
PNPLA3基因I148M多态性对肝癌细胞HepG2细胞周期及Cyclin D1、p53表达的影响
2016-12-05 06:05韩利蓉宋江勤郭飞波
山东医药 2016年40期
韩利蓉,宋江勤,郭飞波
(天门市第一人民医院,湖北天门431700)
PNPLA3基因I148M多态性对肝癌细胞HepG2细胞周期及Cyclin D1、p53表达的影响
韩利蓉,宋江勤,郭飞波
(天门市第一人民医院,湖北天门431700)
目的 探讨含patatin样磷脂酶域蛋白3(PNPLA3)基因I148M多态性对肝癌细胞HepG2细胞周期及Cyclin D1、p53表达的影响。方法 构建PNPLA3基因I148M突变型(突变组)、PNPLA3基因I148M野生型(野生组)的肝癌细胞HepG2,以空载体细胞作为对照组,采用流式细胞仪检测各组细胞周期,Western blotting、荧光定量PCR技术检测各组细胞Cyclin D1、p53蛋白及mRNA表达。结果 突变组G1期细胞所占比例明显低于野生组和对照组,S、G2/M期细胞所占比例明显高于野生组和对照组(P均<0.05);野生组与对照组G1、S、G2/M期细胞所占比例比较差异均无统计学意义(P均>0.05)。三组间Cyclin D1蛋白及mRNA相对表达量比较差异均无统计学意义(P均>0.05),突变组p53蛋白及mRNA相对表达量显著高于野生组和对照组(P均<0.05),而野生组和对照组比较差异均无统计学意义(P均>0.05)。结论 PNPLA3基因I148M多态性能够诱导p53过表达,导致肝细胞周期紊乱,最终发展成肝癌细胞。
肝癌;含patatin样磷脂酶域蛋白3;基因多态性;细胞周期;细胞周期蛋白D1;p53
国内外研究均证实,细胞周期阻滞与肿瘤的发生、发展密切相关[1,2]。含patatin样磷脂酶域蛋白3(PNPLA3)属于PNPLA家族成员之一,与传统激素敏感性脂肪酶相比,PNPLA家族成员N末端均有一个patatin区域,使得其在体内外均具有脂肪合成和分解功能,被认为是调节肝脏脂肪代谢的主要基因之一[3~6]。Rametta等[7]对人类染色体上PNPLA3基因分析发现,I148M多态性位点与肝脏脂肪含量升高有关。Hoekstra等[8]研究亦证实,PNPLA3基因I148M多态性与肝纤维化有关,且是肝癌发生的独立危险因素。由于脂肪代谢紊乱是多种肝病甚至肝癌的发病基础,推测PNPLA3基因I148M多态性可能与肝癌的发生、发展有关。2014年7月~2015年4月,我们观察了PNPLA3基因I148M多态性对人肝癌细胞HepG2细胞周期及Cyclin D1、p53表达的影响。现分析结果并报告如下。
1 材料与方法
1.1 材料 人肝癌细胞株HepG2和PNPLA3基因I148M突变型、PNPLA3基因I148M野生型、空载质粒转染的腺病毒,均购自上海士锋生物科技有限公司;RNA提取试剂盒、荧光定量PCR试剂盒,购自美国赛默飞世尔科技公司;蛋白裂解液、苯甲基磺酰氟(PMSF)、聚偏二氟乙烯(PVDF)膜、SDS-PAGE电泳分离蛋白均购自武汉博士德生物工程有限公司;β-actin抗体、辣根过氧化酶标记的二抗(羊抗鼠IgG抗体)、Cyclin D1单克隆抗体、p53单克隆抗体均购自美国圣克鲁斯生物技术公司;Cyclin D1、p53、β-actin引物由美国金斯瑞生物科技有限公司设计。ABI GeneAmp 9700 PCR仪、Odyssey成像系统及图像分析软件购自美国LI-COR公司。
1.2 细胞培养及腺病毒转染 取人肝癌细胞株HepG2置于DMEM培养液中,37 ℃、5% CO2细胞培养箱内培养。培养3天,待细胞融合≥80%时,加入0.25%胰蛋白酶0.5 mL消化,吸弃旧培养液,PBS洗涤3次,轻拍培养瓶使细胞自瓶壁脱落,混匀后加入新鲜培养基传代培养。取第4代对数生长期HepG2细胞,接种于6孔板中,每孔约2×105个,每孔设3个复孔。胰蛋白酶-EDTA消化后,加入RPMI 1640培养液,CO2培养箱中孵育12 h,分别加入含有PNPLA3基因I148M突变型腺病毒(突变组)、PNPLA3基因I148M野生型腺病毒(野生组)和空载质粒转染的腺病毒(对照组)3×107IU。转染24 h,加入聚凝胺使培养体系终浓度为6 mg/L,继续培养48 h,倒置荧光显微镜下观察并计算细胞转染效率。以倒置荧光显微镜下发现绿色荧光作为转染成功的标志。
1.3 细胞周期检测 取对数生长期HepG2细胞,PBS冲洗2次,移液枪吹打成细胞悬液并置于离心管中,室温下1 000 r/min离心10 min,PBS冲洗5 min×3次;滴加中性乙醇1 mL,4 ℃固定过夜。24 h后,室温下1 000 r/min离心10 min,PBS冲洗5 min×3次,吹打或细胞成悬液,加入适量碘化丙啶(PI,预先用PBS配制成1 mg/mL)和RNase,使其终浓度为50 mg/L,置于水浴锅上37 ℃温浴45 min。采用流式细胞仪检测细胞周期,每次5×104个细胞。重复检测3次,取平均值。
1.4 PNPLA3、Cyclin D1、p53蛋白检测 采用Western blotting技术。取对数生长期HepG2细胞,接种于6孔板,每孔加入含1% PMSF的细胞裂解液100 μL,待细胞完全裂解后,15 000 r/min离心5 min,留取上清液。将上清液移至EP管中,加入等量上样液,100 ℃煮沸5 min,使蛋白完全变性。SDS-PAGE电泳分离蛋白,并转移至PVDF膜上,100 mA、40 min电转后将PVDF膜取出。5%脱脂牛奶封闭2 h,加入1∶2 000稀释的PNPLA3单克隆抗体、1∶1 000稀释的Cyclin D1单克隆抗体和1∶500稀释的p53单克隆抗体,4 ℃孵育过夜;PBS冲洗3次,加入1∶10 000稀释的二抗,室温下震荡3 h,PBS冲洗3次;荧光图像扫描及条带灰度值分析。PNPLA3、Cyclin D1、p53蛋白相对表对量为样本条带与β-actin条带灰度值之比。
1.5 PNPLA3、Cyclin D1、p53 mRNA检测 采用荧光定量PCR技术。取对数生长期HepG2细胞,按照RNA提取试剂盒说明书提取细胞总RNA,并逆转录成cDNA。PCR扩增:PNPLA3上游引物:5′-CGGAAGACTGTGTCGAGCAA-3′,下游引物:5′-GGATCAAGCGCACTGTGGA-3′;Cyclin D1上游引物:5′-CCGTGTGCTAGCATAGCGC-3′,下游引物:5′-TGCTGGTCAATGTGGCTA-3′;p53上游引物: 5′-GACGTT-
CGGTGTCAGTCTGCC-3′,下游引物:5′-TGGCTGCGACTACATTAGGAC-3′;β-actin上游引物:5′-GTGCATGCTCGGCTGCGTCT-3′,下游引物:5′-GGTCTGAGGTTCAGCGTTGC-3′。PNPLA3扩增条件:96 ℃预变性1 min,95 ℃变性60 s,60 ℃退火30 s,72 ℃延伸60 s,35个循环后72 ℃延伸10 min;Cyclin D1扩增条件:95 ℃预变性5 min,95 ℃变性30 s,65 ℃退火45 s,72 ℃延伸1 min,40个循环后72 ℃延伸5 min。p53扩增条件:95 ℃预变性5 min,95 ℃变性15 s,65 ℃退火30 s,72 ℃延伸30 s,45个循环后72 ℃延伸5 min。记录各检测指标的CT值,重复检测3次,取平均值。以β-actin为内参,采用2-ΔΔCT法计算PNPLA3、Cyclin D1、p53 mRNA相对表达量。

2 结果
2.1 腺病毒转染HepG2细胞情况 携带不同基因型的腺病毒转染HepG2细胞48 h,倒置荧光显微镜下观察绿色荧光蛋白阳性表达率均高达90%,说明表达目的基因的细胞株构建成功。见插页Ⅲ图9。
2.2 各组细胞周期分布比较 见表1。

表1 各组细胞周期分布比较±s)
注:与对照组比较,*P<0.05;与野生组比较,#P<0.05。
2.3 各组PNPLA3、Cyclin D1、p53蛋白及mRNA表达比较 见表2。
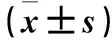
表2 各组PNPLA3、Cyclin D1、p53蛋白及mRNA相对表达量比较
注:与对照组比较,*P<0.05;与野生组比较,#P<0.05。
3 讨论
PNPLA3基因属于patatin样磷脂酶家族成员,具有非特异性水解酶的活性。PNPLA3基因多态性与人类非酒精性脂肪肝的遗传易感性密切相关。近年研究显示,PNPLA3基因多态性与多种肝脏疾病密切相关,其中I148M位点的多态性与肝脏脂肪含量显著升高有关[9]。由于这种脂肪升高不受遗传、体质量、原发病和乙醇摄入的影响,故有学者认为PNPLA3基因I148M多态性可能是导致肝脏病变的主要原因[10]。国外研究证实,PNPLA3基因I148M多态性不仅会增加肝脏脂肪含量,而且与肝脏组织学病变有关[11]。Pirazzi等[12]研究发现,PNPLA3基因I148M多态性与肝纤维化有关,并可能参与了肝细胞癌的发生。关于PNPLA3基因I148M多态性诱导肝脏病变的机制目前尚不清楚,但可以肯定的是与脂肪代谢有关。Burza等[13]研究发现,PNPLA3蛋白能够水解TG,其I148M位点多态性会导致水解酶的活性丧失,证实I148M多态性可能通过抑制PNPLA3的活性而导致TG分解受阻,而TG蓄积极易导致肝脏脂肪含量增加。对PNPLA3蛋白三维结构解析发现,其148位点氨基酸更换并未改变活性中心和空间结构,而是蛋氨酸的侧臂遮盖了活性中心与底物的结构,从而导致酶对TG水解活性降低[14]。但Valenti等[15]对小鼠PNPLA3基因敲除后并未获得小鼠脂肪肝模型,也未对小鼠体内TG的代谢造成影响。上述研究结果均提示PNPLA3基因I148M多态性对肝脏病变的影响作用机制较为复杂,需要深入研究。
细胞周期紊乱会导致细胞能量和物质代谢异常,最终引起细胞恶变[16,17]。Romeo等[18]认为,肝癌细胞周期明显异常,铂类等抗肿瘤药物的主要作用机制就是改变肿瘤细胞周期,诱导细胞发生凋亡。细胞周期主要有G1、S、G2、M期,其中G1/S期和G2/M期在维持细胞周期运转方面最为重要。当细胞通过G1/S期和G2/M期后,则无需依赖外界的分裂信号,能独立完成细胞周期[19]。因此G1/S期和G2/M期被认为是调控细胞周期的关键时期。Cyclin D1、p53均属于细胞周期调节蛋白,Cyclin D1是由人类CCND1基因所编码的一类蛋白质,其可通过结合并激活G1期特有的周期蛋白依赖性激酶CDK4,磷酸化G1期周期抑制蛋白(Rb),而磷酸化的Rb从其所结合的E2F转录因子上解离,从而推动细胞周期由G1期进入到S期。p53基因是细胞生长周期中的负调节因子,与细胞周期的调控、DNA修复、细胞分化、细胞凋亡等生物学功能有关。Cyclin D1、p53是参与调节G1/S期、G2/M期最主要的蛋白,Cyclin D1、p53表达失调与恶性肿瘤的发生、发展密切相关。Valenti等[20]研究证实,肝癌组织Cyclin D1、p53蛋白表达显著高于癌旁正常组织,随着治疗的好转,肝癌组织Cyclin D1、p53蛋白表达量逐渐降低。本研究结果显示,突变组p53 mRNA及蛋白表达量均高于野生组和对照组,提示PNPLA3基因多态性可能通过影响肝癌细胞p53表达,达到调节细胞周期的作用。本研究还发现,突变组G1期细胞所占比例低于野生组和对照组,S、G2/M期细胞所占比例高于野生组和对照组,提示I148M多态性可能与细胞周期紊乱有关。Sookoian等[21]研究证实,肝细胞周期加速后会导致细胞内能量代谢异常,并促进肝癌细胞的生长,且细胞周期的加速与肝癌细胞的恶性程度密切相关。本研究发现,突变组p53 mRNA蛋白相对表达量高于野生组和对照组,而三组细胞Cyclin D1蛋白和mRNA相对表达量比较差异均无统计学意义,提示PNPLA3基因I148M多态性可能通过上调p53表达而导致细胞周期紊乱,并促使其向肝癌发展。
综上所述,PNPLA3基因I148M多态性能够诱导p53过表达,导致肝细胞周期紊乱,最终发展为肝癌细胞。
[1] 詹启敏,陈杰.细胞周期与肿瘤转化医学[J].中国肿瘤临床,2014,41(1):1-7.
[2] Kitagawa M, Niisato N, Shiozaki A, et al. A regulatory role of K(+)-Cl(-) cotransporter in the cell cycle progression of breast cancer MDA-MB-231 cells[J]. Arch Biochem Biophys, 2013,539(1):92-98.
[3] Huang Y, He S, Li JZ, et al. A feed-forward loop amplifies nutritional regulation of PNPLA3[J]. Proc Natl Acad Sci U S A, 2010,107(17):7892-7897.
[4] He S, McPhaul C, Li JZ, et al. A sequence variation (I148M) in PNPLA3 associated with nonalcoholic fatty liver disease disrupts triglyceride hydrolysis[J]. J Biol Chem, 2010,285(9):6706-6715.
[5] Valenti L, Alisi A, Nobili V. Unraveling the genetics of fatty liver in obese children: additive effect of P446L GCKR and I148M PNPLA3 polymorphisms[J]. Hepatology, 2012,55(3):661-663.
[6] Zampino R, Coppola N, Cirillo G, et al. Abdominal fat interacts with PNPLA3 I148M, but not with the APOC3 variant in the pathogenesis of liver steatosis in chronic hepatitis C[J]. J Viral Hepat, 2013,20(8):517-523.
[7] Rametta R, Ruscica M, Dongiovanni P, et al. Hepatic steatosis and PNPLA3 I148M variant are associated with serum Fetuin-A independently of insulin resistance[J]. Eur J Clin Invest, 2014,44(7):627-633.
[8] Hoekstra M, Li Z, Kruijt JK, et al. The expression level of non-alcoholic fatty liver disease-related gene PNPLA3 in hepatocytes is highly influenced by hepatic lipid status[J]. J Hepatol, 2010,52(2):244-251.
[9] Pingitore P, Pirazzi C, Mancina RM, et al. Recombinant PNPLA3 protein shows triglyceride hydrolase activity and its I148M mutation results in loss of function[J]. Biochim Biophys Acta, 2014,1841(4):574-580.
[10] Larrieta-Carrasco E, Acua-Alonzo V, Velázquez-Cruz R, et al. PNPLA3 I148M polymorphism is associated with elevated alanine transaminase levels in Mexican Indigenous and Mestizo populations[J]. Mol Biol Rep, 2014,41(7):4705-4711.
[11] Zampino R, Florio A, Coppola N, et al. PNPLA3 I148M variant as a risk factor for carotid atherosclerosis in chronic hepatitis C[J]. Int J Cardiol, 2014,172(1):291-292.
[12] Pirazzi C, Adiels M, Burza MA, et al. Patatin-like phospholipase domain-containing 3 (PNPLA3) I148M (rs738409) affects hepatic VLDL secretion in humans and in vitro[J]. J Hepatol, 2012,57(6):1276-1282.
[13] Burza MA, Pirazzi C, Maglio C, et al. PNPLA3 I148M (rs738409) genetic variant is associated with hepatocellular carcinoma in obese individuals[J]. Dig Liver Dis, 2012,44(12):1037-1041.
[14] Nobili V, Bedogni G, Donati B, et al. The I148M variant of PNPLA3 reduces the response to docosahexaenoic acid in children with non-Alcoholic fatty liver disease[J]. J Med Food, 2013,16(10):957-960.
[15] Valenti L, Dongiovanni P, Ginanni Corradini S, et al. PNPLA3 I148M variant and hepatocellular carcinoma: a common genetic variant for a rare disease[J]. Dig Liver Dis, 2013,45(8):619-624.
[16] 周珏宇,石嵘,郑文岭,等.microRNA与肿瘤细胞周期的关系及其研究方法进展[J].基础医学与临床,2013,33(1):9-14.
[17] 李维妙,夏靖华,冯征,等.肿瘤高表达细胞周期相关蛋白在非小细胞肺癌中的表达及临床意义[J].细胞与分子免疫学杂志,2015,31(5):668-671.
[18] Romeo S, Sentinelli F, Dash S, et al. Morbid obesity exposes the association between PNPLA3 I148M (rs738409) and indices of hepatic injury in individuals of European descent[J]. Int J Obes (Lond), 2010,34(1):190-194.
[19] Valenti L, Nobili V, Al-Serri A, et al. The APOC3 T-455C and C-482T promoter region polymorphisms are not associated with the severity of liver damage independently of PNPLA3 I148M genotype in patients with nonalcoholic fatty liver[J]. J Hepatol, 2011,55(6):1409-1414.
[20] Valenti L, Colombo M, Fargion S. Modulation of the effect of PNPLA3 I148M mutation on steatosis and liver damage by alcohol intake in patients with chronic hepatitis C[J]. J Hepatol, 2011,55(6):1471-1472.
[21] Sookoian S, Pirola CJ. Meta-analysis of the influence of I148M variant of patatin-like phospholipase domain containing 3 gene (PNPLA3) on the susceptibility and histological severity of nonalcoholic fatty liver disease[J]. Hepatology, 2011,53(6):1883-1894.
10.3969/j.issn.1002-266X.2016.40.019
R735.7
B
1002-266X(2016)40-0058-04
2016-05-26)
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
天津医科大学学报(2019年3期)2019-08-13
国际呼吸杂志(2019年4期)2019-03-12
中华老年多器官疾病杂志(2016年7期)2016-04-28
西南农业学报(2016年6期)2016-04-16
癌症进展(2016年10期)2016-03-20
肿瘤预防与治疗(2015年1期)2015-09-26
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
医学研究杂志(2015年5期)2015-06-10
中国当代医药(2015年16期)2015-03-01
