
管涔林区两种绣线菊幼苗对不同光环境的形态响应
2016-12-20 05:34任达王暾郭晋平张芸香
山西农业大学学报(自然科学版) 2016年11期
任达,王暾,郭晋平,张芸香
(1.山西农业大学 林学院,2.山西农业大学 城乡建设学院,山西 太谷 030801)
管涔林区两种绣线菊幼苗对不同光环境的形态响应
任达1,王暾1,郭晋平2,张芸香1
(1.山西农业大学 林学院,2.山西农业大学 城乡建设学院,山西 太谷 030801)
[目的]森林光环境是影响林下灌木植物正常生长和良好更新的重要因素之一。为了探求土庄绣线菊和三裂绣线菊一年生幼苗对不同森林光环境的适应特点和二者的区别,本研究选择林下和林外两种森林光环境。[方法]通过测定土庄绣线菊和三裂绣线菊一年生幼苗植株的高、地径、地径比、叶鲜重、比叶面积、叶面积比、生物量和生物量分配,从而得到两种绣线菊一年生幼苗在形态特征上的不同光适应性,为两种绣线菊的造林使用、更新和维护提供依据。[结果]结果表明:不同森林光环境下两种绣线菊的根、茎、叶生物量及总生物量表现出相同的规律。两种绣线菊在林内环境中均显著提高了地上生物量的比例。林外环境土庄绣线菊根生物量比显著高于林内植株,在叶生物量比指标上则相反。从株高、地径、高径比来看,林内遮荫环境明显降低了土庄绣线菊和三裂绣线菊的地径,同时显著提高了土庄绣线菊的高径比。从叶面积特点来看,两种绣线菊叶生物量林外和林下差异显著,而且比叶面积对光响应一致,林下环境使比叶面积明显增大。林下环境土庄绣线菊存活率略高于三裂绣线菊,林外环境三裂绣线菊存活率明显高于土庄绣线菊。说明二者并不是传统的喜光植物,三裂绣线菊对不同森林光环境有着很强的适应能力,土庄绣线菊具有较强的耐阴性。[结论]所以在以后的造林的计划中可以在林下多配置土庄绣线菊,在林外种植三裂绣线菊,以达到造林复层的目标,提高植被对水土保持的作用。
土庄绣线菊; 三裂绣线菊; 森林光环境; 形态; 生物量; 生物量分配
1 前言
植物适应光环境的能力是影响植物分布模式和物种丰度的重要因素之一[1]。光照条件的变化能够通过直接影响植物的生理生态特性进而影响植物的生存和生长[2]。林下植物可以通过生物量积累和分配等方面的响应而适应异质光环境[3~5]。林下灌木树种是林下植被的主要成分和森林群落的重要组成部分,对群落整体结构、功能和生态过程有重要影响[6],对构建复层结构森林群落,增强森林水源涵养、水土保持等生态服务功能方面的作用,对森林群落物种多样性、生产力和稳定性的作用,都在不断得到更多的认同。
土庄绣线菊(Spiraeapubescens)和三裂绣线菊(Spiraeatrilobata)是华北山地林区常见的灌木树种,既是分布区山地森林群落形成前灌丛群落的建群种,也是林下植被的重要灌木树种[7, 8]。一般认为两者都属于阳性树种,在阳坡灌丛植被中生长茂盛、发育良好,但相对而言土庄绣线菊比三裂绣线菊在林下和林窗中分布更多,显示两者对光环境的功能性适应特征有明显差异。
本文以土庄绣线菊和三裂绣线菊一年生实生苗为试验材料,通过设置林下和林外两种光环境,研究两者的生物生长量分配、形态响应。探讨两个树种对光环境的功能性适应特征,为揭示其光环境适应性及其与自然分布的关系,进而为基于森林群落物种多样性和稳定性及其与生产力关系的森林生态系统经营和植被生态恢复提供理论依据。
2 材料和方法
2.1 试验地区域环境概况
研究地设在山西省宁武县境内的芦芽山自然保护区内,属于吕梁山脉北段,地理位置在38°40′~38°50′N,111°20′~112°00′E,属暖温带半湿润区,大陆性山地季风气候,夏季凉爽多雨,冬季寒冷,年平均气温6 ℃~10 ℃,1月均温-8 ℃~-12 ℃,7月均温21 ℃~36 ℃,年降水量384~679 mm,年蒸发量1 800 mm,相对湿度50~55%,无霜期130~170 d。土壤主要是在残积和坡积母质上发育的山地森林土,呈垂直地带性分布,自下而上依次为山地褐土、山地淋溶褐土、棕色森林土和亚高山草甸土。芦芽山海拔1850 m以上地段,主要分布着以华北落叶松(Larixprincipis-rupprechtiiMayr)、麦氏云杉(PiceameyeriRehd.etWils.)和细叶云杉(Piceawilsoniimast)为建群种的寒温性针叶林,低中山分布着以油松、辽东栎、白桦、山杨为主的暖温带山地针叶林、落叶阔叶林及其替代性小叶林,山地落叶灌丛植被带各海拔带上都有分布,林下植被灌木层主要有土庄绣线菊(Spiraeapubescens)、三裂绣线菊(Spiraeatriblobata)、毛榛(CorylusmandshuricaMaxim)、胡枝子(LespedezabicolorTurcz.)、黄刺玫(RosaxanthinaLindl)和忍冬(LonicerajaponicaThunb.)等。
2.2 试验材料
2009年9月,种子成熟季节在当地采种,经净种水选后用0.5%高猛酸钾溶液进行表面消毒灭菌,选择4.5 cm×4.5 cm×5.5 cm规格的苗盘,置于培养箱内育苗,温度25 ℃,湿度45%,白天12 h晚上8 h保存。2010年3月放入温室缓苗,到4月中旬,选择200株生长健康的幼苗,平均苗高6±1 cm,平均地径0.4±0.15 mm,将幼苗连同苗盘一起运至试验地点,缓苗2周。
2.3 试验设计与试验地设置
试验采用裂区设计,设置林外和林下2种光环境,选择晴天中午,使用LI-190SA辐射传感器(L-COR lin.,Nebraska,U.S.A)在两个样地同时测定光照强度,并相应计算林下平均光照强度和透光率,测定和计算各样地日平均光通量,林外和林内的日平均光通量分别为59 mol·d-1和10 mol·d-1。每种光环境设置2个区组,每个区组面积4 m×4 m,每个区组定植50株,区组间距≥10 m。
采用泥炭土与蛭石体积比1∶1混合物为培养基质,加配1 kg/m3的NPK复合肥作基肥,实验期间不再添加任何肥料。采用直径8 cm、高10 cm的带孔营养袋。初次栽植时浇透水1次,以后除每周清除杂草1次外,不采取其他特殊人工管理措施。
3 指标测定与数据处理
3.1 指标测定
指标测定在2010年9月8日进行。
(1)苗木成活率测定:统计两种灌木的最终存活株数,计算成活株数占中栽植株数之比,按百分率计成活率;
(2)形态指标测定:用电子游标卡尺量测地径和苗高,计算高径比(H/D);
(3)生物量指标测定:分别在林内和林外试验地内取全部幼苗,剪取植株根、茎和叶,洗净晾干,分别置于80 ℃的在鼓风干燥箱中烘72 h左右至恒重,用精度0.001 g的天平分别测定其干质量,记录根Mr、茎Ms、叶Ml和总生物量Mt。
(1)
(2)
(3)
(4)
其中RMR是根重比,SMR是茎重比,LMR是叶重比,A/B是冠根比。
(4)比叶面积测定:以相邻五株为一组,测定投影叶面积。利用winseedle分析系统(Regent Instruments Inc., Sainte-Foy, QC, Canada)对每一植株的叶片进行数字化扫描和分析,得到投影叶面积。并将叶片放置在80℃烘箱中烘烤,至恒重。用精度0.001 g的天平称量叶干重,精确到0.01 g。计算比叶面积(SLA):为单株总叶面积与叶生物量之比。
3.2 数据处理
对存活率采用(Chi-Square value)检验差异显著性,对其它指标采用Data Desk 6.1.1进行光照和物种的双因素方差分析,对差异显著(p<0.05)的指标用LSD检验进行多重比较。
4 结果和分析
4.1 两种光环境下两种绣线菊幼苗的表型差异分析
对不同森林光环境下土庄绣线菊和三裂绣线菊的形态指标进行双因素方差分析,结果见表1。
表1 不同森林光环境下两种灌木幼苗形态和生物量分配指标方差分析结果
Table 1 P-values for themorphological and biomass allocation parameters of two shrub seedings growing under two light regimes
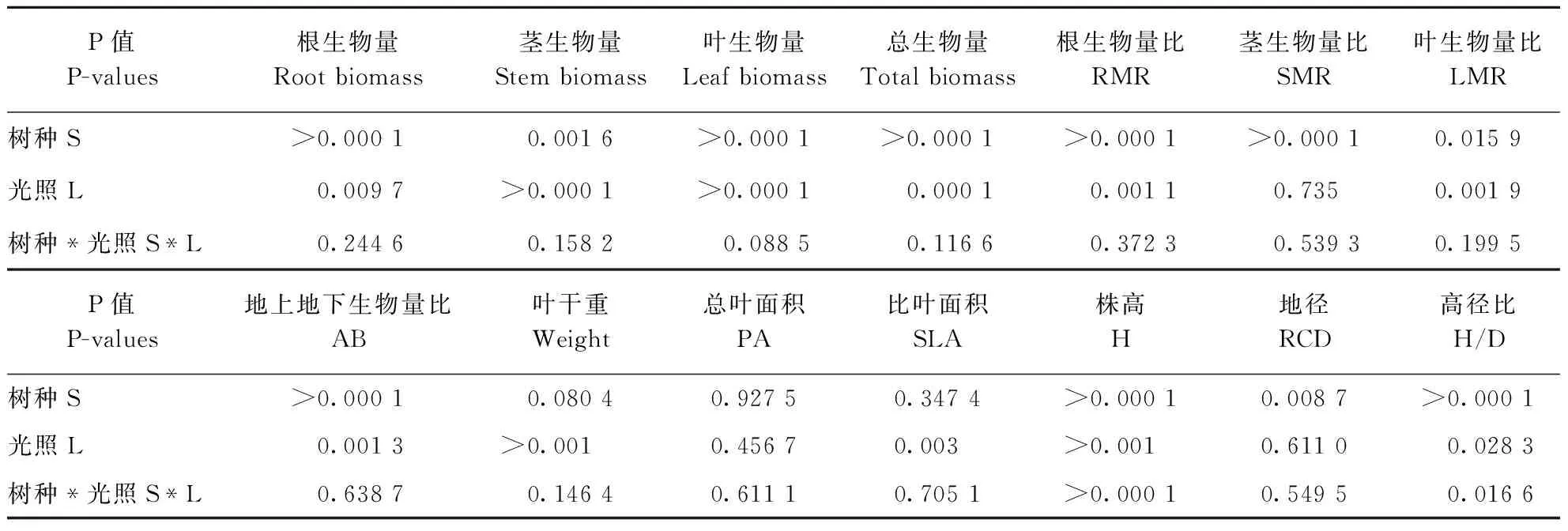
P值P-values根生物量Rootbiomass茎生物量Stembiomass叶生物量Leafbiomass总生物量Totalbiomass根生物量比RMR茎生物量比SMR叶生物量比LMR树种S>0.00010.0016>0.0001>0.0001>0.0001>0.00010.0159光照L0.0097>0.0001>0.00010.00010.00110.7350.0019树种*光照S*L0.24460.15820.08850.11660.37230.53930.1995P值P-values地上地下生物量比AB叶干重Weight总叶面积PA比叶面积SLA株高H地径RCD高径比H/D树种S>0.00010.08040.92750.3474>0.00010.0087>0.0001光照L0.0013>0.0010.45670.003>0.0010.61100.0283树种*光照S*L0.63870.14640.61110.7051>0.00010.54950.0166
树种之间各形态指标除叶干重、总叶面积、比叶面积外存在显著或极显著差异;除茎生物量比、总叶面积、地径外,光照对其他形态指标存在显著的影响;除株高和高径比外,树种和光照交互作用对其他形态指标不存在显著的影响。
4.2 生物生长量响应
由图1可见,不同光环境下两种绣线菊一年生苗的根、茎、叶生物量及总生物量变现出相同的规律,两种绣线菊一年生幼苗各器官生物量及总生物量林外均高于林下,但土庄绣线菊一年生幼苗各生物量指标在林内外差异不显著,说明遮荫环境并不会明显抑制土庄绣线菊幼苗的正常生长和发育。三裂绣线菊幼苗在林外的各生物量指标显著高于林下,林外环境中的幼苗在根生物量,茎生物量,叶生物量及总生物量指标上分别比林下的植株高163%、201%、178%及179%。
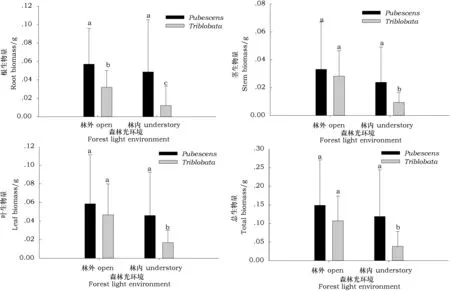
图1 不同森林光环境下两种绣线菊幼苗生物量指标比较Fig.1 The biomass of two natural shrub seedings growing under two light regimes 注:小写字母表示两种树种在不同光环境下的差异显著性(p<0.05),下同。(Note: Different low-case letters indicate the result of multiple comparisons between different light regimes in different shrub species(p<0.05), The same below.)
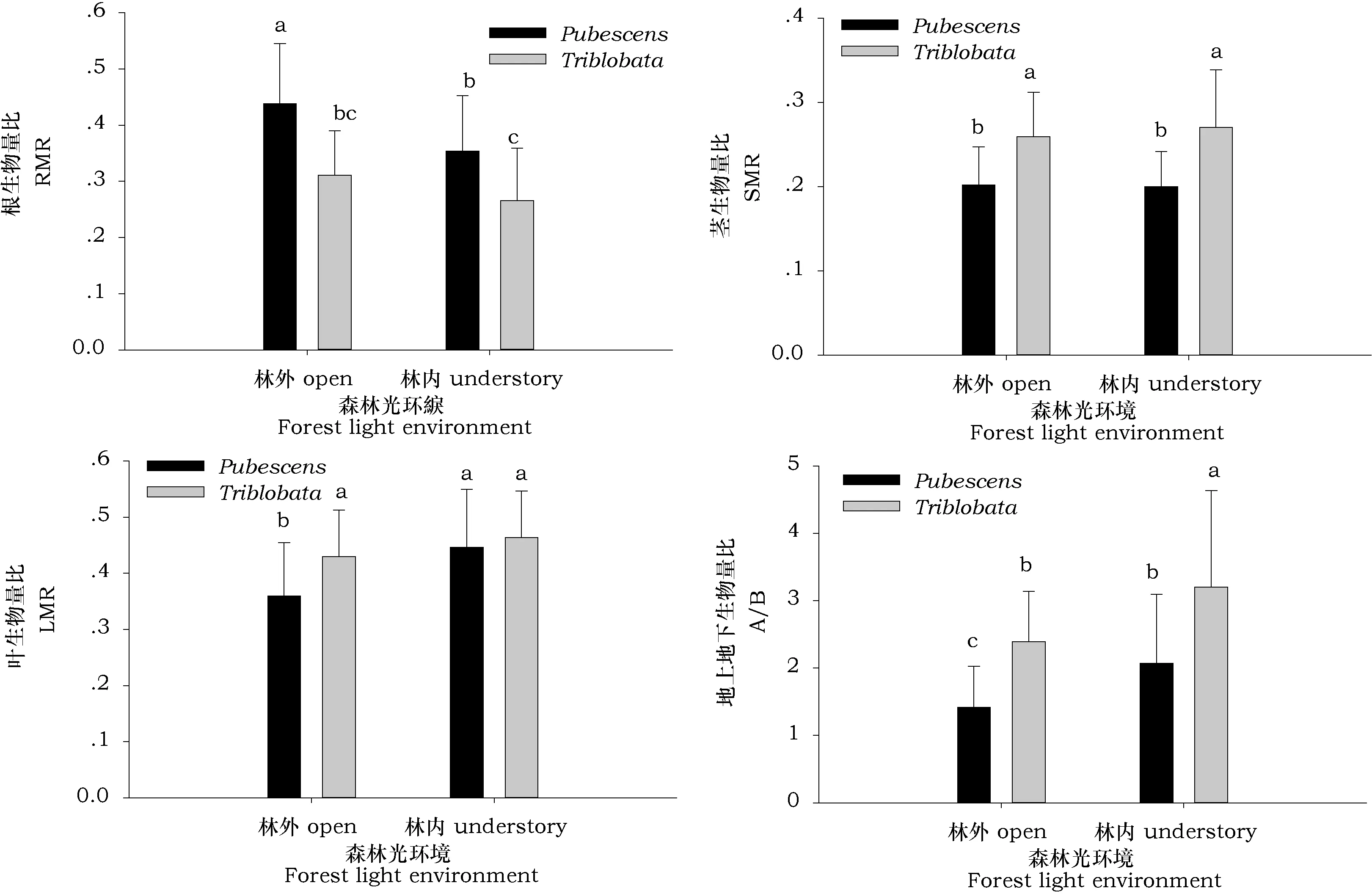
图2 不同森林光环境下两种绣线菊幼苗生物量分配指标比较Fig.2 The biomass allocation of two natural shrub seedings growing under two light regimes
4.3 生物量分配响应
从生物量分配来看(见图2),两种绣线菊一年生幼苗对不同光环境的响应趋势大致相同,林内环境中地上生物量均显著提高,土庄绣线菊和三裂绣线菊林下比林外分别提高67.6%和33.8%。不同森林光环境对两种绣线菊幼苗的茎生物量比没有明显的影响。遮荫提高了土庄绣线菊的叶生物量比,林内比林外提高24%;幼苗根生物量在林内比林外下降19.2%。说明土庄绣线菊和三裂绣线菊一年生幼苗积极调整自身物质的分配来适应低光环境。
4.4 植株形态响应
从株高、地径、高径比三个植株形态指标来看(见图3),两种绣线菊一年生幼苗表现出类似的光环境响应。两种绣线菊幼苗的地径在林内遮荫环境中比林外分别降低20.6%和15.3%,且高径比更大。三裂绣线菊株高生长在低光环境中受到严重抑制,与林外相比下降33.9%。土庄绣线菊一年生幼苗在林下有着更高的高径比,与林外的幼苗相比提高了34.2%。说明为了适应低光环境并获取更多的光能,土庄绣线菊幼苗提高了光合面积,这也与土庄绣线菊幼苗地上地下生物量比的结果相一致。

图3 不同森林光环境下两种绣线菊幼苗株高、地径和高径比指标比较Fig.3 Heights, root collar diameters (RCDs) and H/D of two natural shrub seedings growing under two light regimes
4.5 叶形态响应
两种绣线菊一年生幼苗的总叶面积和叶生物量变化趋势相近(见图4),遮荫环境明显抑制两种绣线菊叶的生长,土庄绣线菊和三裂绣线菊总叶面积分别降低21.9%和21.1%,叶生物量分别降低63.9%和54.2%,而比叶面积分别增大93.4%。和68.7%。说明两种绣线菊幼苗在低光环境下虽叶生物量合成减少,但还是投入更高比例的叶生物量以满足对光能的获取。
4.6 幼苗移植存活率的响应
两种绣线菊幼苗林外林下存活率不同(见表2),林下环境土庄绣线菊存活率略高于三裂绣线菊,林外环境三裂绣线菊存活率明显高于土庄绣线菊。土庄绣线菊林下存活率较高而林外最低,说明其幼苗适于低光环境下生长。三裂绣线菊幼苗对林内外环境的适应力相当。
5 结论与讨论
光照是林下更新中最重要的环境因素,是决定灌木生长、发育和完成正常更新过程的关键,因此植物必须增强适应能力才能适应异质的光环境,光照过强或过于阴蔽都会限制植物的生长和生存[9],植物对光异质性的响应既有比较直接的生理适应,也有长期的形态适应[10]。其中,植物对于光环境的形态学响应主要体现在地径(D)、高度(H)、比叶面积(SLA)、生物量积累和分配等指标上。
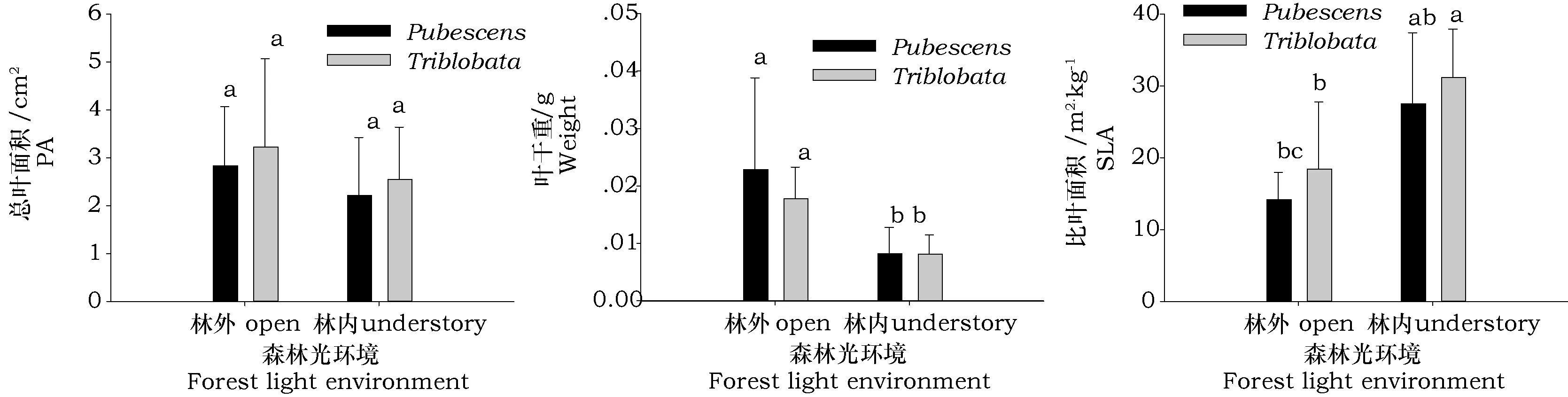
图4 不同森林光环境下两种绣线菊幼苗叶形态指标比较Fig.4 Leaf morphology of two natural shrub seedings growing under two light regimes
Table 2 The seedlings survival rates of two shrub species

树种Species林外Open林内Understory平均值Average卡方值Chi-Squarevalue土庄绣线菊Spiraeapubescens69.44%100.00%84.72%19.97三裂绣线菊Spiraeatrilobata94.38%97.14%95.76%0.259
地径和高度作为的基本的形态学指标,能反映植物对生长环境的适应,而阳生和阴生植物形态特征的光响应也并不一致。李晓征等[11]发现耐阴的红花木莲、醉香含笑和金叶含笑在40%光强下株高生长显著高于100%光环境,华北落叶松除当年播种苗需遮阴外,以后各个生育期需要在全日照条件下生长发育,未遮阴三年生华北落叶松苗木,苗高比遮阴的大40~50%,地径大45~50[12]。在本文的研究中发现三裂绣线菊和土庄绣线菊幼苗株高在林内外没有明显差异,而在地径指标上林外的明显高于林内的,林下高径比显著增加,其通过形态改变来获得更多的光照。说明土庄绣线菊和三裂绣线菊对低光环境有一定的适应性。
比叶面积是体现植物光捕获能力的重要指标,一般与光环境高度相关[13, 14]。比叶面积较大的物种,平均叶寿命低,但光捕获面积、单位重量叶氮含量较高,因此净光合速率较高[15]。一般来说,在冠层中最阴蔽的地方形成的叶子才具有最高的比叶面积[9]。在遮阴环境下,具有较高的比叶面积是有效地捕获有限光资源的一种适应性响应[13]。本研究中处于林下低光环境的土庄绣线菊和三裂绣线菊幼苗表现出了较高的比叶面积,这与Han等[13]得出的结论一致,表明两种绣线菊可以通过增大叶面积捕获更多的光能来适应低光的环境。
植物生物量及其分配能够反映出植物生长过程中资源或能量分配的一般特性[16]。生物量分配随光照强度变化而变化可能是植物能够在不同光环境下生存的重要原因,植物生物量及其分配能够反映出植物生长过程中资源或能量分配的一般特性[17]。陈圣宾等[18]和King[19]认为植物幼苗会在较弱光照环境中将更多生物量投入到叶中以提高光捕获能力。也有人认为,植物对弱光环境的适应主要是提高弱光环境下维持生存的能力,而不是提高其碳收获能力[9]。在本文的研究中发现,通过测定,林外环境土庄绣线菊根生物量比显著增加,叶生物量比,地上地下生物量比显著降低,在林外增加根系有利于对水分的吸收、林下提高叶生物量分配有利于对光资源的捕获,同时,与三裂绣线菊相比,土庄绣线菊在林外环境具有更高的根生物量比、更低的叶生物量比和地上地下生物量比,说明土庄绣线菊通过最大程度的改变生物量分配以适应全光照环境。三裂绣线菊的光响应趋势与土庄绣线菊相同,只有地上地下生物量比在林外与林下差异显著。
存活率是检验植物对光环境适应与否最直接明显的指标。通过存活率比较,可以明显看出,土庄绣线菊对低光环境适应能力很强,在强光环境下适应能力差,三裂绣线菊在林内外光适应性都很强。这也与土庄绣线菊在林下常见,而三裂绣线菊常分布于林外旷地及阳坡相一致。总体来说,三裂绣线菊的光适应性高于土庄绣线菊。因此在山地生态修复时可以根据两种绣线菊的特性采取不同的保护措施,在林外增加三裂绣线菊的植被覆盖,在林内增加土庄绣线菊的比例来形成最佳的群落结构。
[1]冯玉龙,曹坤芳,冯志立,等.四种热带雨林树种幼苗比叶重,光合特性和暗呼吸对生长光环境的适应[J].生态学报,2002,22(6):901-910.
[2]张秀茹.不同水分和光照处理对紫穗槐幼苗生理生态特征的影响[D].山东大学,2010.
[3]张卫强,王明怀,曾令海,等.东江中上游3种造林树种的光合-光响应特征[J].中国水土保持科学,2010,8(6):115-120.
[4]王荣,郭志华.不同光环境下枫香幼苗的叶片解剖结构[J].生态学杂志,2007,26(11):1719-1724.
[5]孙一荣,朱教君,于立忠,等.不同光强下核桃楸、水曲柳和黄菠萝的光合生理特征[J].林业科学,2009,45(9):29-35.
[6]吕丽丽.关帝山云杉次生林下灌木树种的多样性及其分布格局[D].山西农业大学,2015.
[7]桂炳中,陈东青,高惠茹.华北地区三裂绣线菊栽培养护[J].中国花卉园艺,2015(14):44-45.
[8]冯强,刘宁,王暾.不同自然光环境下土庄绣线菊的生理生态响应[J].中南林业科技大学学报,2010, 30(9): 27-33.
[9]Valladares F, Wright S J, Lasso E, et al. Plastic Phenotypic Response to Light of 16 Congeneric Shrubs from a Panamanian Rainforest[J]. Ecology,2008, 81(7): 1925-1936.
[10]Schlichting C D. The Evolution of Phenotypic Plasticity in Plants[J]. Annual Review of Ecology & Systematics. 2003, 17(1): 667-693.
[11]李晓征,彭峰,徐迎春,等. 不同光强下6种常绿阔叶树幼苗的生理特性[J]. 南方农业学报. 2005, 36(4): 312-315.
[12]牛云,张宏斌,潘爱华,等. 张掖地区华北落叶松苗期生长规律的研究[J]. 河西学院学报. 2003(2): 47-50.
[13]Han Y. H. Chen K K G J. Effects of light on growth, crown architecture, and specific leaf area for naturally established Pinus contorta var. latifolia and Pseudotsuga menziesii var. glauca saplings[J]. Canadian Journal of Forest Research. 1996, 26(7): 1149-1157.
[14]Reich P B, Walters M B, Tjoelker M G, et al. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate[J]. Functional Ecology. 1998, 12(3): 395-405.
[15]Ellsworth D S, Reich P B. Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest[J]. Oecologia. 1993, 96(2): 169-178.
[16]杨昊天,李新荣,刘立超,等. 荒漠草地4种灌木生物量分配特征[J]. 中国沙漠. 2013, 33(5): 1340-1348.
[17]Montgomery R. Relative importance of photosynthetic physiology and biomass allocation for tree seedling growth across a broad light gradient[J]. Tree Physiology. 2004, 24(2): 155-167.
[18]陈圣宾,宋爱琴,李振基. 森林幼苗更新对光环境异质性的响应研究进展[J]. 应用生态学报. 2005, 16(2): 365-370.
[19]King D A. Allocation of above-ground growth is related to light in temperate deciduous saplings[J]. Functional Ecology,2003, 17(4): 482-488.
(编辑:梁文俊)
Morphological characteristics of theSpiraeapubescensandSpiraeatrilobataseedlings growing under two forest light environments in GuanCen Mountains
Ren Da1, Wang Jun1, Guo Jinping2, Zhang Yunxiang1
(1.CollegeofForestryShanxiAgriculturalUniversity, 2.CollegeofUrbanandRuralConstruction,ShanxiAgriculturalUniversity,Taigu030801,China)
[Objective] Forest light environment is an important factor affecting forest tree growth and complete the normal renewal process. In order to explore the illumination change influence onSpiraeapubescensandSpiraeatrilobataseedlings,we selected two kinds of forest light environments,open and understory.[Methods]Determination ofSpiraeapubescensandSpiraeatrilobataseedlings height, diameter ratio, specific leaf area, leaf area ratio, biomass and biomass allocation,light adaptability ofSpiraeapubescensandSpiraeatrilobataseedlings in the physiological and ecological characteristics, for spiraea’s forest renewal, maintenance and provide the basis for recovery. [Results] The results show that: The root, stem, leaf biomass and total biomass of the two kinds of spiraeain in different forest light environment showed the same rule. Two species of spiraea in forest environment significantly increased the proportion of above ground biomass. When pubescens seeds in open area, their root biomass is significantly increase in leaf biomass ratio index instead. From the plant height, ground diameter, the ratio of height to diameter, canopy shade environment obviously and theSiraeapubescensandSpiraeatrilobatasize decrease, and significantly improve theSpiraeapubescensratio of height to diameter. From the characteristics of two kinds of leaf area, leaf biomass of spiraea forest was significant. The response of the leaf area was the same to two kinds of spiraea in understory,and their leaf area were significantly increased. In the shading environment, the survival rate of theSpiraeapubescensis slightly higher than the survival rate ofSpiraeatrilobata. The survival rate ofSpiraeatrilobatawas significantly higher thanSpiraeapubescensin shading environment. The two kinds spiraea are not traditional hi light plants, spiraea trilobata in different forest light environment has a strong ability to adapt,Spiraeapubescenswith strong shade tolerance. [Conclusion] In the future afforestation plan, pubescens seedlings could plant under high trees, in the open environment, we could plant moreSpiraeatrilobataseedlings, so we can achieve the goal of afforestation, and enhancing the effect of vegetation on soil and water conservation.
Spiraeapubescens,Spiraeatriblobata, Forest light environmental, Morphological response, Biomass, Biomass allocation
2016-06-24
2016-07-12
任达(1991-),男(汉),山西榆社人,硕士研究生,研究方向:森林生态
*通讯作者:郭晋平,教授,博士生导师,Tel:13935406926;E-mail: jinpguo@126.com
山西省国际合作项目 (2015081001);山西省攻关项目(20140311014 -3); 高等学校博士基金(20121403110001)
S793
A
1671-8151(2016)11-0815-06
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
今日农业(2020年19期)2020-12-14
新疆农业科学(2020年1期)2020-02-14
现代园艺(2018年2期)2018-03-15
中国林业产业(2016年5期)2016-04-03
中国林业产业(2016年5期)2016-04-03
中国麻业科学(2015年5期)2015-12-28
中国科技信息(2015年2期)2015-11-16
武夷学院学报(2014年5期)2014-07-19
植物营养与肥料学报(2014年1期)2014-03-11
