
甘肃马先蒿生物量分配及其寄主物候对覆雪的响应
2016-12-09 06:17胡玉昆
草业科学 2016年10期
陆 均,胡玉昆
(1.中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011; 2.中国科学院大学,北京100049)
甘肃马先蒿生物量分配及其寄主物候对覆雪的响应
陆 均1,2,胡玉昆1
(1.中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011; 2.中国科学院大学,北京100049)
在研究区冬季平均降雪深±21.54%的覆雪处理水平下,对新疆巴音布鲁克高寒草原甘肃马先蒿(Pediculariskansuensis)及其寄主的物候以及甘肃马先蒿的生物量分配进行研究,并揭示其和寄主的竞争及繁殖策略。结果表明,1)覆雪处理对甘肃马先蒿及其寄主生长季变化的影响无一致规律;2)覆雪处理使得甘肃马先蒿及其大部分寄主的花期物候提前,秋季物候延后,甘肃马先蒿物候开始时间从结实期开始逐渐提前,到种子散播期几乎完全领先寄主;3)覆雪处理后甘肃马先蒿的繁殖分配和根冠比显著降低(P<0.05);4)增雪处理后甘肃马先蒿的株高、冠幅、花柱高度以及侧根尺寸与自然降雪间均有显著差异(P<0.05),而减雪处理后除侧根外各部分差异不大。总体上,甘肃马先蒿与寄主的物候在覆雪处理后存在一定程度分化;其生物量分配及个体形态对增雪处理的响应更显著,覆雪处理后甘肃马先蒿降低繁殖投资从而增加营养投资。
高寒草原;甘肃马先蒿;物候;生物量分配;覆雪
据联合国政府间气候变化专门委员会(IPCC)第五次评估报告,北半球1983-2012年可能是过去1 400年中最暖的30年[1]。气候变暖背景下新疆地区降雪年际多变,南、北疆及天山山区积雪深度均呈小幅增长,其中天山山区增幅最大[2-3]。雪是热量和水分的缓冲介质,适当深度的雪层覆盖有利于植物生长和繁殖[4-5]。雪生植物长期生长在严酷的生境中,在物候特征、繁殖对策、生长节律和生理生态等方面逐渐形成了一系列独特的适应机制[4,6]。因此,从个体生态学的角度,系统深入地研究某些关键植物的物候、繁殖对策和生长节律,有利于阐明这类植物对极端环境的适应机制[7]。
动植物物候期是气候变化的“诊断指纹”[8]。植物物候是植物应对环境变化时表现出的一系列生活史对策的体现,关系到植物体本身的生存繁衍和群落生物多样性的维持[9],植物物候对气候变化相对较小的响应也可以产生显著的竞争结果[10]。雪被覆盖使得植物物候产生延迟现象。融雪较晚的洼地植物在春季推迟返青,秋季物候延迟[7]。许多植物都具有在雪融化后迅速出叶和开花,甚至在雪下开花的特征,反映出雪场生境植物生长期短暂的一种适应机制[11]。
植物生活史对策对气候变暖的响应和植物群落结构的变化机制是近二十多年来生态学研究的热点[12-13]。植物生物量分配是指植物将一年所同化的资源用于该器官构件的比例,实际上是指总资源分配给该器官的比例[14]。近年来根部半寄生植物甘肃马先蒿(Pediculariskansuensis)在多地出现蔓延趋势[15-17],而相关研究大多集中在形态特征分析[18-19]、化学防除[20]、养分限制[21]、修剪寄主的影响[22]等方面,也有生物量分配的报道[16],但甘肃马先蒿物候研究或者结合气候变化特别是冬季降雪变化的研究还处于空白。在新疆地区年际降雪多变、积雪厚度增加的趋势下,通过覆雪处理模拟极端降雪年份,对甘肃马先蒿在开花期的资源分配对策及其寄主的物候进行研究,以期为深入了解两者在不同生长阶段的竞争关系特别是寄生关系,揭示甘肃马先蒿的种群繁殖和扩张策略提供新的思路。
1 材料与方法
1.1 研究区域概况
研究区位于新疆维吾尔自治区和静县巴音布鲁克草原。该草原地处天山中段南麓,海拔约为2 400~3 800 m,地势相对平坦,由西北向东南倾斜,总面积23 853 km2[15]。年均气温-4.8 ℃,1月最低气温至-48 ℃,7月最高气温为30.5 ℃,年均降水量265.7 mm,年均蒸发量1 022.9~1 247.5 mm,年日均温≥10 ℃的积温227.2 ℃·d,年日照时数2 466~2 616 h,全年积雪日150~180 d,无绝对无霜期,属典型的高寒气候,主要土壤类型为亚高山草原土,成土母质为黄土,表土干燥紧实[23]。主要草地类型有高寒草原、高山沼泽、高寒草甸。当地高等植物有50科160属262种,主要优势物种有羊茅(Festucaovina)、紫花针茅(Stipapurpurea)、冰草(Agropyroncristatum)、嵩草(Kobresiabellardii)和草地早熟禾(Poapraensis)等[24]。
研究区内与甘肃马先蒿伴生的常见植物有9科20种,均可被甘肃马先蒿寄生[25]。近年来甘肃马先蒿在新疆蔓延面积已达到2.33万hm2,并以每年0.33万hm2的速度扩展[15];马先蒿种群数量迅速膨胀,被认为是入侵型毒杂草,加速草地退化[16],并对畜牧业生产造成了严重危害。
1.2.1 样地设置 样地位于中国科学院巴音布鲁克草原生态试验站西北方向2 km处,地理坐标为83°42′07″ E,42°53′28″ N,海拔2 500 m。样地于2014年4月实行铁丝网围封避免放牧干扰,规格为边长100 m×100 m,避开铁丝网边缘自东向西设置样方,样方规格为3 m×4 m,样方之间缓冲区宽度为2 m。每个样方内对角设置两个破坏性取样小区,大小为1 m×1 m。
1.2.2 覆雪处理 设置增雪、减雪和对照(自然降雪,CK)3个处理,每个处理3个重复。在减雪处理样方架设用以支撑挡雪的透光塑料挡板的铁架,铁架高0.5 m,挡板长宽比样方长宽各大0.5 m,四周用塑料网格包围防止大风将雪刮进该样方。覆雪处理在2014年冬季(以12月至次年2月定为研究区冬季[3])进行,选取其中若干次降雪将挡板上的积雪转移到增雪处理样方,并根据研究区冬季平均降雪深[3],设定覆雪处理量约占冬季平均降雪深25%,计算出覆雪总深度约为48.0 cm。每次覆雪试验完成后及时拆除挡板避免其它降雪事件以及挡板造成的影响。
1.2.3 物候观测 选取甘肃马先蒿及其较有代表性的寄主[25]共6个物种(表1)作为物候观测对象,避开破坏性取样小区和样方边缘,每个物种在每个处理随机标记30株[26](比如羊茅在减雪处理的3个样方内一共标记30株),物候调查在2015年4月-9月进行,以儒略历(1月1日为第1天)作为时间量度[27],调查时间从每种观测物种的萌发延续到种子散播,每次调查时间间隔一般4~6 d,花期3~5 d[28]。调查时用钢卷尺测量每株高度,观察并记录其所处的物候期。物候期记录项目区分禾本科和一般草本:1)一般草本:返青期、营养期、始花期、开花盛期、花末期、结实期和果实成熟期;2)禾本科花药较小,难以区分花期状态,故分:返青期、展叶期、分蘖期、抽穗期、结实期和果实成熟期。其中,始花期为有10%植株上初次有个别花瓣完全展开时,开花盛期为有50%植株处于开花期,花末期为有80%植株处于开花期[29]。
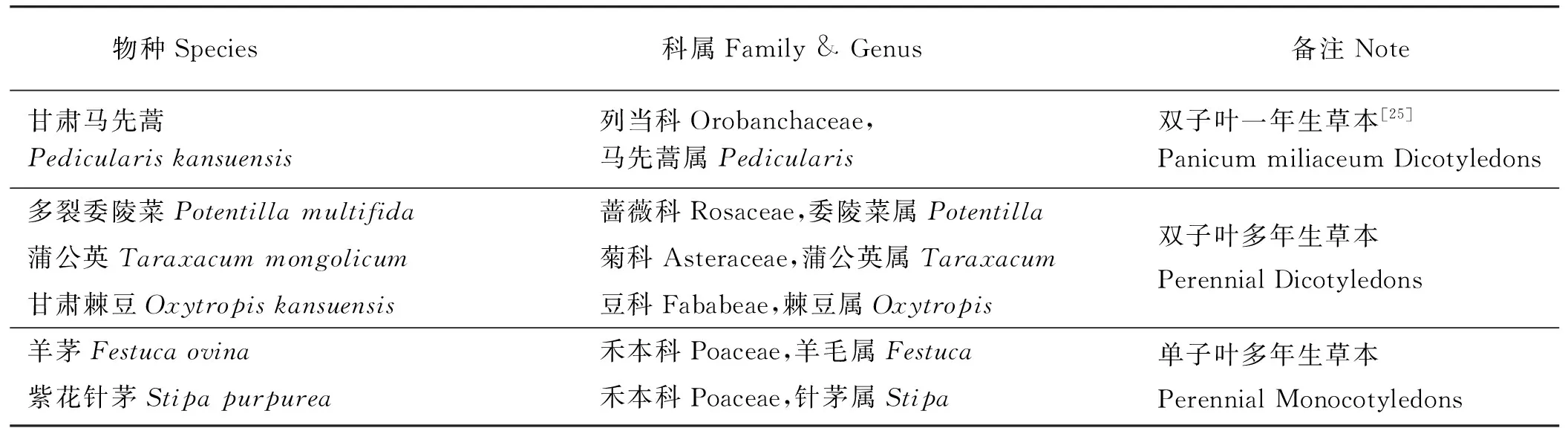
表1 研究对象概况Table 1 Overview of the study objects
1.2.4 取样和数据获取 于甘肃马先蒿的开花盛期,在每个处理的破坏性取样小区随机选取甘肃马先蒿15株,测量其植株高度、花柱高度、冠幅和分枝数[16];并以该株甘肃马先蒿的冠幅投影为范围,记录冠幅下出现的物种及其数量;用十字镐小心挖取整株所选取的甘肃马先蒿,并测量其侧根的长宽以及根深,标记后在室内将每株甘肃马先蒿的根、茎叶和花三部分构件分开袋装,以便烘干后称取各部分的生物量,其中花构件包括其支持部分[30];各部分构件的生物量分配计算:根分配=根部分生物量/个体生物量,茎叶分配=茎叶生物量/个体生物量,花分配=花生物量/个体生物量[30]。在9月底调查样方内物种的组成、盖度及其垂直高度;用刈割法收取地上生物量,并根据禾本科、豆科和杂类草分开挑选,标记后置于烘箱烘至恒重再用电子天平称量干重;土壤温度用地温计组测量,深度范围为5、10、15、20和25 cm;气温和湿度用美国Onset公司的HOBO测量,每30 min自动记录一次数据;降雪量由中科院巴音布鲁克草原生态试验站的气象站获得。
1.2.5 数据处理 采用Excel 2007和SPSS 19.0软件对数据进行统计分析;用LSD method检验比较各处理间的差异,显著水平取P<0.01为差异极显著,P<0.05为差异显著;作图用Excel 2007。
2 结果与分析
2.1 覆雪处理对甘肃马先蒿及其寄主物候的影响
2014年冬季期间共进行5次覆雪处理,累计覆雪深度42.0 cm,占该区冬季多年平均降雪深的21.54%。覆雪处理对甘肃马先蒿及其寄主的生长季和不同物候期产生不同影响(图1)。从生长季看,覆雪处理(增雪和减雪)使得甘肃马先蒿生长季缩短,且增雪处理比减雪处理更明显,甘肃棘豆对覆雪处理的响应表现出与甘肃马先蒿相同的趋势,但其生长季平均持续只有130.3 d;覆雪处理(增雪和减雪)使得植物生长季缩短的还有紫花针茅和羊茅,但对这两种植物而言减雪处理的作用比增雪处理更明显;另外,蒲公英各物候阶段比其它物种提前,覆雪处理(增雪和减雪)使其生长季延长,且增雪处理更明显,表现为增雪>减雪>对照;
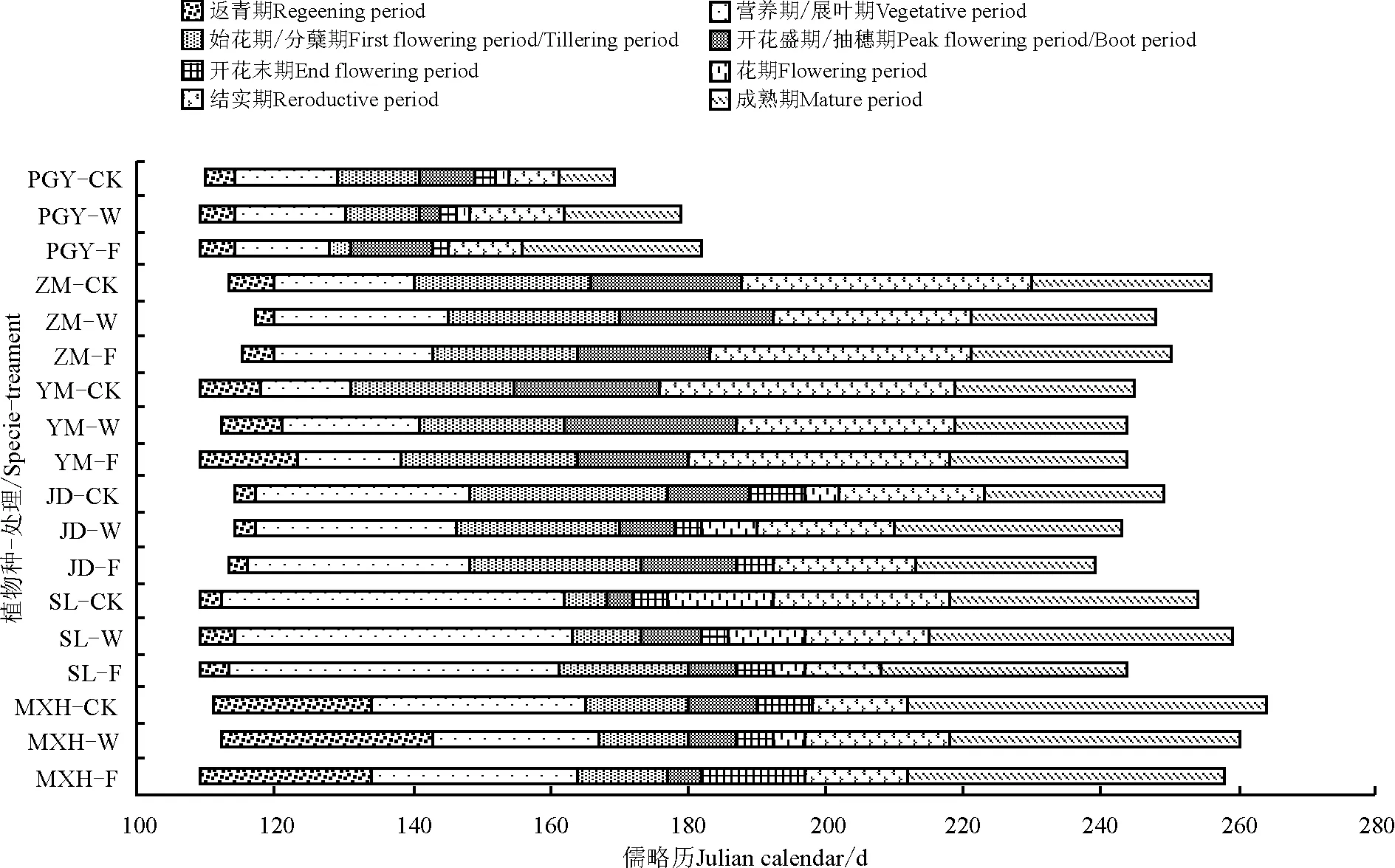
图1 覆雪处理下甘肃马先蒿及其寄主的物候图谱Fig.1 The phenological map of Pedicularis kansuensis and its hosts under snow cover change
注:MXH,甘肃马先蒿;SL,多裂委陵菜;YM,羊茅;ZM,紫花针茅;PGY,蒲公英;JD,甘肃棘豆;F,增雪处理;W,减雪处理;CK,对照。始花期、开花盛期和开花末期对应为该段末端的时间。
Note: MXH,Pediculariskansuensis; SL,Potentillamultifida; YM,Festucaovina; ZM,Stipapurpurea; PGY,Taraxacummongolicum; JD,Oxytropiskansuensis; F, snow cover addition; W, snow cover reduced; CK, control group. The beginning of flowering period, The full of flowering period and the end of flowering period is corresponding to the time of that stage.
减雪处理使得多裂委陵菜的生长季延长,增雪处理使得其生长季缩短。故覆雪处理(增雪和减雪)使得甘肃马先蒿、甘肃棘豆、紫花针茅和羊茅的生长季缩短,蒲公英的生长季延长,多裂委陵菜表现不一致,总体上无一致规律。
【查阅资料】制作固体酒精时,用由硬脂酸和氢氧化钠反应生成的硬脂酸钠作为凝固剂,再加入酒精,酒精填充在硬脂酸钠骨架间隙中,即可得到固体酒精。固体酒精在较高的温度下燃烧时,硬脂酸钠可完全转化为硬脂酸和氢氧化钠,硬脂酸和酒精均可完全燃烧生成二氧化碳和水。
从不同生长阶段的物候期看,增雪处理使得甘肃马先蒿、甘肃棘豆和蒲公英的春季物候期平均提前约1.3 d,紫花针茅春季物候期推迟2.0 d;减雪处理使得甘肃马先蒿、羊茅和紫花针茅春季物候期平均推迟3.0 d,蒲公英春季物候期提前1.0 d;多裂委陵菜的春季物候期开始时间无变化。从花期物候(一般草本包括始花期、开花盛期和开花末期,禾本科为抽穗期和花末期)看,增雪处理使得甘肃马先蒿、多裂委陵菜、紫花针茅和蒲公英的花期物候期提前,羊茅花期物候期推迟;减雪处理使得甘肃马先蒿、多裂委陵菜、紫花针茅、羊茅和蒲公英的花期物候期提前,甘肃棘豆花期物候延后。从秋季物候期(结实期和成熟期)看,覆雪处理使得一些物种的秋季物候期提前幅度较大,其中增雪处理使得甘肃马先蒿、多裂委陵菜和羊茅的秋季物候期平均推迟4.0 d,其它物种期平均提前12.0 d;减雪处理使得蒲公英和甘肃棘豆秋季物候期平均提前9.0 d,其它物种平均推迟5.8 d。
2.2 覆雪处理下甘肃马先蒿的生物量分配
覆雪处理下甘肃马先蒿的生物量分配存在差异(图2),但不同处理下甘肃马先蒿各部分的生物量分配由大到小顺序一致,表现为茎叶>花>根。与对照相比,覆雪处理(增雪和减雪)后甘肃马先蒿的繁殖分配(花分配)降低;根分配在增雪处理后显著降低(P<0.05),而茎叶分配有所提高,但不显著(P>0.05);总体上营养分配提高。覆雪处理(增雪和减雪)后甘肃马先蒿的根冠比降低,且增雪处理较对照效果显著)。另外,减雪处理与增雪处理虽然为两种极端处理,但两者之间甘肃马先蒿各构件的生物量分配差异并不显著。
2.3 覆雪处理下甘肃马先蒿个体形态特征的响应
覆雪处理后甘肃马先蒿植株各部分生长器官表现出差异性响应(表2)。覆雪处理(增雪和减雪)后,甘肃马先蒿的分枝数和根深在增雪、减雪和对照之间无显著差异(P>0.05);而增雪处理后,甘肃马先蒿的株高、冠幅、花柱高度以及侧根尺寸与对照相比均发生显著差异(P<0.05),表现为增雪的株高和侧根尺寸更大,而冠幅和花柱高度降低;减雪处理后除了侧根尺寸与对照相比有显著差异外,其它部分均无显著差异;增雪与减雪相比,增雪的株高、冠幅和花柱高度增大,同时差异显著,其它如分枝数、根深和侧根尺寸虽存在一定差异,但并不显著(P>0.05)。
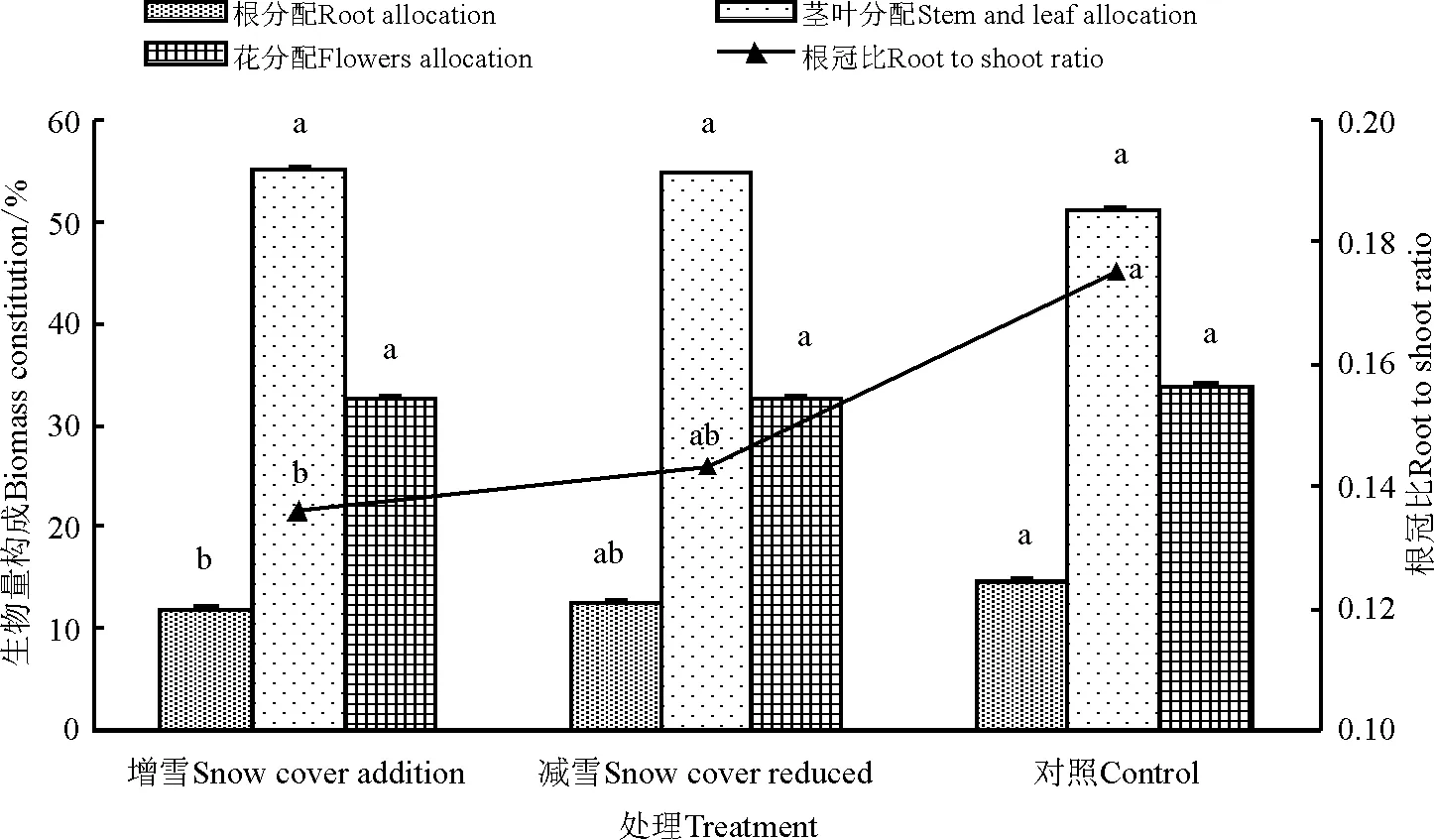
图2 覆雪处理下甘肃马先蒿根、茎和叶比重和根冠比Fig.2 The value of root,stem and leaves and root shoot ratio of Pedicularis kansuensis under snow cover change
注:不同小写字母表示同一项目不同处理间差异显著(P<0.05)。
Note: Different lower case letters in the same item mean significant difference among different treatments at 0.05 level.

表2 甘肃马先蒿个体形态特征对覆雪处理的响应Table 2 The response of the individual form feature about Pedicularis kansuensis for snow cover change
注:同列不同小写字母表示差异显著(P<0.05)。
Note:Different lower case letters within the same column mean significant difference among different treatments at 0.05 level.
3 讨论
3.1 覆雪处理下甘肃马先蒿及其寄主的物候
植物物候研究实质上是关于植物的生活史循环,以及生活史事件出现的时间与所处环境之间关系[31]的研究,植物生活史时间上的选择对植物自身的生存和繁殖成功至关重要[32]。
影响植物生长和发育所必需的有效积温是决定植物生长周期的重要因素之一,而覆雪厚度对植物萌发前的有效积温产生明显影响[7]。同时,雪层越薄融化的时间越早,该生境的局部小气候相对暖和,植物的生长季较长;雪层厚的地方融化时间比较晚,植物生长季节很短[7]。本研究表明,覆雪处理造成甘肃马先蒿及其寄主的生长季长短变化不一,这与前人的研究结果不太一致[7,33-34],可能是因为其空间尺度较大造成局部气候不一致,而本研究在大样方(100 m×100 m)内进行,融雪前覆雪层以上的气温基本无差异;同时,雪层厚度改变了甘肃马先蒿和寄主在萌发前的有效积温,影响融雪时间,从而使这些植物的萌发和返青时间乃至整个生长季做出调整。另外,在时间尺度上,其它研究大多是基于整个生长季增温而言,而本研究做覆雪处理只相当于影响其春季萌发前的局部气候条件,融雪后外界气温一致。
另外,覆雪处理还使得甘肃马先蒿及其寄主的不同物候期做出调整。植物春季物候与春季平均气温高度相关[35],也有研究认为与当月和上月的均温相关[36],而甘肃马先蒿种子萌发受温度变化的影响显著,表现出其萌发率和萌发速度随着温度升高而显著提高[25],故增雪处理相当于提升萌发前的有效积温,使其及寄主的春季物候提前,而减雪处理起到相反作用,这与其它研究基本一致[34,37]。甘肃马先蒿的物候也会受寄主的影响,因为寄生植物的形态大小、生长速率、光合作用、开花时间和开花树木等由寄主的不同而差异很大[38]。有研究表明,其更倾向于寄生禾本科植物[25],在萌发阶段马先蒿可能通过化感作用受寄主影响,故春季物候与寄主差异不大。积雪融化后,甘肃马先蒿及其寄主的外界生长条件(水分,养分、光照、气温)一致,区别在于它们对这些资源的争夺能力。
通过对比物候期起止时间还可以发现,甘肃马先蒿的返青期、营养期、甚至花期的开始时间都比寄主(蒲公英除外)对应的物候期要滞后,但从结实期开始逐渐提前,到种子散播期几乎完全领先其它寄主。可能是受寄主的营养输送限制,甘肃马先蒿在繁殖期之前的生长比寄主迟缓,但在生殖期由于前期的养分积累得以加速,甘肃马先蒿选择加快种子散播提前完成生活史不失为提高种群有效繁殖的一种方法。因为通过调节物候分化,使得自身和寄主或者其它物种在某个生长阶段的起始时间发生变化,从而在此阶段获得更强的竞争力。可以预见,在该区冬季降雪量增加的背景下,甘肃马先蒿通过调节物候变化使其和寄主产生差异,从而实现有效竞争。
3.2 覆雪处理下甘肃马先蒿的生长策略
植物生物量分配模式是植物对当前环境的适应性特征,体现物种生活史进化的结果[39]。植物生物量分配受水分[40]和海拔[41]影响显著。本研究结果显示,覆雪处理后甘肃马先蒿的繁殖分配和根冠比降低,增雪处理后茎叶分配有适度提高,但根分配显著降低,说明覆雪处理后甘肃马先蒿降低繁殖投资而增加营养投资。相关繁殖分配的大量研究认为,个体大小与繁殖分配存在异速关系,即植物个体越大,投入到花的资源所占比例越小[42],本研究结果与其一致。增雪处理后群落内的不同植物对土壤水分、养分和光资源争夺也进一步加强,其半寄生特性决定了甘肃马先蒿也可以自行进行光合作用补充营养,其生长策略是将更多资源支配到营养器官上,提高地上部分的生物量,以维持较大的有效光合面积,充分利用光能,进一步减少对寄主的营养输送的依赖,在繁殖前进行养分充分积累,最终在繁殖期领先其它物种,这与甘肃马先蒿及其寄主的物候期差异相契合。
植物表型特征的改变有利于植物个体在不同生境下的地生长和繁殖。甘肃马先蒿的表型特征可能受围封年限影响[16],本研究显示其表型特征也受覆雪影响。与自然降雪相比,增雪后甘肃马先蒿的株高和侧根尺寸更大、冠幅减小、花柱高度降低。对寄主而言,寄生植物会降低寄主的生物量,寄生植物影响寄主的生长繁殖和新陈代谢[22];同时,根部半寄生植物的吸器多产生于侧根[43],而侧根长度的增加为吸器的形成提供了更多的机会,能更好地从寄主上汲取养分,消耗寄主在开花前的营养积累,同时能更有效地争夺土壤的速效养分。
另外,甘肃马先蒿出现株高增大而冠幅减小的趋势,可能是由于增雪后群落内的生长环境更有利、各种植物对光热条件的竞争加剧造成的。花柱高度和分枝数决定了后期甘肃马先蒿用于繁殖的种子数量,显然,甘肃马先蒿降低花柱高度(繁殖分配)是为了利用更多的养分去保证实现更有效繁殖的种子质量。在近几十年当地冬季降雪量增加的背景下,甘肃马先蒿与寄主的特殊寄生关系影响到反映其生长和繁殖策略的个体形态,加上寄主个体形态的响应方式存在差异,可能引发各物种种群动态改变,最终改变群落组成,进而影响群落演替和生态系统功能。
4 结论
在研究区冬季平均降雪深±21.54%的覆雪处理下,甘肃马先蒿及其寄主的物候期对覆雪处理的响应不尽一致。覆雪处理使得甘肃马先蒿及其大部分寄主的花期物候期提前,秋季物候期推迟,但生长季的响应无一致规律,甘肃马先蒿的物候进程从结实期开始逐渐领先寄主。
另外,覆雪处理后甘肃马先蒿的繁殖分配和根冠比降低;在个体形态方面,增雪处理后甘肃马先蒿的相对高度、冠幅、花柱高度以及侧根尺寸与自然降雪对照相比均存在显著差异,而减雪处理后除了侧根其它各部分差异不大。总体而言,覆雪处理后甘肃马先蒿降低繁殖投资而增加营养投资。
增雪处理和减雪处理都使得甘肃马先蒿的物候期产生较大变化,但其生物量分配及个体形态对增雪处理的响应更明显。覆雪处理下甘肃马先蒿的物候、生物量分配以及个体形态特征的响应反映了其种群应对部分环境因子的改变,为保证种群繁衍而采取的有效竞争和繁殖策略。
References:
[1] 秦大河,Stocker T,259名作者,TSU(驻尼泊尔和北京).IPCC第五次评估报告第一工作组报告的亮点结论.气候变化研究进展,2014,10(1):1-6.
Qin D H,Stocker T,259 Authors,TSU(Ben & Beijing).Highlights of the IPCC Working Group I.Fifth assessment report.Advances in Climate Change Research,2014,10(1):1-6.(in Chinese)
[2] 胡列群,李帅,梁凤超.新疆区域近50 a积雪变化特征分析.冰川冻土,2013,35(4):793-800.
Hu L Q,Li S,Liang F C.Analysis of the variation characteristics of snow covers in Xinjiang region during recent 50 years.Journal of Glaciology and Geogcryology,2013,35(4):793-800.(in Chinese)
[3] 李效收,张明军,汪宝龙,王媛媛,王圣杰.天山地区冬季降雪量及其集中度和集中期的变化特征.资源科学,2012,34(8):1556-1564.
Li X S,Zhang M J,Wang B L,Wang Y Y,Wang S J.The change characteristics of winter snowfall,snow concentration degree and concentration period in the Tianshan Mountains.Resources Science,2012,34(8):1556-1564.(in Chinese)
[4] Waker M D,Ingersoll R C.Webber P J.Effects of inter annual climate variation on phenology and growth of two alpine forbs.Ecology,1995,76:1067-1083.
[5] Inouye D W,McGuire A D.Effects of snow pack on timing and abundance of flowering inDelphiniumnelsonii(Ranuncu laceae):Implications for climate change.American Journal of Botany,1991,78:997-1001.
[6] Billings W D.Arctic and alpine vegetation:Plant adaptions to cold summer climates.London:Arcticanil Alpine Environments,1974:403-443.
[7] 吴彦.季节性雪被覆盖对植物群落的影响.山地学报,2005,23(5):550-556.
Wu Y.Effects of seasonal snow cover on plant community.Journal of Mountain Science,2005,23(5):550-556.(in Chinese)
[8] Root T L,Price J T,Hall K R,Schneider S H,Rosenzweig C,Pounds J A.Fingerprints of global warming on wild animals and plants.Nature,2003,421:57-60.
[9] 莫非,赵鸿,王建永,强生才,周宏,王绍明,熊友才.全球变化下植物物候研究的关键问题.生态学报,2011,31(9):2593-2601.
Mo F,Zhao H,Wang J Y,Qiang S C,Zhou H,Wang S M,Xiong Y C.The key issues on plant phenology under global change.Acta Ecologica Sinica,2011,31(9):2593-2601.(in Chinese)
[10] Kramer K,Leinonen I,Loustau D.The importance of phenology for the evaluation of impact of climate change on growth of boreal,temperate and Mediterranean forests ecosystems:An overview.International Journal of Biometeorol,2000,44(2):67-75.
[11] Galen C,Stanton M L.Responses of snowbed plant species to changes in growing-season length.Ecology,1995,74:1546-1557.
[12] Wilson S D,Tilman D.Plant competition and resource availability inresponse to disturbance and fertilization.Ecology,1993,74:599-611.
[13] He J S,Bazzaz F A,Schmid B.Interactive effects of diversity,nutrients and elevated CO2on experimental plant communities.Oikos,2003,97:337-348.
[14] 钟章成.植物种群的繁殖对策.生态学杂志,1995,14(6):37-42.
Zhong Z C.Reproductive strategies of plant populations.Chinese Journal of Ecology,1995,14(6):37-42.(in Chinese)
[15] 宋宗水.巴音布鲁克草原生态恢复与综合治理已迫在眉睫.中国农业资源与区划,2006,27(1):21-25.
Song Z S.It is an urgent task to recover & comprehensively manage the grassland ecology of Bayinbuluke.Chinese Journal of Agricultural Resources and Regional Planning,2006,27(1):21-25.(in Chinese)
[16] 陈哲,周华坤,赵新全,温军,叶鑫,于龙.三江源区不同建植期人工草地甘肃马先蒿生物量分配.西北植物学报,2011,31(5):995-1002.
Chen Z,Zhou H K,Zhao X Q,Wen J,Ye X,Yu L.Resource allocation ofPediculariskansuensisof artificial grassland with different planting dates in Three Rivers source regions in Qinghai,China.Acta Botanica Boreali-Occidentalia Sinica,2011,31(5):995-1002.(in Chinese)
[17] 孟有儒.甘肃16种有毒杂草病害调查及利用其病害防、抑有毒杂草可能性评价.草业科学,2004,21(9):65-68.
Meng Y R.The disease investigation of 16 weed species and evaluation of their possibility in weeds control and inhibition.Pratacultural Science,2004,21(9):65-68.(in Chinese)
[18] 郁文彬,蔡杰,王红,陈建群.马先蒿属植物花冠分化与繁殖适应的研究进展.植物学通报,2008,25(4):392-400.
Yu W B,Cai J,Wang H,Chen J Q.Advances in floral divergence and reproductive adaptation inPedicularisL.(Orobanchaceae).Chinese Bulletin of Botany,2008,25(4):392-400.(in Chinese)
[19] 袁一斌,慕军鹏,彭幼红,孙书存.多齿马先蒿与刺齿马先蒿花部形态、昆虫访花频率及种子产量比较研究.应用与环境生物学报,2011,17(4):467-472.
Yuan Y B,Mu J P,Peng Y H,Sun S C.Comparative study on flower trait,pollinator visitation rate and seed production of twoPedicularisspecies.Chinese Journal of Applied and Environmental Biology,2011,17(4):467-472.(in Chinese)
[20] 柳妍妍,胡玉昆,于建梅,李凯辉,高国刚,王鑫.轮叶马先蒿危害现状及防除对策.干旱区研究,2008,25(6):778-782.
Liu Y Y,Hu Y K,Yu J M,Li K H,Gao G G,Wang X.Study on harmfulness ofPedicularismyriophyllaand its control measure.Arid Zone Research,2008,25(6):778-782.(in Chinese)
[21] 陈燕,管开云,李爱荣,隋晓琳.氮磷供给对两种马先蒿根系形态建成的影响.植物分类与资源学报,2014,36(1):56-64.
Chen Y,Guan K Y,Li A R,Sui X L.Effects of nitrogen and phosphorus supply on root morphology of twoPedicularisspecies.Plant Diversity and Resources,2014,36(1):56-64.(in Chinese)
[22] Duncan D C,Andy W,Janis A.Parasite-grass-forb interactions and rock-paper-scissor dynamics:Predicting the effects of the parasitic plant Rhinanthus minor on host plant communities.Journal of Ecology,2009,97:1311-1319.(in Chinese)
[23] 贺桂香,李凯辉,宋韦,公延明,刘学军,胡玉昆,田长彦.新疆天山高寒草原不同放牧管理下的CO2、CH4和NO2通量特征.生态学报,2014,34(3):1-8.
He G X,Li K H,Song W,Gong Y M,Liu X J,Hu Y K,Tian C Y.The fluxes of carbon dioxide,methane and nitrous oxide in alpine grassland of the Tianshan Mountain,Xinjiang.Acta Ecologica Sinica,2014,34(3):1-8.(in Chinese)
[24] 王鑫,胡玉昆,热合木都拉·阿迪拉,李凯辉,范永刚,柳妍妍.高寒草地主要类型土壤因子特征及对地上生物量的影响.干旱区资源与环境,2008,22(3):196-200.
Wang X,Hu Y K,Rehemudula·Adila,Li K H,Fan Y G,Liu Y Y.The character of soil factors of the main community in high and cold grasslands and influence on ground biomass.Journal of Arid Land Resources and Environment,2008,22(3):196-200.(in Chinese)
[25] 隋晓琳.甘肃马先蒿在新疆巴音布鲁克草原蔓延的生理生态学机理.北京:中国科学院大学硕士学位论文,2013.
Sui X L.Eco-physiological mechanism for the spatial expansion ofPediculariskansuensisin Bayanbulak grassland of Xinjiang.Master Thesis.Beijing:University of Chinese Academy of Sciences,2013.(in Chinese)
[26] 叶鑫,周华坤,刘国华,姚步青,赵新全.高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应.植物生态学报,2014,38(2):147-158.
Ye X,Zhou H K,Liu G H,Yao B Q,Zhao X Q.Responses of phenological characteristics of major plants to nutrient and water additions inKobresiahumilisalpine meadow.Chinese Journal of Plant Ecology,2014,38(2):147-158.(in Chinese)
[27] Rabinowitz D,Rapp J K,Sork V L,Rathcke B J,Reese G A,Weaver Jan C.Phenological properties of wind and insect pollinated prairie plants.Ecology,1981,62:49-56.
[28] 宛敏渭,刘秀珍.中国物候观测方法.北京:科学出版社,1979.
[29] 章志龙,刘旭东,贾鹏,杜国祯.氮添加对高寒草甸植物花期物候和群落结构的影响.草业科学,2013,30(5):728-735.
Zhang Z L,Liu X D,Jia P,Du G Z.Effects of nitrogen addition on flowering phenology,functional traits and community structure of alpine meadow in the eastern Qinghai-Tibet Plateau.Pratacultural Science,2013,30(5):728-735.(in Chinese)
[30] 牛克昌,赵志刚,罗燕江,杜国祯.施肥对高寒草甸植物群落组分种繁殖分配的影响.植物生态学报,2006,30(5):817-826.
Niu K C,Zhao Z G,Luo Y J,Du G Z.Fertilization effects on species reproductive allocation in an alpine meadow plant community.Journal of Plant Ecology,2006,30(5):817-826.(in Chinese)
[31] Fenner M.The phenology of growth and reproduction in plants.Perspectives in Plant Ecology,Evolution and Systematics,1998,1(1):78-91.
[32] Rathcke B,Lacey E P.Phenological patterns of terrestrial plants.Annual Review of Ecology and Systematics,1985,16:179-214.
[33] 白洁,葛全胜,戴君虎.贵阳木本植物物候对气候变化的响应.地理研究,2009,28(6):1606-1614.
Bai J,Ge Q S,Dai J H.Response of woody plant phenophases to climate change for recent 30 years in Guiyang.Geograghical Reasearch,2009,28(6):1606-1614.(in Chinese)
[34] Annette M.Plant phenological anomalies in germany and their relation to air tempreature and NAO.Climate Change,2003,57:243-263.
[35] 柳晶,郑有飞,赵国强,陈怀亮.郑州植物物候对气候变化的响应.生态学报,2007,27(4):1471-1479.
Liu J,Zheng Y F,Zhao G Q,Chen H L.Responses of phenology to climate change in Zhengzhou area.Acta Ecologica Sinica,2007,27(4):1471-1479.(in Chinese)
[36] 陈效逑,韩建伟.我国东部温带植物群落的季相及其时空变化特征.植物生态学报,2008,32(2):336-346.
Chen X Q,Han J W.Seasonal aspect stages of plant communities and its spatial temporal variation in temperate eastern China.Journal of Plant Ecology,2008,32(2):336-346.(in Chinese)
[37] 李荣平,周广胜,王玉辉,韩喜.羊草物候特征对气候因子的响应.生态学杂志,2006,25(3):277-280.
Li R P,Zhou G S,Wang Y H,Han X.Phenological responses ofLeymuschinesisto climate factors.Chinese Journal of Ecology,2006,25(3):277-280.(in Chinese)
[38] Victoria A B,Joseph E A.Resource limitation and the role of a hemiparasite on a restored prairie.Oecologia,2012,169:783-792.
[39] Boggs G.Resource allocation in variable environment:Comparing insects and plant.In:Bazzaz F A,Grace J.(eds).Plant Resource Allocation.San Diego,London:Academic Press,1997:73-91.
[40] 王银柱,王冬,刘玉,匡彦蓓,武高林.不同水分梯度下能源作物芒草和柳枝稷生物量分配规律.草业科学,2015,32(2):236-240.
Wang Y Z,Wang D,Liu Y,Kuang Y B,Wu G L.Biomass allocation ofMiscanthussinensisandPanicumvirgatumresponse to different water conditions.Pratacultural Science,2015,32(2):236-240.(in Chinese)
[41] 谢晓玲,周蓉,谢焕松,沈振西,邓自发.藏北高原不同海拔四川嵩草克隆种群表型和繁殖对策.草业科学,2014,31(12):2270-2276.
Xie X L,Zhou R,Xie H S,Shen Z X,Deng Z F.The clonal phenotypic variation and reproductive strategies ofKobresiasetchwanensisalong altitude in Northern Tibet.Pratacultural Science,2014,31(12):2270-2276.(in Chinese)
[42] 赵志刚,杜国祯,任青吉.5种毛茛科植物个体大小依赖的繁殖分配和性分配.植物生态学报,2004,28(1):9-16.
Zhao Z G,Du G Z,Ren J Q.Size-Dependent reproduction and sex allocation in five species of Ranunculaceae.Chinese Journal of Plant Ecology,2004,28(1):9-16.(in Chinese)
[43] Neumann U,Vian B,Weber H C,Salléb G.Interface between haustoria of parasitic members of the Scrophulariaceae and their hosts:A histochemical and immunocytochemical approach.Protoplasma,1999,207:84-97.
(责任编辑 王芳)
The response of biomass allocation and hosts phenology ofPediculariskansuensisto snow cover
Lu Jun1,2, Hu Yu-kun1
(1.Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China;2.University of Chinese Academy of Sciences, Beijing 100049, China)
In the present study, the response of biomass allocation ofPediculariskansuensisand the phenology ofP.kansuensisand its hosts to the snow cover changes with the level ±21.54 % for annual average snowfall depth in winter of alpine grassland in Bayinbuluke of Xinjiang were study to analyze their competition and reproductive strategies. Our results showed that the snow cover made the growing season ofP.kansuensisand hosts having no consistent change law compared to CK. The flowering phenology ofP.kansuensisand most typical hosts advanced under snow cover, which made their autumn phenology postponed. The beginning time of phenology period ofP.kansuensiskeep ahead of hosts from productive phase to seed dispersal. The reproductive allocation and root to shoot ratio ofP.kansuensissignificantly decreased(P<0.05). The plant height, canopy, column height and the size of lateral root ofP.kansuensisunder snow cover addition had significant difference with CK(P<0.05), however, except with lateral root, plants had no significant difference after snow cover removed. Totally, the same phenology stages ofP.kansuensisand its hosts had certain differentiation after snow cover change, while the biomass allocation and individual morphology were more sensitive to snow cover which reduced reproductive investment and enhanced nutrient investment under snow cover change
alpine grassland;Pediculariskansuensis; phenology; biomass allocation; snow cover
Hu Yu-kun E-mail:huyk@ms.xjb.ac.cn
10.11829/j.issn.1001-0629.2015-0691
2015-12-04 接受日期:2016-03-14
新疆联合基金(U1303201)第一作者:陆均(1989-),男,广西容县人,在读硕士生,研究方向为恢复生态学。E-mail:lj-081011@163.com
胡玉昆(1957-),男,山东淄博人,研究员,研究方向为恢复生态学。E-mail:huyk@ma.xjb.ac.cn
S812.1;Q945.79
A
1001-0629(2016)10-1933-09*
陆均,胡玉昆.甘肃马先蒿生物量分配及其寄主物候对覆雪的响应.草业科学,2016,33(10):1933-1941.
Lu J,Hu Y K.The response of biomass allocation and hosts phenology ofPediculariskansuensisto snow cover.Pratacultural Science,2016,33(10):1933-1941.
猜你喜欢
农业工程学报(2022年12期)2022-09-09
热带作物学报(2022年6期)2022-07-08
今日农业(2020年19期)2020-12-14
学生天地(2020年29期)2020-06-09
学生天地(2020年31期)2020-06-01
学生天地(2020年19期)2020-06-01
学生天地(2019年30期)2019-08-25
西南农业学报(2017年5期)2017-06-23
河北林业科技(2016年5期)2016-11-08
中国科技信息(2015年2期)2015-11-16
