
微生物合成腺苷蛋氨酸的研究进展
2017-02-21 08:59赵伟军黄磊徐志南浙江大学生物化学工程学院杭州310027
生物技术通报 2017年1期
赵伟军 黄磊 徐志南(浙江大学生物化学工程学院,杭州 310027)
微生物合成腺苷蛋氨酸的研究进展
赵伟军 黄磊 徐志南
(浙江大学生物化学工程学院,杭州 310027)
腺苷蛋氨酸(SAM)是一种存在于所有细胞中的重要代谢产物,对肝病、抑郁症等疾病具有良好疗效,并且无任何副作用,市场需求巨大。目前,国内外学者对SAM的生物合成进行了广泛的研究,一方面在传统发酵法工业化生产SAM的基础上进一步改进发酵策略;另一方面通过代谢工程优化宿主菌代谢网络或者利用基因电路等巧妙筛选方法来进行理性化育种。综述了微生物高效生产SAM的研究进展,讨论了微生物高效生产SAM的意义及存在的问题,旨为提高SAM工业化生产效率提供指导性意义。
腺苷蛋氨酸;酵母;基因电路;发酵;代谢改造
根据中国疾病预防控制中心的统计数字,2015年中国有9 000万乙肝病毒携带者,其中2 800万慢性乙肝患者,760万丙肝感染者,国人保肝护肝严峻形势迫在眉睫。腺苷蛋氨酸(S-Adenosyl-L-methionine,SAM)作为护肝利胆药中唯一注射用药,即注射用丁二磺酸腺苷蛋氨酸,疗效显著,并且无任何副作用,深受患者青睐。同时,SAM对抑郁症和关节炎也有不错的治疗效果,其市场需求巨大。
腺苷蛋氨酸是普遍存在于所有生物细胞的重要代谢产物。它参与细胞的转甲基反应、转硫基反应及转氨丙基反应,在胞内重要分子的甲基化修饰、硫基储存和转移;以及精氨和亚精氨合成起到了不可替代的作用[1]。
SAM的生产方法有化学合成法、发酵法和酶促转化法。其中化学法合成SAM副产物多,分离纯化困难,工业化前景小。而酶促法因为外源底物ATP价格昂贵、SAM合成酶的制备成本较高等特点而难以实现工业化。发酵法是通过发酵培养微生物,控制发酵条件促使微生物细胞积累SAM并通过提取获得大量SAM的过程,是目前工业化生产SAM的主要途径。本文从高产SAM菌株的理性化筛选育种、微生物发酵过程优化以及代谢工程改造3个方向归纳近年来生产腺苷蛋氨酸的研究进展,总结了目前利用微生物高效生产SAM的研究中存在的一些问题,并指出未来发展方向,旨为SAM的工业化生产提供指导性的建议。
1 高产SAM菌株的理性化筛选育种
虽然SAM作为初级代谢产物广泛存在于所有生物细胞中,但是各生物细胞的SAM含量差异却很大。经过研究比较(表1)发现,在环境中含有丰富L-蛋氨酸的条件下,酵母细胞转化合成并积累SAM的能力最强。主要因为酵母细胞含有巨大的液泡,液泡中富含带负电荷的聚磷酸盐,能吸附并固定住带正电荷的SAM[2]。因此,工业生产中一般都选择酿酒酵母和毕赤酵母作为SAM的生产菌株。但是,酿酒酵母和毕赤酵母不同菌株之间的SAM积累能力也有很大差别,如已报导的酿酒酵母菌株中,Shiozaki等[3]就筛选得到一株Saccharomyces sake k-6,能在10 L发酵罐中培养7 d后得到10.8 g/L的SAM产量;而我们常用的BY4741以及BY4742菌株只有0.1 g/L左右的SAM产量。如果通过HPLC检测SAM的方法来筛选高产SAM菌株,工作量将非常大。然而,近年来国内外学者研究报导了几种巧妙地筛选高产SAM菌株的方法。
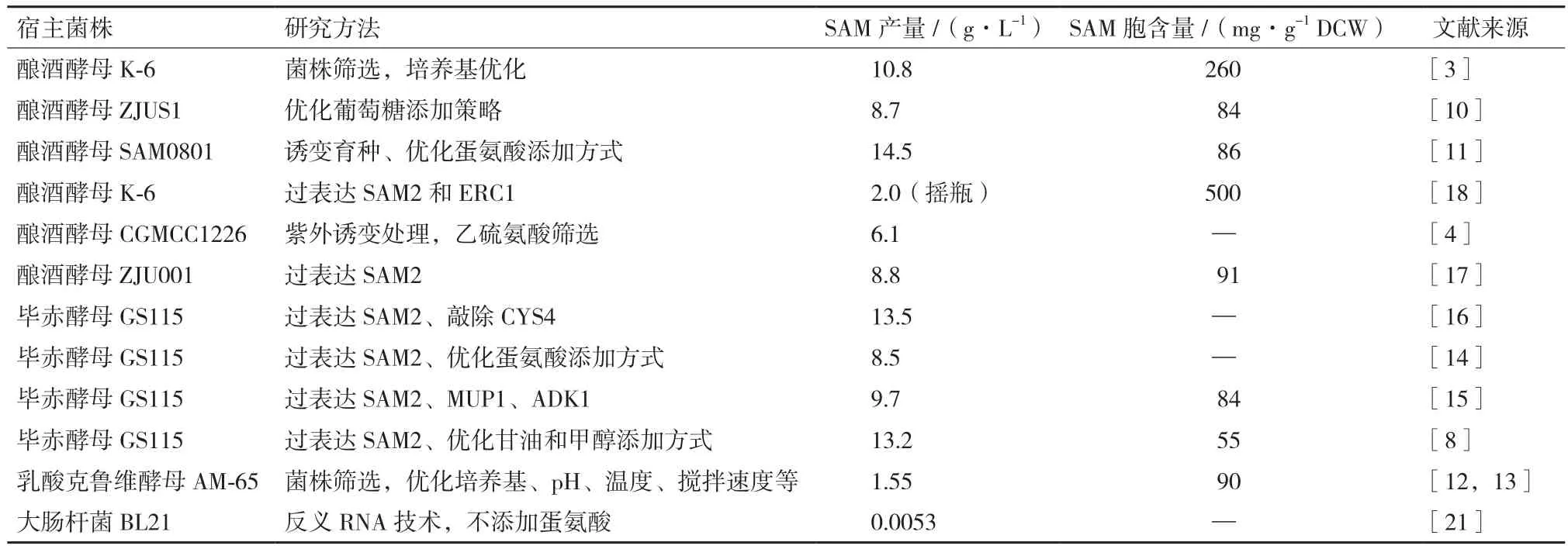
表1 国内外腺苷蛋氨酸产量的研究报道
1.1 基于乙硫氨酸的筛选方法
乙硫氨酸(ethionine)是蛋氨酸类似物,一定浓度下对细胞有毒害作用。有研究发现,酿酒酵母胞内含有抗乙硫氨酸的基因,提高抗乙硫氨酸基因的表达可促进细胞高浓度乙硫氨酸的耐受性,同时酵母细胞内的腺苷蛋氨酸含量也会提高。根据这个规律,Cao等[4]设计了一个利用高浓度乙硫氨酸为筛选条件来筛选高产SAM菌株的实验方案。酿酒酵母菌株CGMCC 2842经过紫外诱变和高浓度乙硫氨酸抗性筛选后得到了SAM产量提升2.7倍的突变菌株,在15 L发酵罐上经过补料分批培养,SAM产量可达到6.1 g/L。
1.2 基于高浓度蛋氨酸的筛选方法
虽然蛋氨酸是合成腺苷蛋氨酸的底物,但如果环境中的蛋氨酸浓度过高,同样会对细胞生长有抑制作用。Kanai等[5]假设能抵抗高浓度蛋氨酸的酿酒酵母菌株也能将蛋氨酸高效合成为腺苷蛋氨酸,并囤积在液泡中。基于这个假设,他们利用高浓度蛋氨酸做为初步筛选条件,从BY4742的含有4 828个单基因突变的单倍体菌种库中筛选到123株抗高浓度蛋氨酸菌株。经过SAM浓度分析检测,其中有81株的SAM浓度有提高,并发现了ADO1这个基因的突变失活能加强蛋氨酸合成途径,从而加强SAM的积累。
1.3 基于制霉菌素的筛选方法
麦角固醇是酿酒酵母等微生物细胞膜的重要组成部分,对确保细胞膜的完整性、膜结合酶的活性、膜的流动性、细胞活力及细胞物质运输等起着重要作用,麦角固醇的体内合成需要消耗大量的SAM来作为甲基供体。制霉菌素(Nystatin)可与真菌细胞膜上的麦角固醇相结合,导致细胞膜通透性发生改变,以致细胞内容物漏失而发挥抗真菌作用。Shobayashi等[6]利用制霉菌素筛选麦角固醇途径缺失的诱变菌株,因为理论上麦角固醇途径缺失,SAM消耗量减少,SAM积累量增多。他们也的确从S. cerevisiae K-9和X2180-1A的诱变菌株中筛选到SAM积累量提高了1.7-5.5倍的菌株,并且通过基因对比发现erg4基因的突变失活对SAM积累量的提升非常关键。
1.4 基于基因电路的高通量筛选方法
在生物细胞内通过引入基因表达与调控元部件构建的代谢调控回路,可实时根据外界信号给出相应反应或指示,这便是基因电路的生物传感器。作为新兴合成生物学的热门之一,基因电路被用于环境监测、快速诊断、理性化育种等领域的研究。其中,作为高通量筛选手段来理性化育种是基因电路一个非常重要的实际应用,Umeyama等[7]就在酿酒酵母YTU225细胞内构建了感应胞内SAM浓度的基因电路,其中经典的能灵敏感应SAM浓度的与门电路,如图1所示。来自大肠杆菌的metJ与B42的复合基因是电路的感受元件,该基因表达的嵌合蛋白能特异地与胞内SAM结合形成复合物,复合物能控制效应元件——荧光蛋白基因Venus的表达。为了能调节电路感受信号阈值——感受SAM浓度范围,Umeyama在效应元件中加入能与抗四环素结合的操纵子(tetO)以及引入抗四环素基因tetR的表达框,利用强力霉素DOX能与抗四环素结合这一特性,便将电路改装成了由SAM和DOX浓度共同作用的与门电路。通过调节外源加入的DOX浓度就能控制电路感受的SAM浓度阈值,以便于控制荧光蛋白表达水平来检测和分析。经过进一步地调试,他们利用这一个基因电路,从构建的随机基因过表达菌种库中筛选到SAM积累加强菌株,并确认GAL11是SAM大量积累的加强基因。尽管GAL11调控SAM合成与积累的机理有待进一步研究和阐明,但是利用基因电路实现了高通量筛选SAM高产菌株,使SAM菌株的理性化育种向前迈进了一大步。
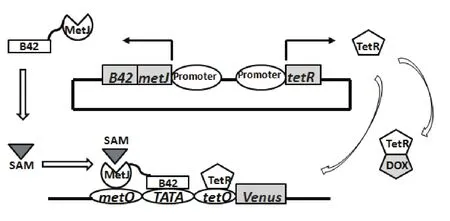
图1 高通量筛选SAM高产菌株的与门基因电路
2 微生物发酵过程优化提高SAM产率的研究
SAM是胞内代谢产物,发酵培养获得的细胞浓度直接关系到SAM产量,高密度发酵培养可以大大提高SAM的体积产量,研究者们在通过优化发酵条件提升SAM产量上做了很多有意义的工作。
2.1 高密度培养毕赤酵母发酵条件的优化
因为毕赤酵母拥有强大的外源基因表达体系以及成熟的高密度发酵技术,学者们也做了很多优化重组毕赤酵母发酵培养条件来生产SAM的研究,其中Hu等[8]在发酵重组SAM合成酶的毕赤酵母时采用的流加策略具有指导意义。他们将发酵培养的诱导时期分为数个间隔,每个间隔为10 h,期间前6 h流加甲醇,后4 h流加甘油。相比传统发酵,这种新型的流加模式一方面保持了甲醇的诱导,使胞内SAM合成酶大量合成;另一方面甘油的流加使菌体内ATP含量充沛,满足SAM合成的底物需求。通过这一发酵策略实现了13.24 g/L的SAM产量,比原来的9.86 g/L提高了34.3%。而Hu等[9]比较了L-蛋氨酸的不同添加方式对发酵重组酵母时生产SAM的影响。研究发现,恒速流加速率过低[0.1 g/(L·h)],作为底物的L-蛋氨酸不足,SAM合成减少;而流加速率过高[0.5 g/(L·h)],毕赤酵母的三羧酸循环和糖酵解途径均受抑制,另一底物ATP的合成减少,SAM的合成也减少了。只有适中速率恒速流加L-蛋氨酸时[流加速率为0.2 g/(L·h)],SAM产量最高,可达8.46 g/L。
2.2 高密度培养酿酒酵母发酵条件的优化
酿酒酵母容易在培养基中葡萄糖浓度过高时产生大量的乙醇,一方面乙醇的积累对酿酒酵母的生长有抑制作用,并且不利于SAM的合成;另一方面使得细胞对碳源的得率系数很低,造成原料的浪费。因而在高密度发酵培养酿酒酵母的时候,尤其要注意控制好葡萄糖等碳源的流加。Lin 等[10]在高密度发酵培养酿酒酵母共生产SAM和谷胱甘肽时,在对数生长期根据指数流加模型设定了不同比生长速率来调控葡萄糖的流加。研究发现,比生长速率μ=0.13 h-1时,发酵液中乙醇含量控制在较低水平,而生物量增长控制在较高速度,最终得到了105 g/L的细胞干重、8.77 g/L的SAM产量和0.81 g/L的GSH产量。国内王杰鹏、谭天伟[11]考察了6种L-蛋氨酸补加策略对酿酒酵母高密度发酵生产SAM的影响,并最终确定了蛋氨酸的加入时机为30 h左右,当菌体干重达到100 g/L时,补加量为每罐40 g蛋氨酸(5 L发酵罐)。最终,发酵58 h左右达到最高生物量干重168 g/L,SAM产量达到14.48 g/L,为目前国内报导最高水平。
2.3 乳酸克鲁维酵母发酵条件的优化
Mincheva等[12,13]筛选到一株积累SAM的乳酸克鲁维酵母,他们先优化了培养基组分,并确定在低蛋白乳清培养基基础上硫酸铵添加量为3.1 g/L、玉米浸出液为12.7 g/L、L-蛋氨酸为4.6 g/L时,SAM的胞含量最高,可达90 mg/g DCW。随后,他们又对批次发酵过程中温度、pH以及搅拌速度进行了优化,并确定当温度为28.5℃、pH为5.3、搅拌速度为270 r/min的时候,SAM的产量最高,可达1.55 g/L。这是为数不多的没有利用酿酒酵母或毕赤酵母来生产SAM的报道,他们的工作为SAM的生产开辟了新的方向和思路。
3 代谢工程改造菌种提高SAM发酵产率的研究
近年来,随着基因工程、代谢工程的蓬勃发展,在酵母、大肠杆菌等遗传背景清楚的宿主菌中,经过代谢网络分析、主观改造宿主代谢途径来提高菌株工业生产性能的方式已经逐步取代传统的诱变育种。酵母中与SAM合成相关的代谢网络如图2所示,本文报导的代谢途径改造来研究SAM生产所涉及的基因均以红色标注。
3.1 毕赤酵母的改造
SAM合成酶可在胞内催化L-蛋氨酸和ATP生成SAM,在酵母体内强化表达SAM合成酶可大幅度提高胞内SAM含量,国内外学者对此进行了大量的研究。SAM合成酶来源很多,但来自酿酒酵母的SAM2因为没有产物抑制效应而被广泛克隆使用。Hu等[14]采用DNA Shuffling技术,将来自大肠杆菌、酿酒酵母、链霉菌3种微生物的SAM合成酶进行DNA重排,构建重排文库。通过转化到毕赤酵母中来比较SAM合成酶活力,他们筛选到一株SAM合成酶酶活提高201%、SAM产量提高103%的重组菌株。经过500 L发酵罐发酵测试,SAM生产力可达到6.14 g/L。Kant 等[15]将SAM合成酶基因SAM2、蛋氨酸通透酶基因MUP1以及腺苷激酶ADK1在毕赤酵母中过表达,探究对SAM胞内合成的影响。研究结果表明,3个基因同时在同一菌株内过表达时,三者的协同效应极大提高了宿主菌的SAM积累能力,比单独过表达SAM2的菌株都提高了77%。最终经过发酵条件优化,3个基因都过表达的菌株的SAM产量达到了9.73 g/L。此外,He等[16]在毕赤酵母中过量表达SAM2的同时还敲除了胱硫醚-β合成酶基因(CYS4),该基因的敲除切断了高半胱氨酸流向合成半胱氨酸的通路,从而使得更多高半胱氨酸流向蛋氨酸合成通路。他们发现这两个代谢途径的改造对SAM的积累有协同作用,SAM产量比原始菌株提高了100多倍,最终在5 L发酵罐中产量达到13.5 g/L。
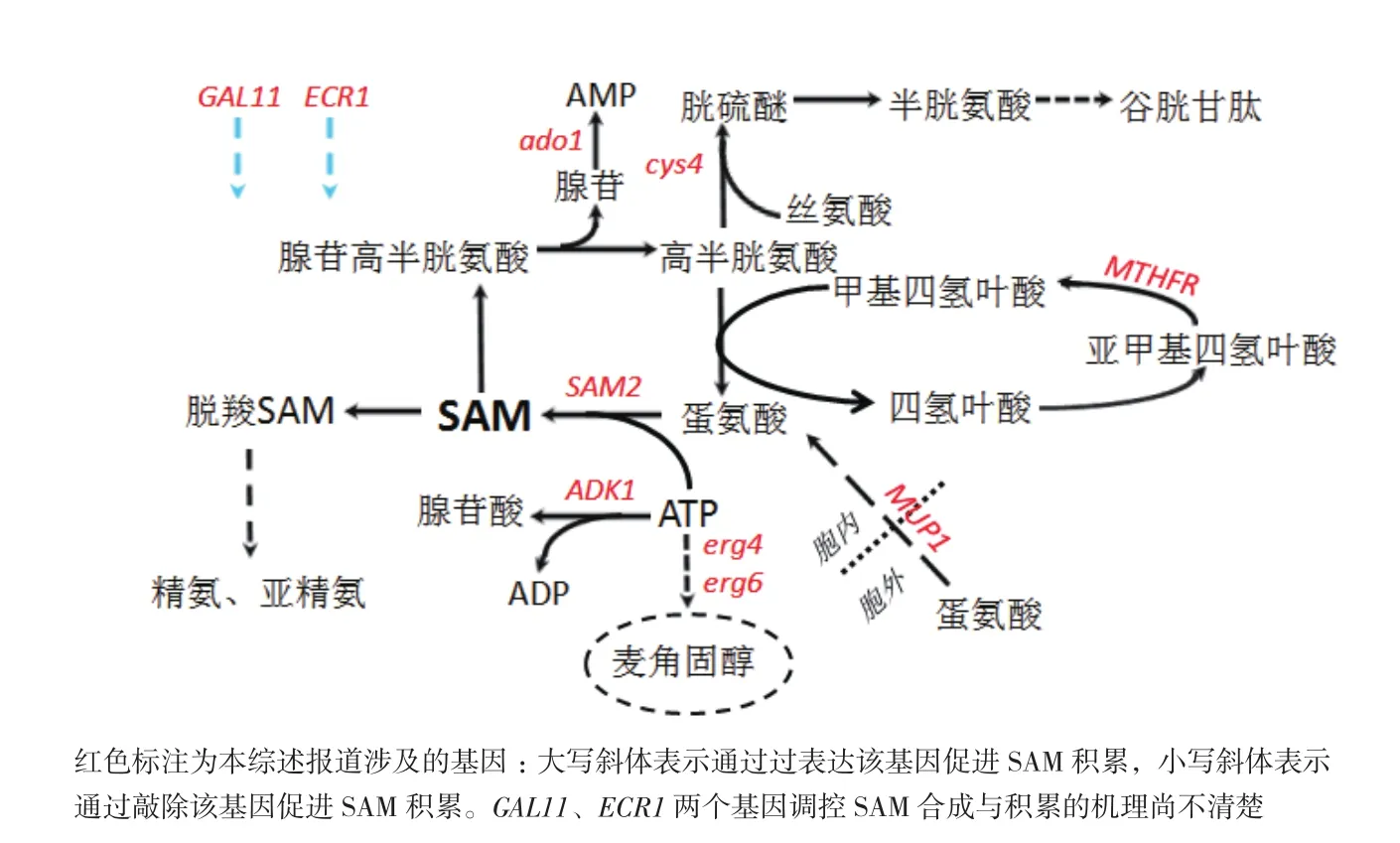
图2 与SAM合成相关代谢网络图
3.2 酿酒酵母的改造
Zhao等[17]利用rDNA位点在基因组上的重复性,将SAM2多拷贝整合在高产SAM的酿酒酵母工业菌株基因组上,获得的重组菌株SAM产量高达8.81 g/L,比原始菌株提高了27.1%。Lee等[18]则在酿酒酵母中强化表达SAM2的同时过量表达ERC1基因,他们因此得到的代谢改造菌株的SAM胞含量竟高达细胞干重的一半,而SAM的摇瓶产量也达到2 g/L。
在酿酒酵母细胞内,5-甲基-四氢叶酸还原酶(MTHFR)是L-蛋氨酸合成途径的关键酶,它是NADPH依赖型蛋白,活性却受到SAM的反馈抑制。而拟南芥植物的MTHFR是NADP依赖型,却对SAM不敏感,不受SAM的反馈抑制。Roje等[19]将酿酒酵母MTHFR的C端用拟南芥植物的MTHFR置换,而N端保持原来序列,这样生成的嵌合体一方面保持了NADPH依赖型;另一方面消除SAM的反馈抑制。嵌合体蛋白在酿酒酵母中表达后,宿主菌保内蛋氨酸含量提高了7倍,而SAM含量却提高了140倍。虽然当时他们做这个工作只是为了探究酿酒酵母胞内一碳代谢网络,但是其研究结果却给工业化生产SAM提供了新的视角。如果能利用代谢工程加强菌株胞内L-蛋氨酸的合成,减少甚至省去生产过程中外源L-蛋氨酸的添加,那将为SAM的大规模工业化生产节省不小的成本。
3.3 大肠杆菌的改造
大肠杆菌积累SAM的能力不强,但因为大肠杆菌基因工程技术成熟、容易培养,用作研究SAM代谢调控的模式菌株也是非常有意义的。NADPH是细胞内合成L-蛋氨酸的关键因子,而ATP是细胞合成SAM的底物,Chen等[20,21]在大肠杆菌中利用反义RNA技术先后调控了大肠杆菌胞内NADPH以及ATP含量,探究了NADPH以及ATP胞内含量对SAM合成的影响。研究发现,胞内NADPH含量以及ATP含量的增加都能提升大肠杆菌的SAM合成能力(表1)。虽然他们构建的重组大肠杆菌SAM产量还很低,但他们利用的反义RNA干扰技术,以及对NADPH、ATP调控策略都能为今后SAM生产的研究工作提供很好的借鉴。
4 展望
随着研究的深入,SAM的更多医疗功效被发现,由于其无任何副作用的特性,获得广大肝病、抑郁症患者的青睐,市场前景十分广阔。虽然生产SAM的研究已经取得了很多进展,但是微生物强大的SAM积累能力并未充分发掘,SAM的工业化生产还存在一些问题。首先,已报道的上罐发酵SAM产量都不高,酿酒酵母以及毕赤酵母最高报道产量都在15 g/L以下;并且外源蛋氨酸的加入,进一步提高了SAM工业化生产的成本。其次,胞内调控SAM合成与积累的机制还不是很明朗,最近研究报道的改造菌株代谢网络来提高SAM产量的方法也都只是在某一到两个点上的摸索,缺乏整体性的研究。比如,GAL11、ECR1两个基因对SAM积累的促进作用已被证实,但它们促进SAM积累的机理却并不清楚。再次,菌株理性化育种中涉及的很多筛选方法有比较大的局限性,虽然与门基因电路在酿酒酵母YTU225中实现了高通量筛选,但其在工业化生产中应用和推广还有待进一步研究与考证。在未来研究微生物高效合成SAM时,我们应加强胞内SAM合成与积累调控机制的研究,进一步利用代谢工程优化宿主菌生产SAM的代谢网络;开拓基因电路等高通量筛选方法,实现工业菌株理性化育种;与此同时,结合新型微生物发酵技术充分发掘其SAM生产力。在注重代谢工程优化宿主菌生产SAM的代谢网络时,一方面我们可以从提升宿主菌积累SAM能力角度出发;另一方面我们还可以从提高碳源、蛋氨酸利用率、节省生产成本的角度出发。此外,探索新的SAM生产宿主菌或者发现新的SAM合成与积累途径也是值得拓展研究的。相信随着认识的深入和技术的发展,利用微生物工业化生产SAM的生产效率还会取得更进一步的提高。
[1]Lu SC. S-adenosylmethionine[J]. International Journal of Biochemistry and Cell Biology, 2000, 32:391-395.
[2]Chan SY, Appling DR. Regulation of S-adenosylmethionine levels in Saccharomyces cerevisiae[J]. Journal of Biological Chemistry, 2003, 278(44):43051-43059.
[3]Shiozaki S, Shimizu S, Yamada H. Production of S-adenosyl-L-methionine by Saccharomyces sake[J]. Journal of Biotechnology, 1986, 4(6):345-354.
[4]Cao XT, Yang MH, Xia Y. Strain improvement for enhanced production of S-adenosyl-L-methionine in Saccharomyces cerevisiae based on ethionine-resistance and SAM synthetase activity[J]. Annals of Microbiology, 2012, 62:1395-1402.
[5]Kanai M, Masuda M, Takaoka Y, et al. Adenosine kinasedeficient mutant of Saccharomyces cerevisiae accumulates S-adenosylmethionine because of an enhanced methionine biosynthesis pathway[J]. Applied Microbiology and Biotechnology, 2013, 97:1183-1190.
[6]Shobayashi M, Mukai N, Iwashita K, et al. A new method for isolation of S-adenosylmethionine(SAM)-accumulating yeast[J]. Applied Microbiology and Biotechnology, 2006, 69:704-710.
[7]Umeyama T, Okada S, Ito T. Synthetic gene circuit-mediated monitoring of endogenous metabolites:identification of GAL11 as a novel multicopy enhancer of S-adenosylmethionine level in yeast[J]. ACS Synthetic Biology, 2013, 2:425-430.
[8] Hu XQ, Chu J, Zhang SL, et al. A novel feeding strategy during the production phase for enhancing the enzymatic synthesis of S-adenosyl-L-methionine by methylotrophic Pichia pastoris[J]. Enzyme and Microbial Technology, 2007, 40:669-673.
[9]Hu H, Qian JC, Chu J, et al. Optimization of l-methionine feeding strategy for improving S-adenosyl-l-methionine production by methionine adenosyltransferase overexpressed Pichia pastoris[J]. Applied Microbiology and Biotechnology, 2009, 83:1105-1114.
[10]Lin JP, Tian J, You JF, et al. An effective strategy for the coproduction of S-adenosyl-L-methionine and glutathione by fedbatch fermentation[J]. Biochemical Engineering Journal, 2004, 21:19-25.
[11]王杰鹏, 谭天伟. 发酵法生产S-腺苷蛋氨酸前体蛋氨酸补加策略[J]. 生物工程学报, 2008, 24(10):1824-1827.
[12]Mincheva K, Kamberova V, Balutzov V. Production of S-adenosyl-L-methionine by a mutant strain of Kluyveromyces lactis[J]. Biotechnology Letters, 2002, 24:985-988.
[13]Mincheva K, Kamberova V, Balutzov V. Optimization of S-adenosyl-L-methionine production by Kluyveromyces lactis on whey in batch culture using a mathematical model[J]. Biotechnology Letters, 2002, 24:1773-1777.
[14]Hu H, Qian JC, Chu J, et al. DNA shuffling of methionine adenosyltransferase gene leads to improved S-adenosyl-L-methionine production in Pichia pastoris[J]. Journal of Biotechnology, 2009, 141:97-103.
[15]Kant HR, Balamurali M, Meenakshisundaram S. Enhancing precursors availability in Pichia pastoris for the overproductionof S-adenosyl-l-methionine employing molecular strategies with process tuning[J]. Journal of Biotechnology, 2014, 188:112-121.
[16]He JY, Deng JJ, Zheng YH, Gu J. A synergistic effect on the production of S-adenosyl-L-methionine in Pichia pastoris by knocking in of S-adenosyl-L-methionine synthase and knocking out of cystathionine-β synthase[J]. Journal of Biotechnology, 2006, 126:519-527.
[17]Zhao WJ, Shi F, Hang BJ, et al. The improvement of SAM accumulation by integrating the endogenous methionine adenosyltransferase gene SAM2 in genome of the industrial Saccharomyces cerevisiae strain[J]. Applied Biochemistry and Biotechnology, 2016, 178(6):1263-1272.
[18]Lee SW, Park BS, Choi ES, et al. Overexpression of ethionine resistance gene for maximized production of S-andenosylmethionine in Saccharomyces cerevisiae sake kyokai No. 6[J]. Korean Journal of Chemical Engineering, 2010, 27(2):587-589.
[19]Roje S, Chan SY, Kaplan F, et al. Metabolic engineering in yeast demonstrates that S-adenosylmethionine controls flux through the methylenetetrahydrofolate reductase reaction in vivo[J]. Journal of Biological Chemistry, 2002, 227(6):4056-4061.
[20]Chen YW, Xu DB, Fan LH, et al. Manipulating multisystem of NADPH regulation in Escherichia coli for enhanced S-adenosylmethionine production[J]. RSC Advances, 2015, 5:41103-41111.
[21] Chen YW, Lou SY, Fan LH, et al. Control of ATP concentration in Escherichia coli using synthetic small regulatory RNAs for enhanced S-adenosylmethionine production[J]. FEMS Microbiology Letters, 2015, 362(15):1-8
(责任编辑 狄艳红)
Research Progress on the Synthesis of S-adenosyl-L-methionine in Microorganism
ZHAO Wei-jun HUANG Lei XU Zhi-nan
(College of Chemical and Biological Engineering,Zhejiang University,Hangzhou 310027)
S-adenosyl-L-methionine(SAM)as an important metabolite exists in all cell types of living organisms,presents promising effects on liver diseases and depressive syndromes without any side effects,thus,it is greatly demanded in medical market. Recently,broad studies on biosynthesis of SAM have been conducted by scholars in the world. On the one hand,fermentation strategies based on traditionally producing SAM were further improved;on the other hand,different rational breeding programs were developed,such as fine tuning host’s metabolic network or screening strains by synthetic gene circuit. In this paper,progress in the research of S-adenosyl-L-methionine production is summarized,and the significances and issues while efficiently producing SAM by microorganism are discussed,aiming at providing the guides for improving SAM productivity at an industrial scale.
S-adenosyl-L-methionine;yeast;synthetic gene circuit;fermentation;metabolic engineering
10.13560/j.cnki.biotech.bull.1985.2017.01.010
2016-05-03
赵伟军,男,博士,研究方向:酿酒酵母代谢工程、发酵工程;E-mail:zwj57757743@zju.edu.cn
猜你喜欢
中国饲料(2022年5期)2022-04-26
检察风云(2022年5期)2022-04-05
传染病信息(2021年6期)2021-02-12
中成药(2017年12期)2018-01-19
福建轻纺(2017年12期)2017-04-10
广东饲料(2016年3期)2016-12-01
国外医药(抗生素分册)(2016年3期)2016-07-12
山东医药(2015年16期)2016-01-12
现代农业(2015年5期)2015-02-28
中国畜牧业(2014年4期)2014-10-16
