
盐旱交叉胁迫对小麦幼苗渗透调节能力的影响
2017-02-20 02:55华智锐李小玲
山西农业科学 2017年2期
华智锐,李小玲
(商洛学院生物医药与食品工程学院,陕西商洛726000)
盐旱交叉胁迫对小麦幼苗渗透调节能力的影响
华智锐,李小玲
(商洛学院生物医药与食品工程学院,陕西商洛726000)
以商麦5226种子为试验材料,采取模拟干旱和盐分交叉胁迫小麦幼苗的方法,通过测定交叉胁迫下小麦种子的发芽率、发芽势、可溶性糖含量、丙二醛(MDA)含量、可溶性蛋白含量等指标,以探讨小麦随盐旱胁迫时间变化渗透调节物质的变化规律,为培育抗盐抗旱的优良小麦品种提供理论基础。结果表明,小麦种子的发芽率、发芽势随着盐旱胁迫程度的增加呈逐渐下降的趋势;在0.6%盐溶液和10%的聚乙二醇交叉胁迫条件下,小麦幼苗具有一定的交叉适应性,发芽率为83%,发芽势为96%,可溶性糖在第2天达到最大值(2.56 mg/g),与对照相比增加了14.29%,降低了细胞水势,维持细胞正常生长代谢;丙二醛缓慢上升再下降,消除活性氧对植物的破坏,提高植物的抗氧化能力;可溶性蛋白在第2天达到最大值(0.34 mg/g),并分解成各种氨基酸,尤其是脯氨酸的含量增多,降低水势,减轻盐旱所造成的伤害。
小麦;盐旱交叉胁迫;渗透调节
小麦是小麦属植物的统称,具有较高的营养价值和食疗作用,它含有蛋白分解酶及多种维生素等成分,可增强人体的抗病能力,是一种在世界各地广泛种植的禾本科植物,起源于中东地区。其种植面积、分布范围居世界首位,是我国主要的粮食作物。商洛市位于秦岭东段南麓,地形构造复杂,气候变化多样,而小麦在商洛的粮食生产中占据主导地位,旱、寒、薄、病是商洛小麦生产的四大胁迫因子。由于其生态环境、气候适宜于优质中筋小麦的生产。因此,选育抗逆性强、品质好、适应性广的小麦品种显得尤为重要。商麦5226正是针对商洛旱地小麦的生产情况而育成的一种质量优、产量高的新型品种,具有抗旱能力强的特点[1]。
环境因子在植物生长发育过程中有非常重要的影响作用,其中,盐分和水分属于极其重要的影响因子[2-5]。近年来,由于人口数量的不断增加,小麦的需求量呈上涨趋势,但耕地面积减少,盐渍、干旱现象日渐凸显,使得农作物受到严重威胁,农作物产量明显降低,并且造成土壤溶液的水势不断下降,植物的水分吸收受到阻碍,细胞缺水,严重时造成植物组织内水分外渗,引起死亡。随着盐旱程度的不断加重,有关植物抗盐旱的研究更加引起人们重视。杨颖丽等[6]对NaCl胁迫下2种春小麦新品种渗透调节反应的比较进行研究,结果显示,盐胁迫诱导2种小麦叶片可溶性糖含量升高。GONG等[7]曾报道,通过干旱锻炼可以提高水稻的抗寒性。有研究表明,干旱胁迫不仅能够抑制蛋白质合成,还能诱导蛋白质的降解,从而使植株体内的总蛋白质含量降低[8]。王兰兰等[9]围绕小麦幼苗对盐胁迫和水分胁迫的生理反应对比进行研究,结果表明,2种渗透胁迫下,与对照相比,小麦幼苗的MDA含量显著升高,可溶性糖含量普遍高于对照。向梅梅等[10]就盐旱交叉胁迫对玉米幼苗渗透调节物质的影响进行研究,结果表明,可溶性糖和丙二醛含量均随盐旱交叉胁迫程度的加重而增加,可溶性蛋白质含量随盐旱交叉胁迫程度的加重呈先上升后下降的趋势。
植物在受到逆境胁迫时,渗透调节是重要的调节途径。盐和干旱是影响植物生长的主要环境因素,盐旱交叉胁迫又称为渗透胁迫,二者都因导致土壤的水势降低使细胞缺水,引起死亡。在逆境胁迫条件下,参与渗透调节的物质可分为2类:其一是由外界进入植物细胞内的无机离子;其二是在细胞内自身合成的有机溶质[2]。
目前,关于盐胁迫和干旱胁迫对小麦幼苗的渗透调节能力影响主要集中在单一因素的盐分或干旱胁迫,而以盐旱交叉胁迫对小麦渗透调节能力影响的研究尚未见报道。但是,在盐旱交叉胁迫对植物的影响研究方面,前人对柽柳[11]、玉米[10]、燕麦[12]、皂角[13]、君迁子[14]、紫荆[15]、银砂槐[16]、紫穗槐[17]等幼苗进行了研究。
本研究在不同的盐浓度与不同干旱处理交叉胁迫下,分析探讨盐旱交叉胁迫下小麦幼苗随盐旱胁迫时间变化渗透调节物质的变化规律,以获得适宜小麦生长的土壤干旱度和盐碱度,为培育抗盐抗旱的优良品种提供理论基础。
1 材料和方法
1.1 材料及仪器试剂
1.1.1 试验材料商麦5226种子由商洛学院秦岭植物良种繁育中心提供。
1.1.2 主要试剂NaCl、聚乙二醇(PEG-6000)、乙醇、石英砂、10%三氯乙酸(TCA)、0.6%硫代巴比妥酸(TBA)、蒽酮、葡萄糖、硫酸、牛血清蛋白、考马斯亮蓝G-250、磷酸、蒸馏水。
1.1.3 主要仪器U755B紫外可见光分光光度计、电磁炉、恒温水浴锅、冰箱、XBJ-20制冰机、TP-412电子天平、H2050R台式高速冷冻离心机、恒温培养箱。
12试验方法
1.2.1 试剂的配制准确称取三氯乙酸50.00 g,定容至500 mL,即为10%TCA;称取硫代巴比妥酸0.6 g,先用1 mol/LNaOH溶解,再用10%TCA定容至100 mL,即为0.6%TBA;称取考马斯亮蓝G-250 100 mg,溶于50 mL 90%乙醇中,再加入85%H3PO4100 mL,再用蒸馏水定容至1 000 mL,即为考马斯亮蓝G-250溶液(贮存在棕色瓶中,常温下可放置30 d)。
蒽酮试剂:称取0.1 g蒽酮溶于100 mL稀硫酸(由76 mL相对密度1.84的硫酸稀释成100 mL)中,贮于棕色瓶内,冰箱保存,可用数日。
1.2.2 小麦种子的处理挑选籽粒饱满、种皮光亮的商麦5226种子为试验材料,先用0.1%HgCl2消毒15 min,然后用无菌水漂洗3~5次。蒸馏水浸种24 h后,将灭菌好的种子放入铺有2层滤纸的直径为11 cm培养皿中,然后用镊子在每个培养皿放入50粒的小麦种子,摆放整齐后,盖上皿盖,置于25℃培养箱进行培养,以便催芽。期间来回翻动种子,注意湿度的保持,72 h后将种子移至培养箱外,自然状态下培养[18]。
与上述方法相同,用镊子在10个铺有2层滤纸的直径为11 cm培养皿放入30粒的小麦种子,摆放整齐后,对其进行胁迫处理,盖上皿盖,置于25℃培养箱进行培养,72 h后将种子移至培养箱外,自然状态下培养。每天定时加入相应的盐、PEG,以维持恒定的浓度。在发芽的第1天开始每天统计发芽数,直至发芽结束,测其7 d后的发芽率和前3 d的发芽势。

1.2.3 小麦幼苗的培养及交叉胁迫处理将小麦种子放入培养皿进行培养(蒸馏水培养),每天定时补充蒸馏水,促使种子萌发,待萌发后小麦生长至两叶一心时,对其进行交叉胁迫处理。取浓度为0.3%(轻度盐胁迫)、0.6%(中度盐胁迫)、0.9%(重度盐胁迫)的盐溶液分别与10%PEG(轻度干旱胁迫)、15%PEG(中度干旱胁迫)、20%PEG(重度干旱胁迫)交叉结合对小麦种子进行胁迫处理(表1),同时以蒸馏水为对照(CK),每处理3次重复。处理后隔天采样测定相关生理指标。

表1 盐旱交叉胁迫试验设计
1.3 生理指标测定及方法
1.3.1 可溶性糖含量的测定样品的提取:称取剪碎混匀的新鲜样品0.3 g,放入三角瓶(50 mL)水中,再加25 mL蒸馏水,放入沸水浴中煮沸20 min,取出冷却。过滤入100 mL容量瓶,用热水冲洗残渣数次,定容至刻度。
显色反应及测定:取4支试管,吸取样品提取液各0.5 mL,加入蒸馏水0.5 mL和5 mL蒽酮试剂,以蒸馏水为对照,将各管摇匀后,在沸水浴中煮沸10 min,取出冷却,在分光光度计620 nm处比色,记录光密度值[19]。

式中,C为查到待测果实的葡萄糖含量,a为样品提取液体积,v为提取液定容体积,n为稀释倍数,W为样品质量。
1.3.2 可溶性蛋白含量的测定蛋白质的提取:称取小麦叶片0.3 g,放入研钵中,添加少量石英砂和蒸馏水研磨成匀浆后,转入10 mL容量瓶,再用蒸馏水冲洗研钵研锤2~3次,将清洗液合并于10mL容量瓶中,并定容至刻度。取2~3 mL匀浆液于10 mL离心管中,5 000 r/min下离心10 min,上清液即为蛋白质提取液。
显色反应及测定:吸取0.1 mL上述蛋白质提取液,加入蒸馏水0.9 mL和考马斯亮蓝G-250试剂5 mL,充分混合,放置2 min后于分光光度计595 nm下比色,并记录吸光值[20]。

式中,C为查得的标准曲线值,Vt为提取液总体积,Vs为测定时加样量,W为样品鲜质量,1 000为将μg换成mg。
1.3.3 丙二醛(MDA)含量的测定MDA提取:称取剪碎的试材0.3 g,加入10%三氯乙酸(TCA)2 mL和少量石英砂,研磨至匀浆,再加入TCA 8 mL进一步研磨,将其转入离心管,于4 000 r/min下离心10 min,取上清液即为样品提取液。
显色反应及测定:取提取液2 mL(对照加2 mL蒸馏水),加入2 mL 0.6%TBA(硫代巴比妥酸)液,转入试管中,试管加塞,于水浴锅中煮沸15 min,流水中迅速冷却,在4 000 r/min下离心2 min。取上清液测定532,600,450 nm处的吸光度[21]。
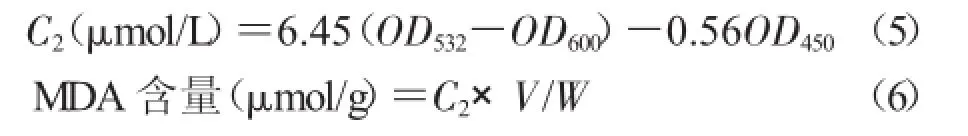
式中,C2为丙二醛的浓度,V为提取液的体积,W为样品的鲜质量。
1.4 数据处理
采用Excel 2007,SPSS 17.0软件进行数据处理和统计分析。
2 结果与分析
2.1 盐旱交叉胁迫对小麦幼苗的发芽率、发芽势的影响

从图1可以看出,与CK相比,小麦种子的发芽率、发芽势整体上均呈现下降的趋势。随着盐旱胁迫程度的加深和胁迫时间的延长,发芽率、发芽势呈现出不同的变化规律,其中,AT1,AT2,BT1的发芽率与对照相比分别提高了30%,17%,17%,而AT1,AT2,BT1的发芽势与对照相比分别提高了23%,20%,23%;在盐浓度为0.3%或0.6%时,发芽率最大,为96%,其次依次是AT2和BT1,BT1发芽势最大,为96%,其次是AT2,表明一定浓度的盐旱胁迫会促进种子的萌发,在盐浓度为0.9%时,其发芽势和发芽率均呈现下降的趋势,说明重度盐旱胁迫会抑制种子的萌发,使根系吸水困难,从而导致生长缓慢。
2.2 盐旱交叉胁迫对小麦幼苗的可溶性糖含量的影响
从图2可以看出,在盐旱交叉胁迫下小麦叶片中可溶性糖含量存在不同差异,变化趋势较为复杂。其中,在AT1,AT2,AT3,BT2,CT3处理下,与对照相比无显著性差异(P>0.05),随着盐旱交叉胁迫时间的延长,可溶性糖含量呈现出先下降后上升的趋势,说明适度的水分提高了可溶性糖含量;而在其余的浓度处理下,存在有显著性差异(P<0.05),其中,BT1差异最为显著(P<0.01)。在浓度为0.6%盐溶液与干旱胁迫下,小麦幼苗可溶性糖含量第2天达到最大值(2.56 mg/g),与对照相比增加了14.29%,其可溶性糖含量呈现先升高后下降的变化趋势。

2.3 盐旱交叉胁迫对小麦幼苗的可溶性蛋白含量的影响

从图3可以看出,在盐旱交叉胁迫下小麦叶片中可溶性蛋白含量差异明显,变化趋势较为复杂。其中,AT1,AT2,AT3处理下,与对照相比无显著性差异(P>0.05);随着时间延长,呈现先下降后上升的趋势,说明在抗逆境中可溶性蛋白起到了作用。而其余各处理与对照相比存在显著性差异(P<0.05),其中,BT1的差异最为明显(P<0.01),随着胁迫时间的延长和程度的加深,在第2天时,BT1处理下的可溶性蛋白含量达到最大值(0.34 mg/g),比对照增加了240%,并呈现先上升后下降的趋势,而在重度胁迫处理下蛋白含量呈现逐渐降低的趋势。
2.4 盐旱交叉胁迫对小麦幼苗的丙二醛含量(MDA)的影响
由图4可知,在盐旱交叉胁迫下小麦叶片中丙二醛(MDA)含量存在不同差异,变化趋势复杂。随着盐旱交叉胁迫程度的增加和胁迫时间的延长,MDA含量总体上呈现上升趋势,其中,在AT3,BT1,BT3,CT1处理下,与对照相比,无显著性差异(P>0.05);而在其他浓度的处理下有显著性差异(P<0.05),丙二醛含量随着胁迫时间和胁迫程度的加重而增加。在浓度为0.6%盐处理下,MDA含量呈现一个缓慢上升的趋势,说明适度的水分增加了小麦在此浓度下的交叉适应性;而浓度为0.9%盐与干旱胁迫处理下,随着时间的延长,丙二醛含量呈现逐渐增大的趋势。

3 结论与讨论
植物在遭受逆境胁迫时,可以通过某些生理生化的变化来缓解不良环境对其造成的影响,从而减弱逆境所带来的危害[22]。渗透调节是通过积累无机离子和小分子有机物来维持植物渗透平衡的调节机制[23]。在适度的盐旱交叉胁迫下,渗透调节物质起到了调节作用,使得植物提高了自身的抗盐旱能力,对外界不良环境表现出一定的交叉适应性,交叉逆境条件下植物渗透调节能力的增强可能是交叉适应的生理机制[24]。
发芽率和发芽势是表示植物种子发芽能力的重要指标,发芽率能反映种子的发芽能力,而发芽势能表征种子的发芽速度和幼苗的生长速度[17]。盐旱胁迫下小麦的发芽率与发芽势呈现逐渐下降的趋势。本研究表明,在浓度为0.6%NaCl与10% PEG处理下,发芽率为83%,发芽势为96%,表明在此浓度下能够促进小麦的萌发,有利于呼吸;而在浓度为0.9%NaCl与不同干旱处理下,会抑制种子的萌发,影响根部吸收水分的能力,难以维持植株正常生长。
可溶性糖是植物体内重要的渗透调节物质[25]。它不仅能够使得细胞膜和原生质胶体处于一个稳定状态,同时还可以作为合成其他有机溶质的碳架和能量来源。可溶性糖含量的增加对增加植物细胞汁液浓度,降低细胞的水势,从而提高植物吸水能力十分有利[26]。本研究表明,可溶性糖含量在AT1,AT2,AT3,BT2,CT3处理下,与对照相比无显著性差异(P>0.05),随着胁迫时间的延长,呈现先下降后上升的趋势,说明适度的水分使得可溶性糖含量增加;而在其他的浓度处理下,存在显著性差异(P<0.05),其中,BT1差异最为显著(P<0.01)。在浓度为0.6%盐溶液与干旱胁迫下,第2天达到最大值(2.56 mg/g),与对照相比增加了14.29%,其可溶性糖含量呈现先升高后下降的趋势。可溶性糖对盐旱胁迫比较敏感,可以作为判定是否具有抗性的一个指标,可溶性糖含量的增加,说明它能够降低细胞渗透势,维持细胞膨压,使细胞内部与外界环境保持稳定状态,保证细胞的正常生长和代谢。
MDA是植物在逆境条件下或器官衰老时,发生膜脂过氧化作用的产物之一,通常表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱[27]。研究表明,随着盐旱交叉胁迫程度的增加和时间的延长,MDA含量基本上呈上升趋势。本研究结果表明,在AT3,BT1,BT3,CT1处理下,MDA与对照相比,无显著性差异(P>0.05),而在其他浓度的处理下有显著性差异(P<0.05),丙二醛含量随着胁迫时间和胁迫程度的加重而增加。在浓度为0.6%盐处理下,呈现一个缓慢上升再下降的趋势,说明适度的水分增加了小麦在此浓度下的交叉适应性,可能是由于保护酶消除了活性氧对植物的损害,导致丙二醛含量降低,缓解了膜脂的过氧化;而在浓度为0.9%盐与干旱胁迫处理下,随胁迫时间的延长,丙二醛含量呈现逐渐升高的趋势,说明在该处理下抗氧化系统被破坏,植物体内的活性氧大量积累,导致膜脂的过氧化加重,对细胞膜造成了比较严重的伤害。
植物体内的可溶性蛋白质多数为参与相关代谢的酶类,其含量是了解和研究植物总代谢的一个重要指标[28]。本研究表明,在AT1,AT2,AT3处理下,可溶性蛋白含量与对照相比无显著性差异(P>0.05),随着时间延长,呈现先下降后上升的趋势,表明可溶性蛋白能够增强小麦的抗性。而其余各处理与对照相比存在显著性差异(P<0.05),其中,BT1的差异最为明显(P<0.01),随着胁迫时间的延长和胁迫程度的加深,在第2天时,BT1处理下的可溶性蛋白含量达到最大值(0.34 mg/g),比对照增加了240%,并呈现先上升后下降的趋势,可能是由于盐旱胁迫下植物体内的蛋白质分解加速,各种氨基酸含量增加,其中脯氨酸含量大量积累,降低了叶片水势,缓解了盐渍和干旱对其的伤害;而在重度胁迫处理下蛋白含量呈现逐渐下降的趋势,说明合成可溶性蛋白受到抑制,降解速度加快,渗透能力也随之下降。
综上所述,适当的盐旱交叉胁迫,能使抗氧化系统增强,提高植物的抗逆境适应能力。本研究使用不同NaCl溶液与不同浓度的PEG交叉处理小麦种子,结果表明,可溶性糖含量增加,细胞渗透势降低,细胞维持了一定的膨压,正常呼吸和各项生理活动得到保证,丙二醛(MAD)含量降低,消除了大量的活性氧,减轻了膜脂的过氧化,可溶性蛋白含量提高,蛋白质分解加速,脯氨酸含量积累,减轻盐害,从而抵抗了胁迫对植物的损害。其中,在0.6%NaCl与10%PEG处理下,对提高小麦的抗逆性效果最好。然而,关于小麦在大田逆境(盐旱交叉)胁迫下抗逆能力及抗逆生理方面还有待于进一步研究。
[1]于浩世,王新军,华智锐,等.抗逆高产优质小麦品种商麦5226的选育[J].干旱地区农业研究,2008,26(3):76-79.
[2]肖春旺,周广胜,马风云.施水量变化对毛乌素沙地优势植物形态与生长的影响[J].植物生态学报,2002,26(1):69-76.
[3]LANGE O L,KAPPEN L,SCHULZE E D.Water and plant life[M].NewYork:Springer ver,1976:207-224.
[4]詹海仙,畅志坚,魏爱丽,等.干旱胁迫对小麦生理指标的影响[J].山西农业科学,2011,39(10):1049-1051.
[5]马富举,李丹丹,蔡剑,等.干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响[J].应用生态学报,2012,23(3):724-730.
[6]杨颖丽,李翠祥,滕玉瑾,等.NaCl胁迫下2种春小麦新品种渗透调节反应的比较[J].西北师范大学学报,2016,52(1):93-99.
[7]GONG M,LIU Y L,ZHU P R.Physiological effects of chilling and drought hardeningon cold resistance ofthe clones ofhybrid japonica rice[J].NanjingAgr Univ,1989,12(1):22-27.
[8]WEI L M.A studyon carbohydrate and protein of several xerophytes [J].Arid Zone Research,1991,8(4):38-41.
[9]王兰兰,刘新,刘贺楠,等.小麦幼苗对盐胁迫和水分胁迫的生理反应对比[J].沈阳师范大学学报,2013,31(1):120-123.
[10]向梅梅,陈江,位杰,等.盐旱交叉胁迫对玉米幼苗渗透调节物质含量的影响[J].新疆农垦科技,2015(5):7-9.
[11]朱金方,陆兆华,夏江宝,等.盐旱交叉胁迫对柽柳幼苗渗透调节物质含量的影响[J].西北植物学报,2013,33(2):357-363.
[12]刘建新,王金成,王瑞娟,等.旱盐交叉胁迫对燕麦幼苗生长和渗透调节物质的影响[J].水土保持学报,2012,26(3):244-248.
[13]孙艳菊.皂角、君迁子和紫荆苗木对盐旱交叉胁迫反应的研究[D].泰安:山东农业大学,2007.
[14]张慎鹏,孙明高,张鹏,等.盐旱交叉胁迫对君迁子渗透调节物质含量的影响[J].西北林学院学报,2008,23(5):18-21.
[15]孙方行.紫荆苗期在盐分干旱及其交叉胁迫下反应的研究[D].泰安:山东农业大学,2006.
[16]庄伟伟,李进,曹满航,等.NaCl与干旱胁迫对银砂槐幼苗渗透调节物质含量的影响[J].西北植物学报,2010,30(10):2010-2015.
[17]赵建诚,秦柱南,孙超,等.紫穗槐种子萌发对盐旱逆境的生理响应[J].种子,2012,31(5):26-29.
[18]祝社民.水杨酸对PEG胁迫下商麦5226幼苗生长的影响[J].商洛学院学报,2014,28(6):65-67.
[19]郝建军,康宗利,于洋,等.植物生理学实验技术[M].北京:化学工业出版社,2006:141-142.
[20]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:141-143.
[21]邹琪.植物生理学实验指导[M].北京:中国农业出版社,2000:173-174.
[22]庄伟伟,李进,曹满航,等.盐旱交叉胁迫对银砂槐幼苗生理生化特性的影响[J].武汉植物学研究,2010,28(6):730-736.
[23]赵萍萍.逆境胁迫下外源调节因子对小麦幼苗生长的影响[D].郑州:河南师范大学,2013.
[24]李洁,武杭菊,胡景江,等.干旱-低温交叉逆境下小麦渗透调节能力的变化与交叉适应的关系[J].干旱地区农业研究,2009,27(6):149-153.
[25]李怀伟.旱涝—低温交叉胁迫小麦对抗寒性的影响[D].哈尔滨:东北农业大学,2013.
[26]张敏.不同小麦品种耐盐差异的生理生化机制研究[D].泰安:山东农业大学,2007.
[27]宫德衬.小麦抗旱性生理指标研究[D].烟台:烟台大学,2014.
[28]陈成升,谢志霞,刘小京.旱盐互作对冬小麦幼苗生长及其抗逆生理特性的影响[J].应用生态学报,2009,20(4):811-816.
Effects of Salt and Drought Cross Stress on Osmotic Adjustment Ability of Wheat Seedling
HUAZhirui,LI Xiaoling
(College ofBiologyPharmacyand Food Engineering,ShangluoUniversity,Shangluo726000,China)
The paper took Shangmai 5226 seeds as experimental material,adopted the simulation method of intercross stress of drought and salt on wheat seedlings,under the intercross stress by measuring the wheat seed germination rate,germination potential, soluble sugar content,malondialdehyde(MDA)content,soluble protein content and other indicators,toexplore the change rule ofosmotic regulation substances of wheat with salt drought stress time,to provide theoretical basis for the cultivating of excellent wheat varieties of salt and drought resistance.The results showed that the germination rate and germination potential of wheat seeds decreased gradually with the increase of salt and drought stress.In 0.6%NaCl solution concentration,with 10%PEG stress had certain cross adaptation, germination rate was 83%,the germination potential was 96%,soluble sugar in 2 days reached maximum,and compared to the control increased by 0.32 mg/g,decreased the cell water potential,maintain the normal growth and metabolism of cells,malondialdehyde slowly increased and then decreased,eliminating the damage of reactive oxygen species of plants,improving the antioxidant capacity of plants, soluble protein in the second dayreached the maximum0.34 mg/g,and decomposition intovarious aminoacids,especiallyproline content increased,decreased water potential,reduced the salt and drought caused bydamage.
wheat;salt and drought cross stress;osmotic adjustment
S512.101
A文献标识码:1002-2481(2017)02-0166-06
10.3969/j.issn.1002-2481.2017.02.05
2016-10-21
陕西省科技厅农业攻关项目(2011K01-18)
华智锐(1980-),男,湖北黄石人,副教授,主要从事植物抗性生理与植物育种研究工作。
猜你喜欢
现代园艺(2020年19期)2020-10-02
初中生世界·八年级(2019年6期)2019-08-13
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
现代农村科技(2019年4期)2019-01-06
中成药(2018年8期)2018-08-29
小学生导刊(低年级)(2016年9期)2016-10-13
小学生导刊(低年级)(2016年6期)2016-07-02
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
