
酿酒酵母人工杂合启动子与天然启动子活性比较
2017-02-21 08:59:13唐瑞琪熊亮白凤武赵心清上海交通大学生命科学技术学院上海0040大连理工大学生命科学与技术学院大连603
生物技术通报 2017年1期
唐瑞琪熊亮白凤武,赵心清(. 上海交通大学生命科学技术学院,上海 0040;. 大连理工大学生命科学与技术学院,大连 603)
酿酒酵母人工杂合启动子与天然启动子活性比较
唐瑞琪1熊亮2白凤武1,2赵心清1
(1. 上海交通大学生命科学技术学院,上海 200240;2. 大连理工大学生命科学与技术学院,大连 116023)
系统地比较了人工启动子PaTEF1与天然启动子PTEF1和PTDH3在不同条件下的活性差别,结果表明,人工启动子PaTEF1的活性并不是在任何条件下都高于天然启动子的活性,而与宿主、培养基以及细胞生长阶段有关。在3个宿主背景中,BY4741中PaTEF1的活性最高,而LX03中最低。在YPD100中,人工启动子PaTEF1活性分别为天然启动子PTEF1和PTDH3活性的1.4-4.6倍和0.9-2.0倍;而在YPE(5%和7%)中,PaTEF1活性与PTEF1和PTDH3活性之比在0.7-1.3以及0.8-1.3之间。在YPE中培养时,PaTEF1的活性为在YPD100中培养时的1.7-2.0倍,PTEF1和PTDH3的活性在YPE中为YPD100中培养时的2.7-7.1倍和1.3-3.4倍,启动子在YPE中的活性较YPD100中更高,但人工启动子的活性变化较天然启动子更小。此外,在不同遗传背景的菌株中,启动子活性从对数期早期到对数期中期和从对数期中期到稳定期的变化趋势不同。
酿酒酵母;人工杂合启动子;TEF1启动子;宿主菌株;环境条件
酿酒酵母是目前最常用的生产生物燃料和生物基化学品的底盘细胞之一。作为重要的细胞工厂,对酿酒酵母细胞中代谢通路进行精细调节对于提高生物产品的产量、产率和生产强度至关重要[1]。启动子作为转录调控以及合成生物学的重要元件,被广泛应用于基因表达的研究和菌株构建中。天然的启动子主要可以分为组成型启动子和诱导型启动子两类,前者能维持恒定的基因转录水平,而后者的表达则依赖于特殊诱导剂的添加[2]。研究者们鉴定了许多不同的启动子,包括天然启动子和启动子的突变体[3,4]。研究表明,启动子中的元件可以作为模块进行合成生物学改造来调节启动子的表达强度,通过将不同的上游增强元件(UASCLB和UASCIT)与不同的天然启动子核心(PGPD、PCYC、PTEF和PLEUM)串联,得到的杂合启动子的活性是相应的天然启动子活性的1.63-10.02倍[5]。作为一种新的遗传操作元件,人工启动子的高强度和稳定性对于精细调节细胞中关键途径的表达具有重要意义。
目前发表的大部分研究结果倾向于创造更高的启动子多样性,然而,研究个别启动子在不同条件下的启动活性同样重要。启动子的活性会受到多种环境因素的影响,例如,酿酒酵母多个启动子在不同碳源(葡萄糖、蔗糖、乳糖和乙醇)中的活性具有显著差异[6]。当细胞处在不同的生长时期时,启动子的活性也略有不同。此外,启动子在碳源转换期(diauxic shift)的活性也存在明显差异[6]。基因的最优表达水平在不同宿主菌株中也不尽相同,并受到宿主遗传背景的影响[7]。
葡萄糖是细胞工厂生产目标产物时最常见的碳源,随着发酵的进行,当葡萄糖耗尽之后,细胞转换为利用培养基中其他碳源的模式。乙醇既是酿酒酵母乙醇发酵中的主要代谢产物,在发酵后期乙醇也可能被当做支持菌株生长的碳源。在某些高附加值产品的生产过程中,乙醇也被用作生产碳源,例如,以乙醇作为碳源有利于青蒿素的生产[8]。高浓度乙醇也对宿主菌株产生明显的抑制作用[9],因此,研究不同浓度乙醇存在条件下启动子的活性具有重要参考价值。
虽然目前已经有不少关于启动子在不同碳源和不同细胞生长时期的研究,但是大部分研究只对单一宿主中或单一条件下的不用启动子活性进行了比较,没有同时比较不同宿主、不同培养基以及不同细胞生长阶段的启动子活性的研究。此外,现有对人工启动子和天然启动子活性比较的研究还非常少。本研究选取了由3个串联的上游增强元件UASCLB和TEF1启动子核心组装而成的人工启动子P[10]aTEF1与天然启动子进行了比较,研究人工启动子PaTEF1与天然启动子PTEF1以及PTDH3在不同宿主及同一宿主不同细胞生长阶段或者不同培养基条件下的活性差别,为进一步应用酿酒酵母人工启动子进行合成生物学研究以及人工启动子的生物技术应用和进一步改造提供参考。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒和引物 本研究所用到的菌株、质粒和引物,如表1-表3所示。
1.1.2 培养基 LB培养基:5 g/L酵母浸粉,10 g/L蛋白胨,10 g/L NaCl;YPD100培养基:10 g/L酵母浸粉,20 g/L蛋白胨,100 g/L葡萄糖;YPE培养基含10 g/L酵母浸粉和20 g/L蛋白胨,5% YPE和7% YPE分别加入5%(V/V)和7%(V/V)的无水乙醇;YPD活化培养基:10 g/L酵母浸粉,20 g/L蛋白胨,20 g/L葡萄糖。固体培养基添加琼脂粉至20 g/L。大肠杆菌转化子筛选时添加100 mg/L氨苄青霉素;酿酒酵母转化子筛选和细胞培养时添加200 mg/L潮霉素。
1.2 方法
1.2.1 报告载体的构建 首先以pKT127为模板,yEGFP-F/R为引物PCR扩增得到带多克隆位点Linker和ADH1终止子序列的yEGFP片段,经Xma I/Kpn I双酶切后连接到用相同酶处理的pRS41H载体中,得到载体pRS41H-yEGFP。以S288c基因组为模板,PTEF1-F/R和PTDH3-F/R为引物分别进行PCR,扩增得到PTEF1和PTDH3启动子片段;以p416-UASCLB(3X)-PTEF为模板,PaTEF1-F/R为引物PCR扩增得到PaTEF1启动子片段。人工启动子PaTEF1比野生型PTEF1多了3个重复的CLB2启动子激活序列UASCLB,该序列位于基因CLB2起始密码子上游-867至-627,长度为240 bp[5]。将上述启动子片段通过相应的酶切位点连接到pRS41H-yEGFP,得到相应的报告载体 pRS41H-PTEF1-yEGFP、pRS41H-PTDH3-yEGFP和pRS41H-PaTEF1-yEGFP。
1.2.2 菌株的构建和筛选 将测序验证正确的启动子报告载体及其空载(pRS41H-yEGFP)通过醋酸锂转化法[14]分别转化到酿酒酵母BY4741、Ethanol Red和CEN.PK 113-5D LX03(下文简称LX03)菌株中。在含有200 mg/L潮霉素的抗性YPD固体培养基中筛选转化子。挑取单菌落,在含有200 mg/L潮霉素的YPD培养基中扩大培养,含有启动子的转化子通过荧光显微镜检测细胞GFP荧光。空载体的转化子在含有200 mg/L潮霉素的YPD中培养后提取质粒,转化大肠杆菌并进一步验证质粒的序列。
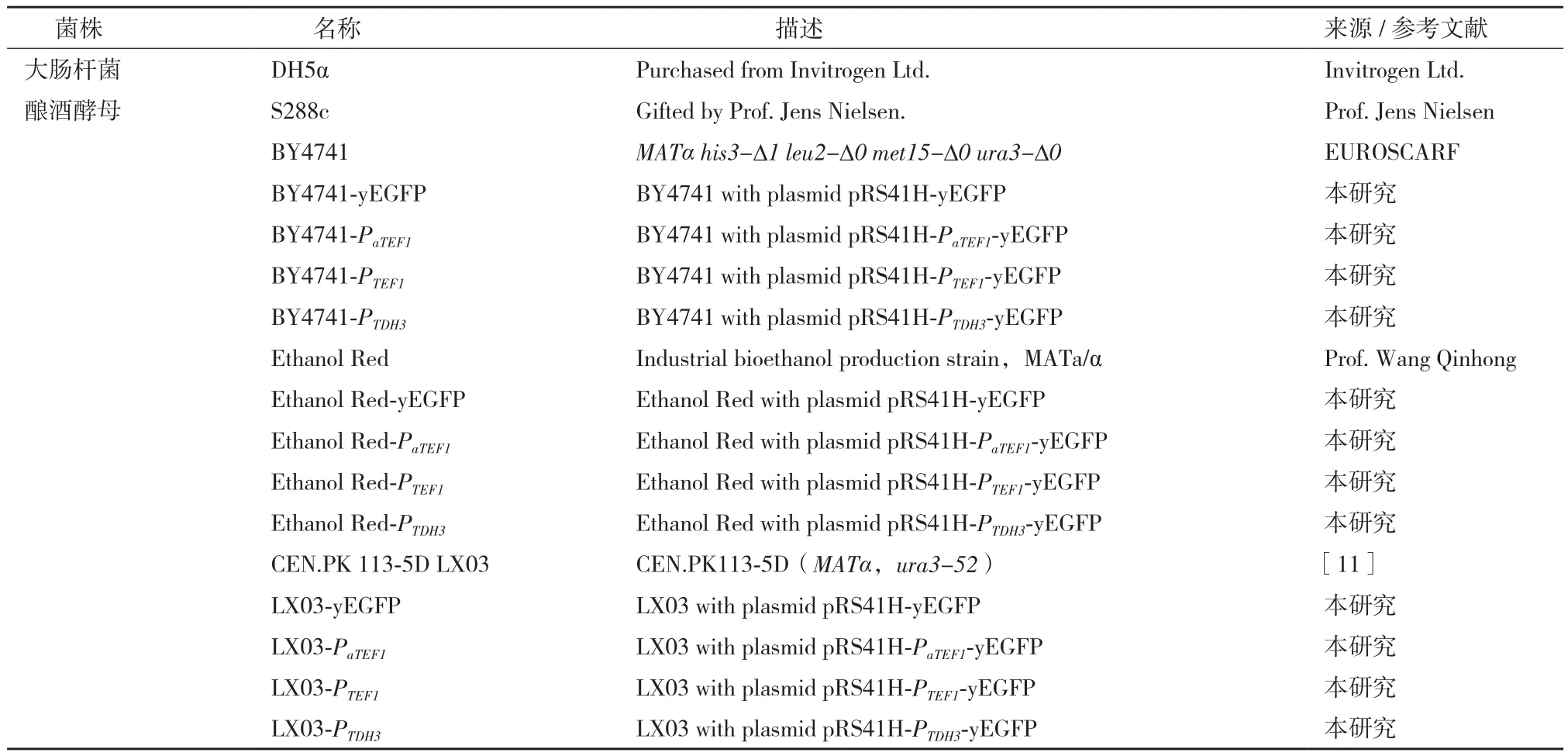
表1 本研究所用的菌株

表2 本研究所用的引物

表3 本研究所用的质粒
1.2.3 酿酒酵母细胞培养 为了高通量检测不同宿主以及不同培养基中的启动子活性,本研究采用方形U底全挡板24孔板(常州英德生物科技有限公司,YD010124B),在30℃和200 r/min条件下,对上述12个酿酒酵母菌株进行培养。培养体积为3 mL,并比较了各菌株在YPD100、5% YPE和7%YPE三种培养基条件下的生长和启动子活性。取在-80℃甘油管中保存的菌在5 mL YPD活化培养基中培养过夜后,将活化后的菌液按初始OD600为0.2转接至上述培养基中进行培养,并取样检测细胞生长及启动子活性。每个菌株在培养和检测时设置两个平行。1.2.4 细胞生长的测定 每隔12 h(YPD100)或24 h(YPE)取0.1 mL不同培养基中培养的菌液,稀释到合适浓度后,用酶标仪(Thermo-Fisher ScientificTM,MultiskanTMGO,MA,USA),在600 nm波长下检测细胞浓度。
1.2.5 GFP荧光检测 取24孔板中培养的菌液0.1 mL,在4℃下3 000 ×g离心3 min收集细胞,用1 mL预冷的ddH2O洗涤细胞,离心后弃上清并用预冷的ddH2O重悬细胞至适合流式细胞仪检测的浓度。通过流式细胞仪(BD FACSAria II®,NJ,USA)检测单个细胞中yEGFP的荧光强度,进而对启动子强度进行分析。yEGFP荧光通过蓝光(488 nm)激发,用530/30规格的滤光片检测。每份样品(细胞悬液)共收集50 000个细胞进行检测和统计。通过前向散射光检测器(FSC-A)和侧向散射光检测器(SSC-A)对细胞大小进行检测。GFP荧光与细胞的大小有关,本研究根据Peng等[6]描述的方法进行校正。
2 结果
2.1 报告载体和菌株的构建
根据1.2.1所描述的方法,构建得到不含启动子的pRS41H-yEGFP载体和含有3种不同启动子的报 告 载 体:pRS41H-PaTEF1-yEGFP、pRS41H-PTEF1-yEGFP和pRS41H-PTDH3-yEGFP(图1)。将上述4个载体分别转化酿酒酵母BY4741、Ethanol Red和LX03菌株,得到表1所示的12个报告载体过表达菌株,并用于后续研究。
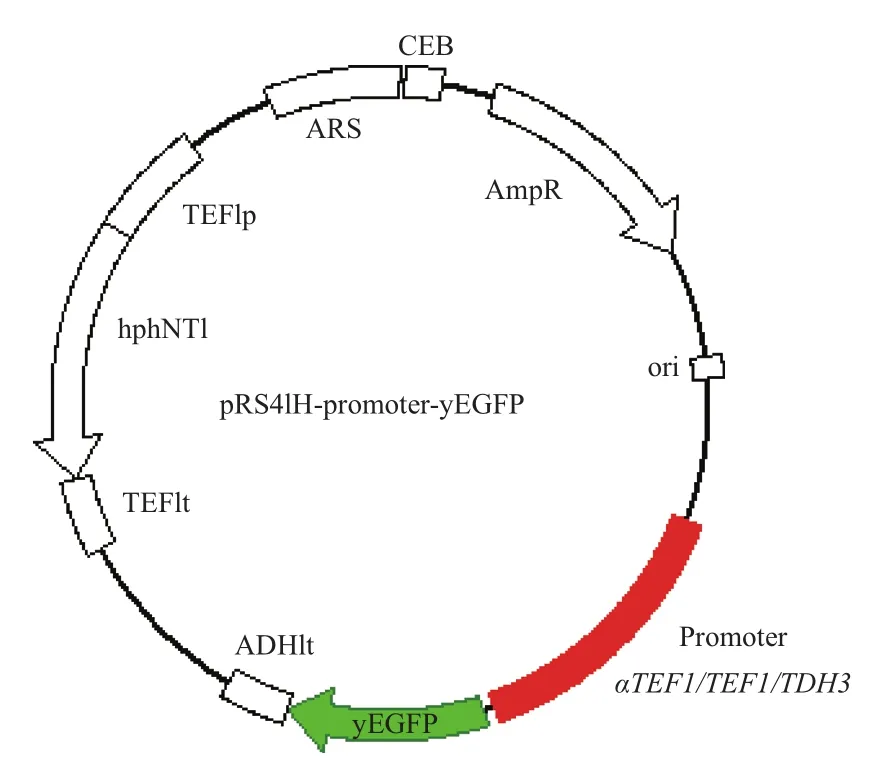
图1 报告载体示意图
2.2 细胞生长的比较
酿酒酵母BY4741、Ethanol Red和LX03菌株在YPD100、5% YPE和7% YPE条件下的生长情况如图2所示。YPD100中培养的各个菌株无明显延滞期,在36 h之后进入稳定期,BY4741背景的菌株生长速率显著低于以另外两个菌为宿主的菌株,而以Ethanol Red为宿主的菌株生长速率略高于以LX03为宿主的菌株。同时,5% YPE和7% YPE中BY4741背景的菌株生长速率也要慢于以Ethanol Red或LX03为宿主的菌株,但LX03背景的菌株生长速率略快于以Ethanol Red为宿主的菌株。
2.3 各菌株在不同条件下的启动子强度
菌株在不同培养基中生长情况不同,因此经过预实验确定各个菌株在3种培养基中培养时的生长时期,并按照1.2.5描述的方法检测细胞荧光。
酿酒酵母BY4741、Ethanol Red和LX03菌株在YPD100、5% YPE和7% YPE条件下的GFP荧光强度如图3所示。含有空载体的菌株检测到的荧光强度几乎可以忽略不计,因此细胞中的荧光强度直接反映相应的启动子强度。在YPD100中3种遗传背景的菌株的人工启动子PaTEF1活性均高于天然启动子PTEF1活性,前者是后者的1.4-4.6倍;而BY4741、Ethanol Red和LX03菌株的PaTEF1活性分别是PTDH3活性的0.93、2.03和1.14倍。在5%和7% YPE中,LX03背景的菌株中PaTEF1活性较PTEF1更高,较PTDH3更低;而Ethanol Red背景的菌株中PaTEF1的活性较PTEF1更低,较PTDH3更高,BY4741背景的菌株中PaTEF1的活性较PTDH3更高,与PTEF1相比则是有高有低;3个菌株在YPE培养基中的PaTEF1活性与PTEF1和PTDH3活性之比分别在0.7-1.3和0.8-1.3之间。在不同遗传背景的菌株中,启动子在YPE中的活性均较YPD中更高,在YPE中培养时细胞的PaTEF1、PTEF1和PTDH3启动子活性分别为在YPD100中培养时的1.7-2.0、2.7-7.1和1.3-3.4倍。在3种培养基中,PaTEF1和PTDH3的活性在BY4741背景的菌株中最高,在LX03背景的菌株中最低;而PTEF1的活性则在Ethanol Red背景的菌株中最高,在LX03背景的菌株中最低。
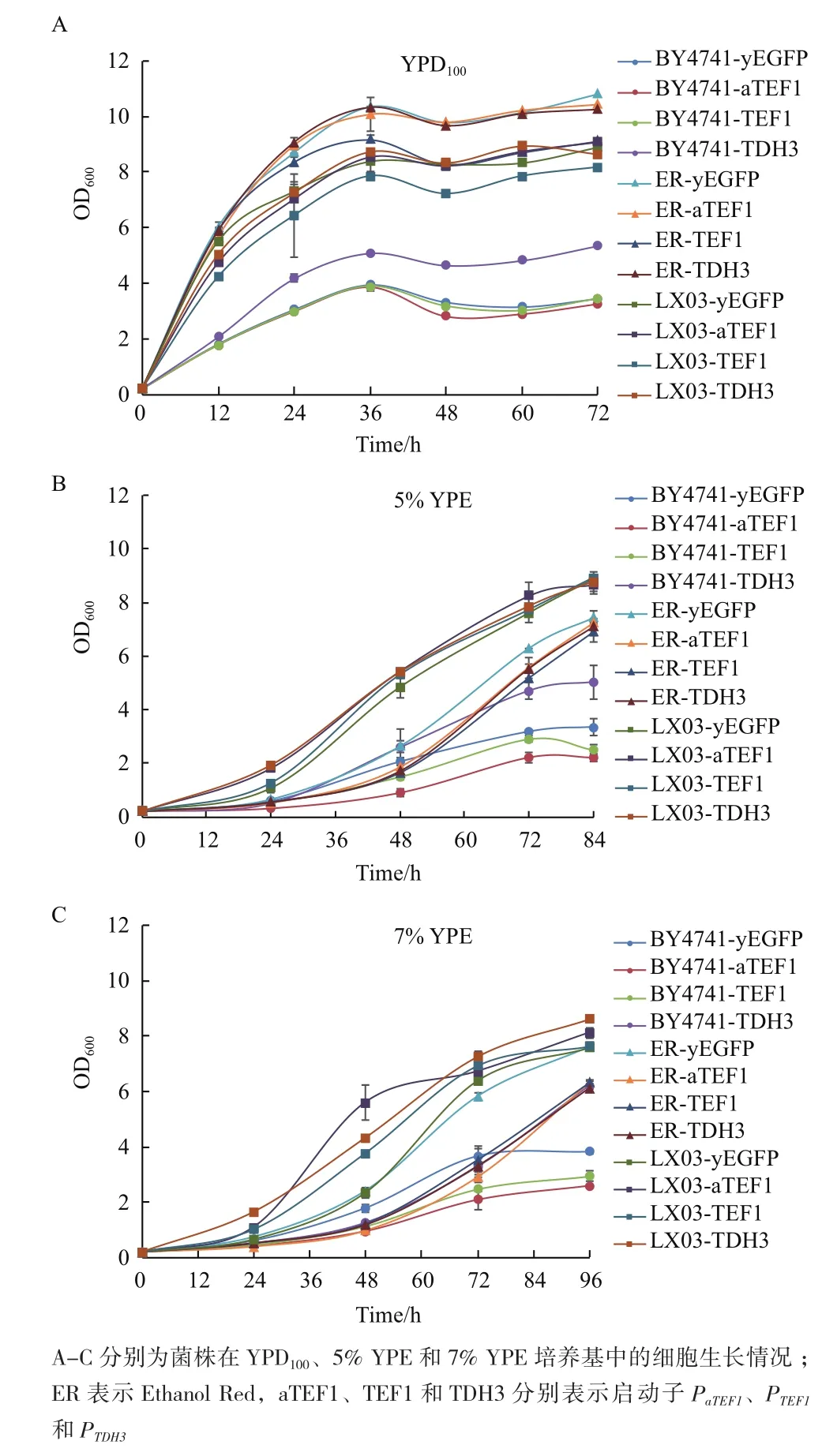
图2 菌株在不同培养基条件下的生长情况比较
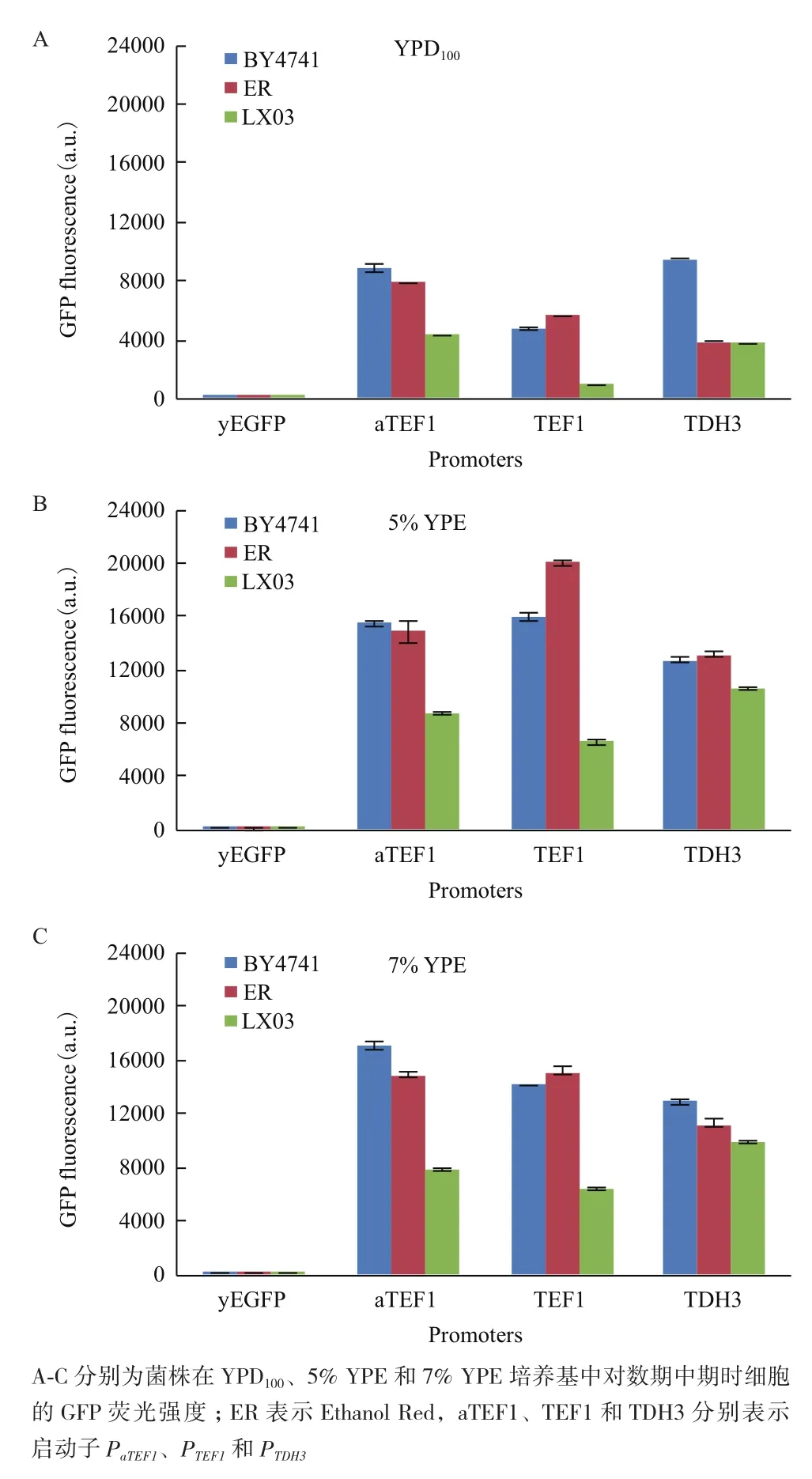
图3 菌株在不同培养基条件下的GFP荧光比较
此外,对3个菌株在YPD100培养基中,对数期早期、对数期中期和稳定期的细胞荧光强度进行了比较,相应的取样时间点分别为8、24和48 h(图4,表4)。启动子活性从对数期早期到对数期中期的变化较对数期中期到稳定期的变化大。对比对数期中期和对数期早期,PTDH3启动子在3种背景的菌株中启动子活性变化小,而PaTEF1启动子在BY4741背景的菌株中对数期中期的活性较对数期早期的活性减弱(前者是后者的0.73倍),PTEF1启动子在LX03背景的菌株中活性增强(前者是后者的1.43倍)。
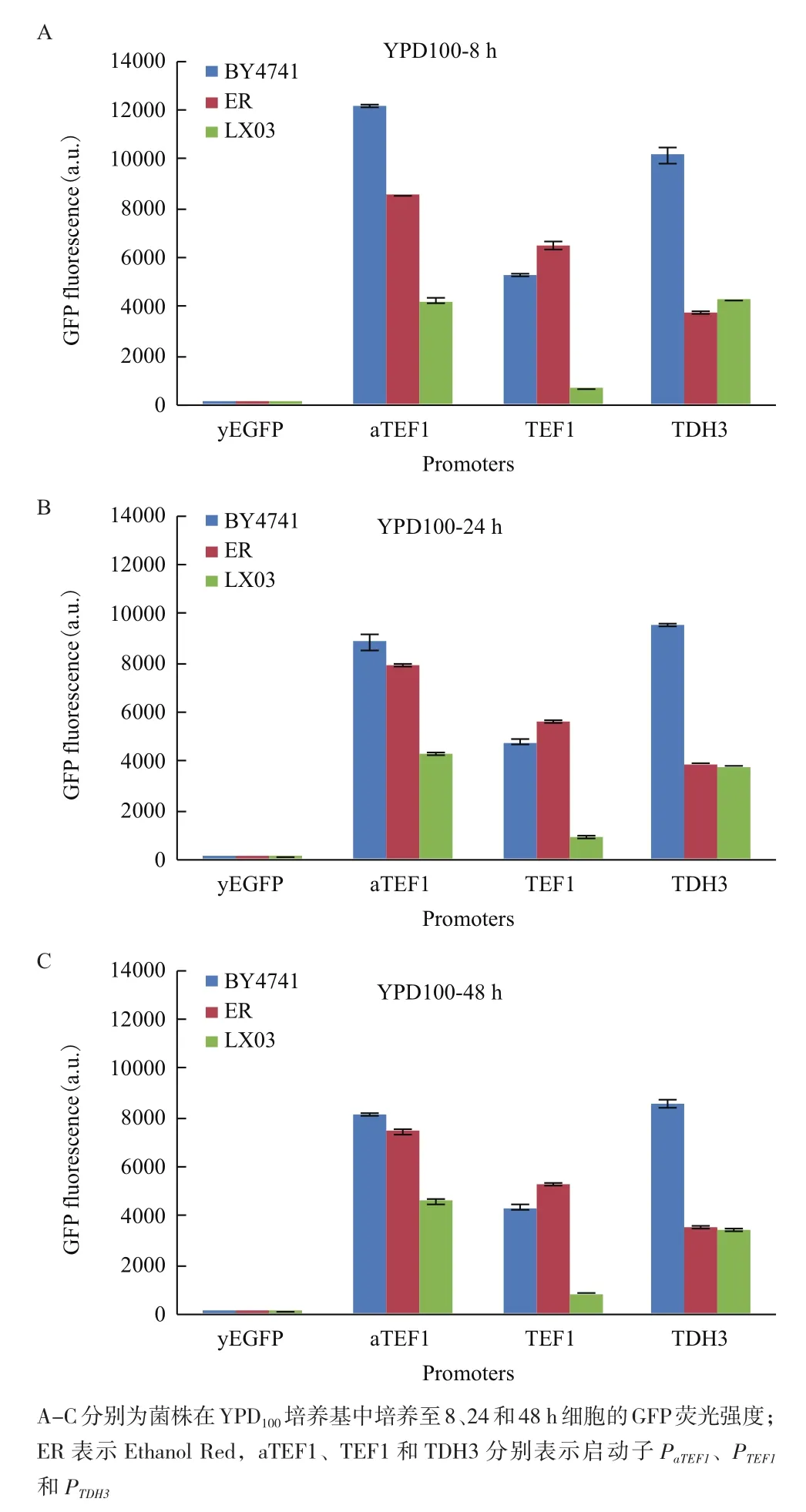
图4 菌株在YPD100培养基中不同细胞生长阶段的GFP荧光比较
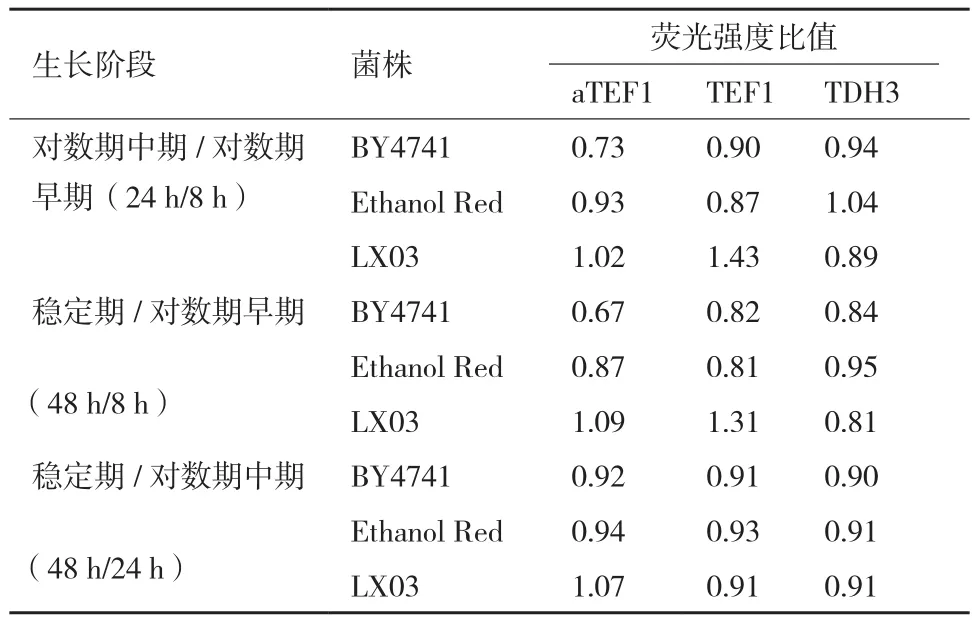
表4 菌株在YPD100培养基中不同生长阶段的GFP荧光强度比较
3 讨论
酿酒酵母是普遍使用的底盘细胞,对其细胞中关键代谢途径的基因表达强度进行精细调节对于提高生产菌株的性能至关重要。细胞内代谢的调控水平包括转录水平、翻译水平和翻译后修饰水平。蛋白质的翻译后修饰调控较为复杂,不便于进行遗传操作。相较于翻译水平,转录水平的调控对基因表达水平的提升倍数更多,可达到102数量级,而翻译水平调控对基因表达水平的提升却很低,例如密码子优化使酿酒酵母双加氧酶基因表达水平提高2.9倍左右[15]。因此,通过采用不同强度的启动子来调控转录水平进而改变代谢流是一个常用的方法[16]。例如,通过在RIM15基因的5'-UTR区域插入PCK1基因的启动子可以抑制RIM15基因在早期葡萄糖丰富时的表达而激活其在后期乙醇发酵时的表达,进而提高乙醇发酵性能[17]。此外,在其他代谢途径中,研究者们也通过选择不同强度的启动子来调控不同酶的表达水平。如我国学者[18]对酿酒酵母中萜类化合物合成途径中多达12个基因的启动子活性进行了表征,并采用具有不同活性的启动子对代谢途径进行了重新设计,进而获得了角鲨烯、油酸和麦角固醇产量显著提高的酿酒酵母菌株。在过去20多年里,研究者们鉴定了许多不同的启动子,包括天然的启动子及其突变体。Partow等[19]对多个组成型启动子以及诱导型启动子PGAL1和PGAL10在葡萄糖和半乳糖中的活性进行了表征;Alper等[5]通过对野生型TEF1启动子进行突变得到了活性跨度较大的TEF1启动子突变体;Peng等[6]则同时鉴定了19个启动子在不同碳源中的活性。在现有启动子的基础上,研究者们又创造性地将启动子中的调控元件进行了组装,得到了具有更强活性的人工启动子[4]。
与挖掘启动子的多样性相比,研究个别启动子在不同条件下,尤其是工业发酵环境条件下的启动活性和稳定性对于提升细胞工厂的性能同样重要。基因的最优表达水平在不同宿主中也不尽相同,并受到宿主遗传背景的影响。因此本研究选择了不同宿主进行启动子活性分析,包括BY4741、Ethanol Red和LX03,其中BY4741为常用的实验室菌株,Ethanol Red为工业菌株,LX03则是本课题组之前研究工作中构建的木糖代谢菌株[10]。Partow等[19]研究发现,当把8 h时的PTEF1启动子活性定义为100%时,24 h和48 h时PTEF1启动子活性分别为156%和136%。在本研究中,仅LX03背景菌株在葡萄糖条件下的启动强度变化与Partow等[19]的研究结果一致,而BY4741和Ethanol Red背景菌株中PTEF1在24 h和48 h相对于8 h的启动子活性均出现了下降(表4)。Partow等[19]所用的菌株为CEN.PK 113-5D,与LX03的出发菌株相同。由此说明,在相同遗传背景的菌株中,启动子活性随着细胞生长阶段的变化趋势相同。PaTEF1在YPD100中的活性高于PTEF1的活性,而在YPE中则表现为在LX03背景的菌株中PaTEF1的活性高于PTEF1的,在Ethanol Red背景的菌株中PaTEF1的活性较PTEF1更低,而在BY4741中有高有低。在3种培养基中,PaTEF1的活性在BY4741背景的菌株中最高,在LX03背景的菌株中最低。而PTEF1的活性则在Ethanol Red背景的菌株中最高,在LX03背景的菌株中最低。表明启动子活性会随宿主遗传背景的不同而变化。同一启动子的活性在不同宿主中不尽相同,这可能与不同宿主的代谢调控网络不同有关。此外,人工启动子的活性并非总是高于天然启动子活性,而是随着酿酒酵母遗传背景和培养基条件的不同发生变化。
研究表明,启动子的活性也会受到包括碳源和生长时期等多种因素的影响[19,20],因此本研究检测了3种培养基条件下对数期早期、对数期中期和稳定期的启动子活性。葡萄糖是细胞工厂生产目标产物时最常用的碳源,当葡萄糖耗尽之后,细胞转换为利用培养基中其他碳源的模式。乙醇既是酿酒酵母乙醇发酵中的主要代谢产物,在发酵后期也作为细胞生长的碳源。乙醇也被用作某些高附加值产品,如角鲨烯、游离脂肪酸等[18]生产过程的碳源,研究表明以乙醇作为碳源有利于青蒿素的生产[8]。此外,高浓度乙醇对宿主菌株产生明显的抑制作用[21]。因此研究乙醇为碳源条件下的启动自强度也具有重要的意义。Peng等[6]研究了以2%乙醇为碳源时启动子活性,发现PTEF启动子活性较PPGK1和PTDH3等启动子的活性更强。Yamauchi等[22]发现BTN2启动子可在高浓度(>9%,V/V)乙醇条件下强表达,但在温和乙醇胁迫条件下PBTN2活性则相对较低,采用该启动子也能够在高浓度乙醇条件下启动CUR1、GIC2以及YUR1等异源基因的表达。本研究对不同宿主中野生型启动子和人工启动子在3种培养基YPD100、5% YPE和7% YPE的活性进行了研究和比较。其中,培养基YPE中5%和7%的乙醇既可作为碳源又是一种细胞胁迫因子,相较于2%的乙醇浓度也更接近生产发酵过程中的乙醇浓度。有趣的是,启动子PaTEF1、PTEF1和PTDH3在YPE中的活性均高于在YPD100中的活性,前者是后者的1.3-7.1倍,提示乙醇对上述启动子有一定的激活作用。YPE中启动子活性较YPD中更强,可能是菌株响应乙醇胁迫的一种表现。通过菌株在不同培养基条件下的生长情况(图2)可知,在YPE条件下培养时,菌株的延滞期延长,即乙醇对菌株的生长产生了抑制,这可能使得细胞对蛋白质的需求更大,增加了启动子活性。另外,转录因子与启动子的结合对启动子的活性影响较大[23,24],乙醇作为一种胁迫的同时也是一种不同于葡萄糖的碳源,可能影响转录因子的结合,从而影响启动子活性。
随着合成生物学的发展和启动子研究的深入[4],人工杂合启动子成为一种新的元件构建方法。人工杂合启动子由上游激活序列和核心启动子组合而成,Alper等通过将不同的上游增强元件(UASCLB和UASCIT)与不同的天然启动子核心(PGPD、PCYC、PTEF和PLEUM)串联得到的杂合启动子的活性是相应的天然启动子活性的1.63-10.02倍。尽管人工启动子比野生型启动子活性更强,但前人的研究只考察了酿酒酵母BY4741菌株在YSC-URA(20 g/L glucose)培养基中培养48 h时的人工启动子活性,未对人工启动子在不同宿主、不同碳源以及不同生长时期的活性进行进一步研究。本研究选取的人工启动子是Alper等[5]报道的将3个串联的240-bp UASCLB与PTEF1核心组装而成的启动子UASCLB(3X)- PTEF1,该启动子在葡萄糖条件下的活性是野生型PTEF1的1.63倍,并研究了人工启动子及其天然启动子PTEF1以及常用的强启动子PTDH3在不同宿主、不同培养基条件和细胞不同生长阶段的活性。值得一提的是,在YPD100和YPE培养基中,人工启动子PaTEF1的活性变化倍数为1.7-2.0,而天然启动子PTEF1和PTDH3的变化倍数分别为2.7-7.1和1.3-3.4,这可能从侧面反映出人工启动子的活性在不同培养基中较天然启动子更稳定。
4 结论
本研究系统全面地分析和比较了人工启动子和天然启动子在不同条件下和不同宿主中的性能。宿主的遗传背景和培养基条件对启动子活性的影响较大,同时,启动子活性可能随着细胞生长阶段的不同而不同,另外,在不同遗传背景的菌株中,启动子活性从对数期早期到对数期中期和从对数期中期到稳定期的变化趋势不同。这些结果表明启动子的选用需要考虑宿主遗传背景和不同的培养基条件以及基因的表达在不同细胞生长阶段的需求。人工启动子在不同培养基中的活性比天然启动子更加稳定,因此在需要基因稳定表达的情况下,人工启动子可能是一种更好的选择。
致谢:感谢美国德克萨斯大学奥斯汀分校Hal S. Alper教授惠赠质粒p416-UASCLB(3X)-PTEF。
[1]Kavšček M, Stražar M, Curk T, et al. Yeast as a cell factory:current state and perspectives[J]. Microbial Cell Factories, 2015, 14(1):1.
[2]Mnaimneh S, Davierwala AP, Haynes J, et al. Exploration of essential gene functions via titratable promoter alleles[J]. Cell, 2004, 118(1):31-44.
[3]Weinhandl K, Winkler M, Glieder A, et al. Carbon source dependent promoters in yeasts[J]. Microbial Cell Factories, 2014, 13(1):1.
[4]Blazeck J, Alper HS. Promoter engineering:recent advances in controlling transcription at the most fundamental level[J]. Biotechnology Journal, 2013, 8(1):46-58.
[5]Blazeck J, Garg R, Reed B, et al. Controlling promoter strength and regulation in Saccharomyces cerevisiae using synthetic hybrid promoters[J]. Biotechnology Bioengineering, 2012, 109:2884-2895.
[6]Peng B, Williams TC, Henry M, et al. Controlling heterologous gene expression in yeast cell factories on different carbon substrates and across the diauxic shift:a comparison of yeast promoter activities[J]. Microbial Cell Factories, 2015, 14:91.
[7]Alper H, Fischer C, Nevoigt E, et al. Tuning genetic control through promoter engineering[J]. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(36):12678-12683.
[8]Westfall PJ, Pitera DJ, Lenihan JR, et al. Production of amorphadiene in yeast, and its conversion to dihydroartemisinic acid, precursor to the antimalarial agent artemisinin[J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109:E111-E118.
[9]Zhao XQ, Bai FW. Mechanisms of yeast stress tolerance and its manipulation for efficient fuel ethanol production[J]. Journal of Biotechnology, 2009, 144(1):23-30.
[10]Blazeck J, Liu L, Redden H, et al. Tuning gene expression in Yarrowia lipolytica by a hybrid promoter approach[J]. Applied and Environmental Microbiology, 2011, 77(22):7905-7914.
[11]熊亮, 程诚, 李凯, 等. 菊芋秸秆高浓度物料分步糖化及乙醇发酵[J]. 应用与环境生物学报, 2016, 22(3):382-387.
[12]Sheff MA, Thorn KS. Optimized cassettes for fluorescent protein tagging in Saccharomyces cerevisiae[J]. Yeast, 2004, 21:661-670.
[13]Taxis C, Knop M. System of centromeric, episomal, and integrative vectors based on drug resistance markers for Saccharomyces cerevisiae[J]. Biotechniques, 2006, 40(1):73-78.
[14]Gietz RD, Schiestl RH. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method[J]. Nature Protocols, 2007, 2(1):31-34.
[15]Leavitt JM, Alper HS. Advances and current limitations in transcript-level control of gene expression[J]. Current Opinion in Biotechnology, 2015, 34:98-104.
[16]Blazeck J, Alper H. Systems metabolic engineering:Genome-scale models and beyond[J]. Biotechnology Journal, 2010, 5(7):647-659.
[17]Watanabe D, Kaneko A, Sugimoto Y, et al. Promoter engineering of the Saccharomyces cerevisiae RIM15 gene for improvement of alcoholic fermentation rates under stress conditions[J]. Journal of Bioscience and Bioengineering, 2016, pii:S1389-1723(16)30202-X. doi:10. 1016/j. jbiosc. 2016. 08. 004.
[18]Rasool A, Zhang G, Li Z, et al. Engineering of the terpenoid pathway in Saccharomyces cerevisiae co-overproduces squalene and the non-terpenoid compound oleic acid[J]. Chemical Engineering Science, 2016, 152:457-467.
[19]Partow S, Siewers V, Bjørn S, et al. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae[J]. Yeast, 2010, 27(11):955-964.
[20]Vickers CE, Bydder SF, Zhou Y, et al. Dual gene expression cassette vectors with antibiotic selection markers for engineering in Saccharomyces cerevisiae[J]. Microbial Cell Factories, 2013, 12:96.
[21]Bai FW, Anderson WA, Moo-Young M. Ethanol fermentation technologies from sugar and starch feedstocks[J]. Biotechnology Advances, 2008, 26(1):89-105.
[22]Yamauchi Y, Izawa S. Prioritized expression of BTN2 of Saccharomyces cerevisiae under pronounced translation repression induced by severe ethanol stress[J]. Frontiers in Microbiology, 2016, 7:1319.
[23]Redden H, Alper HS. The development and characterization of synthetic minimal yeast promoters[J]. Nature Communications, 2015, 6:7810.
[24]Ganapathi M, Palumbo MJ, Ansari SA, et al. Extensive role of the general regulatory factors, Abf1 and Rap1, in determining genomewide chromatin structure in budding yeast[J]. Nucleic Acids Research, 2011, 39(6):2032-2044.
(责任编辑 马鑫)
Activity Comparison of the Artificial Hybrid Promoter with Its Native Promoter in Saccharomyces cerevisiae
TANG Rui-qi1XIONG Liang2BAI Feng-wu1,2ZHAO Xin-qing1
(1. School of Life Science and Biotechnology,Shanghai Jiao Tong University,Shanghai 200240;2. School of Life Science and Biotechnology,Dalian University of Technology,Dalian 116023)
The responses of promoter strengths of artificial TEF1 promoter(PaTEF1)and native promoter PTEF1as well as PTDH3were comprehensively compared. The strength of PaTEF1was not always higher than PTEF1,but varied along with the genetic background of host,medium,and cell growth phase. Among the three investigated hosts,PaTEF1showed the highest strength in BY4741,while the lowest in LX03. The strengths of PaTEF1were 1.4-4.6 and 0.9-2.0 times of those of native promoters(PTEF1and PTDH3)in YPD100medium,respectively. The activities of PaTEF1in YPE(5% and 7%)were 0.7-1.3 and 0.8-1.3 times of PTEF1and PTDH3. The activity of PaTEF1in YPE was 1.7-2.0 times of that in YPD100,whereas 2.7-7.1 and 1.3-3.4 times for PTEF1and PTDH3,respectively. The activities of the promoters in YEP were higher than those in YPD100,however,the activity variation of artificial promoters was less than that of native promoter. In addition,the variation trends of promoter strengths from early-log to mid-log phase and from mid-log to stationary phase varied in different host strains.
Saccharomyces cerevisiae;artificial hybrid promoter;TEF1 promoter;host strain;environmental condition
10.13560/j.cnki.biotech.bull.1985.2017.01.013
2016-10-20
国家自然科学基金项目(31461143029,5151101168)
唐瑞琪,女,博士研究生,研究方向:酿酒酵母代谢工程改造;E-mail:rq_tang@sjtu.edu.cn
赵心清,女,博士,教授,研究方向:酿酒酵母及丝状真菌代谢工程改造和生物能源生产;E-mail:xqzhao@sjtu.edu.cn
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05 07:20:20
昆钢科技(2021年6期)2021-03-09 06:10:20
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
中国调味品(2017年2期)2017-03-20 16:18:25
创新作文(小学版)(2016年16期)2016-11-11 05:47:54
现代检验医学杂志(2016年5期)2016-08-20 03:17:04
电源技术(2016年9期)2016-02-27 09:05:25
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
中国科技信息(2015年2期)2015-11-16 08:18:32
