kg
2.2 ABA处理对葡萄果皮中亚油酸含量的影响
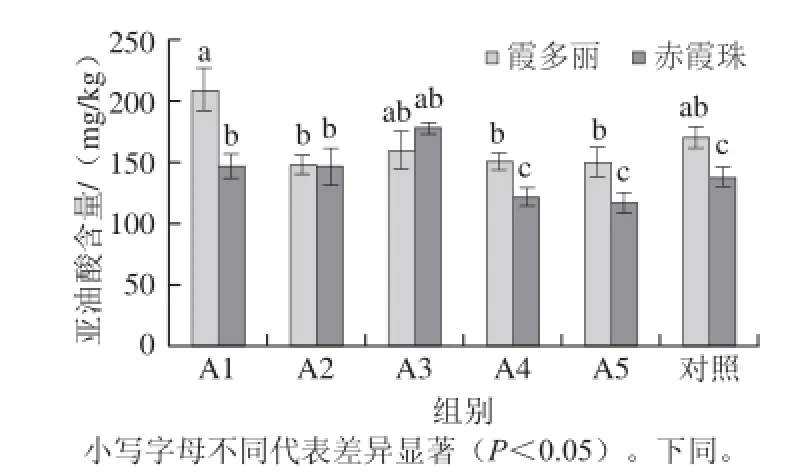
图1 ABA处理对果皮中亚油酸含量的影响Fig.1 Effects of ABA application on linoleic acid content in grape berry skins
ABA最初被当做一种生长抑制剂,后来,人们逐渐认识到其在胚胎发育、果实成熟、逆境胁迫等方面具有积极作用[9]。由图1可知,ABA对不同葡萄品种果皮中亚油酸含量的影响不同。在低质量浓度范围内(≤200 mg/L)ABA处理显著增加霞多丽葡萄果皮中亚油酸含量,但在此质量浓度范围内ABA处理对赤霞珠葡萄果皮亚油酸含量显著高于对照组;在高质量浓度(≥400 mg/L)范围内,霞多丽果皮中亚油酸含量显著低于对照组,而当ABA处理质量浓度为600 mg/L时,赤霞珠葡萄果皮中亚油酸含量达到177.18 mg/kg,显著高于对照组。
2.3 茉莉酸处理对葡萄果皮中亚油酸含量的影响
GLV途径和茉莉酸类物质的合成途径既相互竞争,又相互促进。多不饱和脂肪酸在LOX的催化作用下形成氢过氧化物,氢过氧化物可通过丙二烯氧化物合成酶形成茉莉酸类物质,或通过GLV途径形成GLVs,为了共同的底物,GLV途径和茉莉酸途径相互竞争;而在害虫等对叶片造成伤害时,为了对抗伤害,两条代谢途径又彼此促进对抗虫害的挥发物的合成[10]。
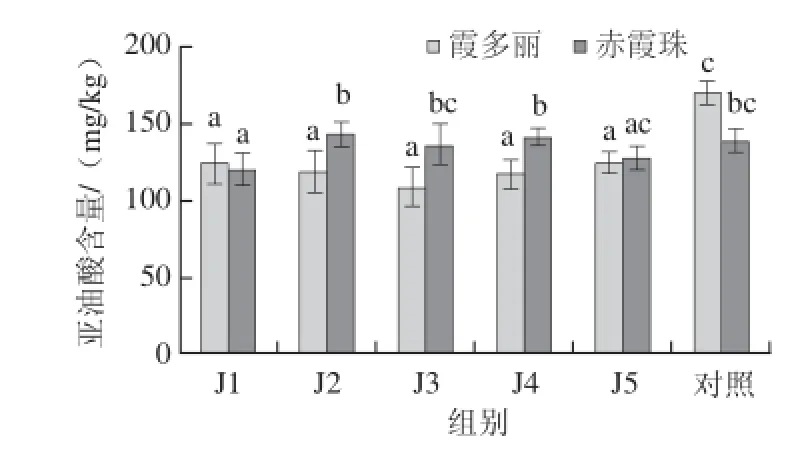
图2 MeJA对葡萄果皮亚油酸含量的影响Fig.2 Effects of MeJA application on linoleic acid content in grape berry skins
由图2可知,随MeJA处理浓度的增大,霞多丽葡萄果皮中亚油酸含量呈先降低后升高的趋势;而赤霞珠果皮中亚油酸含量呈先升高后降低的趋势。不同MeJA处理浓度下霞多丽葡萄果皮亚油酸含量均显著低于对照组。MeJA处理浓度为50 μmol/L时,赤霞珠葡萄果皮亚油酸含量显著低于对照组,MeJA处理浓度≥100 μmol/L时,亚油酸含量与对照组无显著差异。
2.4 光照处理对葡萄果皮中亚油酸含量的影响
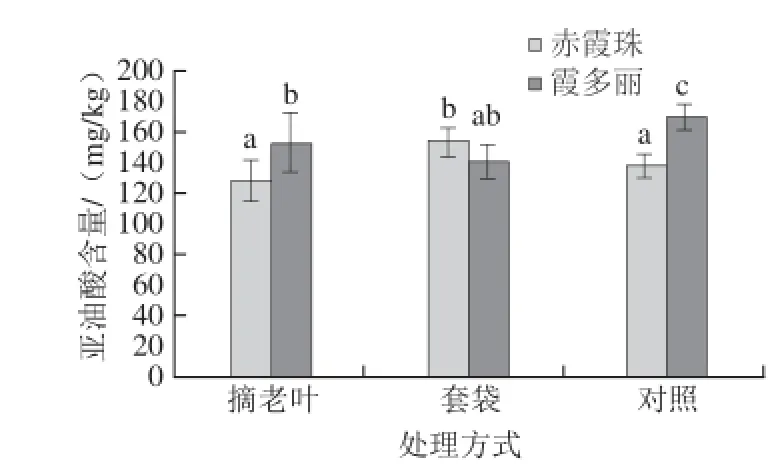
图3 光照强度对葡萄果皮亚油酸含量的影响Fig.3 Effect of light intensity on linoleic acid content grape berry skins
由图3可知,摘老叶处理后,霞多丽和赤霞珠葡萄果皮中亚油酸含量与对照组无显著差异。套袋处理后,霞多丽葡萄果皮中亚油酸含量显著低于对照组,而赤霞珠葡萄果皮中亚油酸含量显著高于对照组。由上述可知,光照强度对葡萄果皮中亚油酸含量的影响与葡萄品种有关。
2.5 采收时间对不同葡萄品种葡萄果皮中亚油酸含量的影响
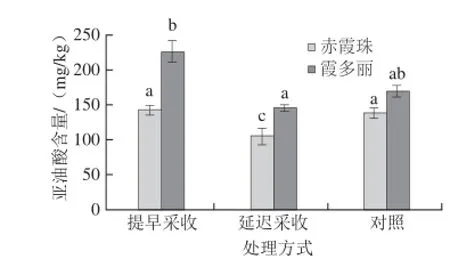
图4 不同采收时间对葡萄果皮亚油酸含量的影响Fig.4 Effect of harvest time on linoleic acid content in grape berry skins
由图4可知,提早一周采收处理,霞多丽葡萄果皮中亚油酸含量显著高于对照组,而赤霞珠葡萄果皮中亚油酸含量与对照组无显著差异。延迟一周采收处理,霞多丽和赤霞珠葡萄果皮中亚油酸含量显著低于对照组。这可能是由于亚油酸作为香气形成的前提物质被脂氧合酶转化为C6类香气物质有关。Gomez等[11]以3 种非芳香葡萄品种为研究对象,结果发现,大部分挥发性物质的含量始终保持很低的水平,而C6化合物有很高的浓度,预测C6化合物含量的变化可以用于最佳成熟度的确定。
2.6 机械损伤对不同葡萄品种葡萄果皮中亚油酸含量的影响
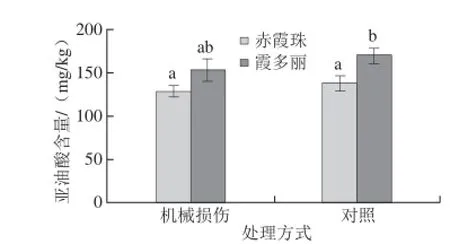
图5 机械损伤对葡萄果皮亚油酸含量的影响Fig.5 Effect of wounding on linoleic acid content in grape berry skins
机械性损伤会使葡萄浆果中脂氧合酶迅速累积,并引起茉莉酸类物质和GLVs含量的增加[7,12]。本研究发现机械损伤的上述的促进作用能一直延续至葡萄成熟。由图5可知,机械损伤处理后,霞多丽和赤霞珠葡萄果皮中亚油酸含量与对照组相比无显著差异。这可能是由于作为脂氧合酶的反应底物,当植株遇到损伤时发生应激反应,亚油酸被脂氧合酶利用最终生成茉莉酸或GLVs。
2.7 葡萄果皮中各脂肪酸组分的含量与相关性
2.7.1 霞多丽葡萄果皮中各脂肪酸组分的相关性
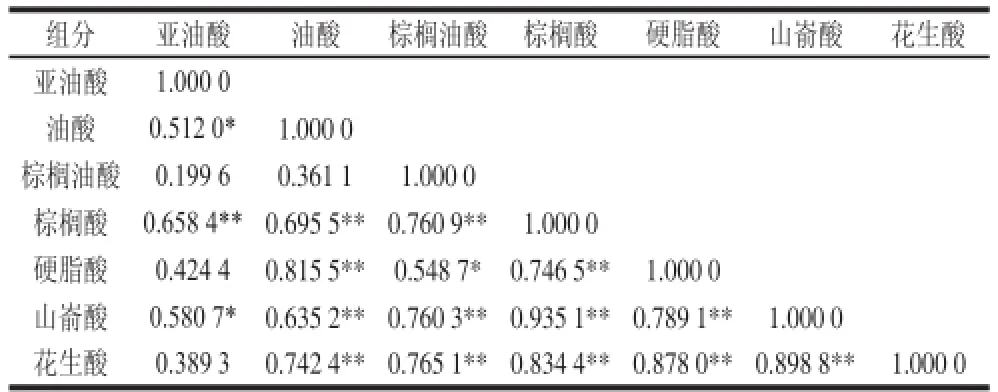
表44 霞多丽葡萄果皮中脂肪酸组分含量间相关性分析Table4 Correlation analysis among fatty acids in skins of Chardonnay berrriieess
由表4可知,在霞多丽葡萄果皮中,各脂肪酸组分呈含量间显著正相关(P<0.05)的有亚油酸与油酸、亚油酸与山嵛酸、棕榈油酸与硬脂酸;亚油酸与棕榈酸呈极显著正相关(P<0.01),油酸分别与棕榈酸、硬脂酸、山嵛酸、花生酸呈极显著正相关(P<0.01),棕榈油酸分别与棕榈酸、山嵛酸、花生酸呈极显著正相关(P<0.01),棕榈酸分别与硬脂酸、山嵛酸、花生酸呈极显著正相关(P<0.01),硬脂酸分别与山嵛酸、花生酸呈极显著正相关(P<0.01),山嵛酸与花生酸呈极显著正相关(P<0.01),其余均不相关。
2.7.2 赤霞珠葡萄果皮中各脂肪酸组分的相关性
赤霞珠葡萄果皮中主要的7 种脂肪酸为亚油酸、油酸、棕榈油酸、棕榈酸、硬脂酸、山嵛酸、花生酸(表3),对这7 种脂肪酸含量进行相关性分析。
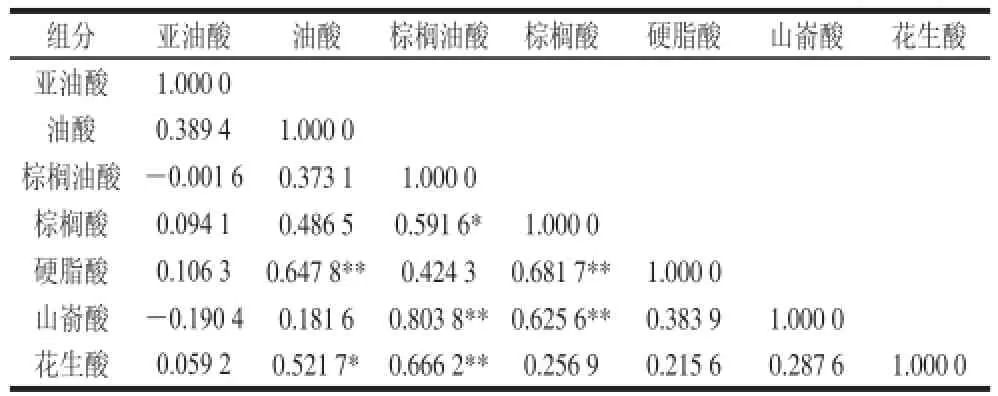
表5 赤霞珠葡萄果皮中脂肪酸组分含量间相关性分析Table5 Correlation analysis among fatty acids in skins of Cabernet Sauvignon berrriieess
由表5可知,油酸含量与硬脂酸含量间呈极显著正相关(P<0.01)、与花生酸含量呈显著正相关(P<0.05),棕榈油酸含量与棕榈酸含量呈显著正相关(P<0.05),与山嵛酸含量、花生酸含量呈极显著正相关(P<0.01),棕榈酸含量与硬脂酸含量呈极显著正相关(P<0.01),与山嵛酸含量呈显著正相关(P<0.05),其余相关性均不显著。
3 讨论与结论
3.1 外源激素处理对葡萄果皮中亚油酸的影响
实验结果显示,ABA质量浓度足够高时,GLV途径中直接底物亚油酸的消耗量大于对照组,可能对葡萄GLV途径的表达起促进作用。说明葡萄转色期的外源ABA处理会对GLV途径产生影响,并且这种影响与ABA的质量浓度和葡萄品种有关,这种不同可能与ABA促进果实发育成熟及果皮花色苷合成等有关[9,13,23-24],具体的解释还需进一步的研究。
同时,两个葡萄品种经过不同浓度的MeJA处理后,亚油酸含量基本都小于对照组,即茉莉酸对葡萄GLV途径表现出一定促进作用。有研究表示,MeJA的产生途径与GLV途径存在相互促进与竞争的关系,可以预测,随外源MeJA的浓度进一步增大,其对GLV途径的促进作用会进一步减小,然后变成抑制作用,这个推测需要进一步的研究来验证。
3.2 光照处理对葡萄果皮中亚油酸含量的影响
实验结果显示,摘老叶处理后,霞多丽和赤霞珠葡萄果皮中亚油酸含量与对照组无显著差异。套袋处理后,霞多丽葡萄果皮中亚油酸含量显著低于对照组,而赤霞珠葡萄果皮中亚油酸含量显著高于对照组,可能与赤霞珠是晚熟品种有关,套袋很好地保护了浆果免受田间不利因素的影响。本研究也验证了张云霞[15]对改善葡萄栽培措施的可行性。
3.3 采收时间对葡萄果皮中亚油酸含量的影响
实验结果显示,提早一周采收,霞多丽葡萄果皮中亚油酸含量显著高于对照组,而赤霞珠葡萄果皮中亚油酸含量与对照组无显著差异。延迟一周采收处理,霞多丽和赤霞珠葡萄果皮中亚油酸含量显著低于对照组。表明适当延迟采收,亚油酸的消耗量增大,据此推测,延迟采收对葡萄GLV途径起促进作用。若生产中适当延迟采收,GLV途径的产物绿叶香气物质含量将增加,有利于提高葡萄酒香气质量[1,17]。但是具体延迟多长时间能让葡萄香气品质达到最佳,还需要更多的研究,另外,采收时间的确定也与葡萄品种,气候条件,葡萄酒的风格等因素有关。
3.4 机械损伤处理对葡萄果皮中亚油酸含量的影响
大量研究证实,机械损伤能够引起植物体内产生大量的活性氧簇(reactive oxygen species,ROS)[25-28]。植物体内过量积累的ROS会引起GLVs的积累[7,29-30]。GLVs作为一种重要的信号分子,能启动植物的防御机制,也能够增强其他相关的抗性反应,在植物应对机械损伤、抵抗病虫害等损害时具有重要的作用。本实验的研究结果表明,机械损伤处理后,霞多丽和赤霞珠葡萄果皮中亚油酸含量与对照组无显著差异,这也表明霞多丽和赤霞珠对机械损伤具有一定的抗性。
3.5 葡萄果皮中各脂肪酸组分的含量
实验在两个葡萄品种的葡萄果皮中,发现了统一的规律:不饱和脂肪酸主要是亚油酸、油酸、棕榈油酸,且多不饱和脂肪酸总量占不饱和脂肪酸总量的70%以上,亚麻酸未检测出;主要的4 种饱和脂肪酸为(含量由高到低):棕榈酸、硬脂酸、山嵛酸、花生酸,且棕榈酸含量远远高于其他3 种酸。有研究表明,葡萄果皮油中含量最高的两种不饱和脂肪酸分别为亚油酸、油酸,亚麻酸未检测出;主要的4 种饱和脂肪酸按含量由高到低依次为:棕榈酸、硬脂酸、花生酸、山嵛酸[28]。这一结果与本实验结果基本一致,造成花生酸和山嵛酸含量多少不同的原因可能是实验材料不同,本实验材料为从新鲜成熟葡萄上剥下的葡萄果皮,而李浡等[30]采用的是葡萄果皮渣,葡萄果皮渣是经过发酵后的葡萄果皮与葡萄籽的混合物,不同于新鲜葡萄果皮。另外,在两个品种葡萄果皮中,都发现了呈显著正相关的脂肪酸组分,但品种间差异较大。
本研究了解了不同栽培措施对酿酒葡萄脂肪酸组分的影响规律,并且分析了各脂肪酸组分的相关性,对酿酒葡萄生产管理提供理论依据,具有重要的生产指导意义。
[1] CLARKE R J, BAKKER J. Wine flavour chemistry[M]. Oxford:Blackwell Pub, 2004: 272-279. DOI:10.1002/9780470995594.fmatter.
[2] MIELE A, BOUARD J, BERTRAND A. Fatty acids from lipid fractions of leaves and different tissues of Cabernet Sauvignon grapes[J]. American Journal of Enology and Viticulture, 1993, 44(2):180-186.
[3] IGLESIAS J L M, DABILA F H, MARINO J I M, et al. Biochemical aspects of the lipids in Vitis vinifera grapes (Macabeo variety)[J]. Food Science & Technology, 1991, 35(7): 705-710. DOI:10.1002/ food.19910350710.
[4] ROUFET M, BAYONOVE C L, CORDONNIER R E. Etude de la composition lipidique du raisin, Vitis vinifera L.: evolution au cours de la maturation et localisation dans la baie (lipid composition of grapevine berries, Vitis vinifera L.: changes during maturation and localization in the berry)[J]. Vitis, 1987, 26(2): 85-97.
[5] GALLANDER J F, PENG A C. Lipid and fatty acid compositions of different grape types[J]. American Journal of Enology and Viticulture, 1980, 31(1): 24-27.
[6] KUHN H, THIELE B J. The diversity of the lipoxygenase family many sequence data but little information on biological signif i cance[J]. FEBS Letters, 1999, 449(1): 7-11. DOI:10.1016/S0014-5793(99)00396-8.
[7] FEUSSNER I, WASTERNACK C. The lipoxygenase pathway[J]. Annual Review of Plant Biology, 2002, 53(4): 275-297. DOI:10.1146/ annurev.arplant.53.100301.135248.
[8] 汪仁, 沈文飚, 翟虎渠, 等. 植物种子脂氧合酶[J]. 植物生理学通讯, 2005, 41(3): 388-394. DOI:10.13592/j.cnki.ppj.2005.03.045.
[9] 鲁旭东, 吴顺. 脱落酸对植物生长发育的调控作用[J]. 孝感学院学报, 2004, 24(3): 10-14.
[10] 陈书霞, 陈巧, 王聪颖, 等. 绿叶挥发物代谢调控及分子机理研究进展[J]. 中国农业科学, 2012, 45(8): 1545-1557. DOI:10.3864/ j.issn.0578-1752.2012.08.010.
[11] GOMEZ E, MARTINEZ A, LAENCINA J. Changes in volatile compounds during maturation of some grape varieties[J]. Journal of the Science of Food and Agriculture, 1995, 67(2): 229-233. DOI:10.1002/jsfa.2740670213.
[12] 李明, 郝建军, 于洋, 等. 脱落酸(ABA)对苹果果实着色相关物质变化的影响[J]. 沈阳农业大学学报, 2005, 36(2): 189-193. DOI:10.3969/j.issn.1000-1700.2005.02.014.
[13] WASTERNACK C. Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development[J]. Annal of Botany, 2007, 100(4): 681-697. DOI:10.1093/aob/mcm079.
[14] WASTERNACK C, STENZEL I, HAUSE B, et al. The wound response in tomato: role of jasmonic acid[J]. Journal of Plant Physiology, 2006, 163(3): 297-306. DOI:10.1016/j.jplph.2005.10.014.
[15] 张云霞. 葡萄设施栽培环境调控技术[J]. 现代农业科技(园艺学), 2010, 38(5): 115. DOI:10.3969/j.issn.1007-5739.2010.05.083.
[16] 赵凌, 沈文飚, 翟虎渠, 等. 植物的脂氢过氧化物裂解酶[J]. 植物生理学通讯, 2004, 40(2): 135-140. DOI:10.13592/j.cnki. ppj.2004.02.001.
[17] 边梅娜, 白红进, 曾红, 等. 赤霞珠葡萄籽油的提取及脂肪酸成分分析[J]. 食品科学, 2013, 34(16): 297-300. DOI:10.7506/spkx1002-6630-201316061.
[18] 鞠延仑, 房玉林, 张莉, 等. 赤霞珠葡萄脂氧合酶活性测定及脂肪酸组分的变化[J]. 食品科学, 2015, 36(5): 115-119. DOI:10.7506/ spkx1002-6630-201505022.
[19] 张莉, 魏冬梅, 王华. 葡萄籽油脂肪酸GC-MS色谱分析条件优化[J]. 食品研究与开发, 2013, 34(3): 63-65. DOI:10.3969/ j.issn.1005-6521.2013.03.017.
[20] 牛翠娇, 史玉琴, 殷志萍. 气相色谱-质谱法测定食品中的反式脂肪酸[J]. 安徽农业科学, 2011, 39(6): 3649-3651. DOI:10.3969/ j.issn.0517-6611.2011.06.181.
[21] BI J L, FELTON G W. Foliar oxidative stress and insect herbivory:primary compounds, secondary metabolites, and reactive oxygen species as components of induced resistance[J]. Journal of Chemical Ecology, 1995, 21(10): 1511-1529. DOI:10.1007/BF02035149.
[22] 杨春英, 刘学铭, 陈智毅. 15 种食用植物油脂肪酸的气相色谱-质谱分析[J]. 食品科学, 2013, 34(6): 211-214. DOI:10.7506/spkx1002-6630-201306047.
[23] JU Y L, LIU M, ZHAO H, et al. Effect of exogenous abscisic acid and methyl jasmonate on anthocyanin composition, fatty acids, and volatile compounds of Cabernet Sauvignon (Vitis vinifera L.) grape berries[J]. Molecules, 2016, 21(10): 1354. DOI:10.3390/molecules21101354.
[24] LANGLOIS J, BALLESTER J, CAMPO E, et al. Combining olfactory and gustatory clues in the judgment of aging potential of red wine by wine professionals[J]. American Journal of Enology and Viticulture, 2010, 61(1): 15-22.
[25] THORDAL-CHRISTENSEN H, ZHANG Z G, WEI Y D, et al. Subcellular localization of H in plants. H2O2accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction[J]. The Plant Journal, 1997, 11(6): 1187-1194. DOI:10.1046/j.1365-313X.1997.11061187.
[26] BOWN A W, HALL D E, MACGREGOR K B. Insect footsteps on leaves stimulate the accumulation of 4-aminobutyrate and can be visualized through increased chlorophyll fluo-rescence and superoxide production[J]. Plant Physiology, 2002, 129(4): 1430-1434. DOI:10.1104/pp.006114.
[27] LEON J, ROJO E, SANCHEZ-SERRANO J J. Wound signaling in plants[J]. Journal of Experimental Botany, 2001, 52: 1-9. DOI:10.1093/ jexbot/52.354.1.
[28] WOJTASZEK P. Oxidative burst: an early plant response to pathogen infection[J]. Biochemical Journal, 1997, 322(3): 681-692. DOI:10.1042/bj3220681.
[29] MATSUI K. Green leaf volatiles: hydroperoxide lyase pathway of oxylipin metabolism[J]. Current Opinion in Plant Biology, 2006, 9(3):274-280. DOI:10.1016/j.pbi.2006.03.002.
[30] 李浡, 李双石, 吴志明, 等. 不同提取方法对葡萄果皮油中脂肪酸的影响[J]. 酿酒科技, 2012(9): 39-45. DOI:10.13746/j.njkj.2012.09.018.
Effects of Filed Management Practices and Harvest Time on Fatty Acid Composition of Cabernet Sauvignon and Chardonnay (Vitis vinifera L.) Berries Skins
JU Yanlun1, LIU Min1, ZHAO Xianfang1, ZENG Jie1, MIN Zhuo1, FANG Yulin1,2,*
(1. College of Enology, Northwest A&F University, Yangling 712100, China; 2. Shaanxi Engineering Research Center for Viti-Viniculture, Yangling 712100, China)
The effects of plant growth regulator, leaf removal, wounding, bagging and harvest time on the fatty acid composition of wine grapes were studied. The experimental treatments were conducted using Cabernet Sauvignon (Vitis vinifera L.) and Chardonnay (Vitis vinifera L.). Our results showed that 16 fatty acids were detected in grape skins. The unsaturated fatty acids mainly consisted of linoleic acid, oleic acid and palmitoleic acid; however, no linolenic acid was detected. In addition, the saturated fatty acids mainly consisted of palmitic acid, stearic acid, behenic acid and arachidic acid. Abscisic acid (ABA), methyl jasmonate (MeJA), light intensity, wounding and harvest time potentially affected the content of linoleic acid. In the berry skins of both varieties, we found a positive relationship among some fatty acids, but large differences were observed between the two varieties. In the skin of Chardonnay grapes, signif i cantly more fatty acids with a positive relationship between each other compared with Cabernet Sauvignon, and what is more, there was a stronger relationship between saturated fatty acids than between unsaturated ones. These results could be helpful for vineyard management and for improving the quality of grapes in terms of fatty acid composition.
fi eld management practices; wine grape; fatty acid composition; correlation
10.7506/spkx1002-6630-201703018
TS255.2
A
1002-6630(2017)03-0107-07
鞠延仑, 刘敏, 赵现方, 等. 栽培措施对酿酒葡萄赤霞珠和霞多丽果皮脂肪酸组分的影响[J]. 食品科学, 2017, 38(3):107-113. DOI:10.7506/spkx1002-6630-201703018. http://www.spkx.net.cn
JU Yanlun, LIU Min, ZHAO Xianfang, et al. Effects of filed management practices and harvest time on fatty acid composition of Cabernet Sauvignon and Chardonnay (Vitis vinifera L.) berries skins[J]. Food Science, 2017, 38(3): 107-113. (in Chinese with English abstract)
10.7506/spkx1002-6630-201703018. http://www.spkx.net.cn
2016-03-13
引进国际先进农业科学技术计划(948计划)项目(2014-Z20);国家现代农业(葡萄)产业技术体系建设专项(nycytx-30-2p-04);陕西省科技统筹创新工程计划项目(2013KTCL02-01)
鞠延仑(1990—),男,博士研究生,研究方向为栽培生理和分子生物学。E-mail:juyanlun@163.com
*通信作者:房玉林(1973—),男,教授,博士,研究方向为栽培生理和分子生物学。E-mail:fangyulin@nwsuaf.edu.cn
猜你喜欢
北方果树(2022年4期)2022-07-15
河北果树(2021年4期)2021-12-02
小学生学习指导(低年级)(2019年9期)2019-09-25
中国洗涤用品工业(2019年4期)2019-05-11
小学生优秀作文(低年级)(2018年12期)2018-12-13
浙江农业科学(2018年9期)2018-09-17
食品界(2016年4期)2016-02-27
物理化学学报(2015年7期)2015-12-30
中国粮油学报(2015年5期)2015-02-06
植物营养与肥料学报(2012年1期)2012-10-26
