
New insights into the functions of PtdIns(3,5)P2in the pathogenisis of neurodegenerative disorders
2016-12-02 02:29FuminoriTsuruta
中国神经再生研究(英文版) 2016年2期
PERSPECTIVE
New insights into the functions of PtdIns(3,5)P2in the pathogenisis of neurodegenerative disorders
The membrane trafficking systems in brain play an important role in the regulation of neuronal processes, such as morphology, neuronal survival and synaptic plasticity. It has been suggested that the phosphatidylinositols (PIs) located on endolysosomal membranes play a key role in controlling this trafficking systems. Recently, we have reported that PtdIns(3,5)P2, which is a low abundant PIs in cells, is involved in the regulation of lysosomal degradation via vesicle transport (Tsuruta et al., 2009; Tsuruta and Dolmetsch, 2015). In this perspective, we provide an overview of the general functions of PtdIns(3,5)P2and discuss their potential role in the pathogenesis of neurodegenerative disorders.
The cellular functions of PtdIns(3,5)P2: PIs are crucial for the regulation of a wide variety of cellular processes. The hydroxyl groups of the inositol ring of PIs is phosphorylated and dephosphorylated by specific phosphatidylinositol kinases and phosphatases. These enzymes control the amount of intracellular PIs at appropriate levels and regulate specific cellular functions. Recently, several groups have actively investigated PtdIns(3,5)P2, which is less abundant than most other PIs, such as PtdIns(4,5)P2. PtdIns(3,5)P2is predominantly localized in endosome and lysosomes, where it is involved in regulating multivesicular bodies (MVBs) invagination, autophagy, and membrane trafficking (Shisheva, 2012; McCartney et al., 2014). Intriguingly, PtdIns(3,5)P2levels are dramatically elevated after osmotic stresses and return to the basal level within a half hour. Because this reaction is dynamic, the understanding of PtdIns(3,5)P2turnover may provide important cues into the role of lipids in regulating neuronal functions. PtdIns(3,5)P2is known to be generated from PtdIns(3)P by phosphatidylinositol 3-pholsphate 5-kinase (Fab1; yeast or PIKfyve; mammals), the only lipid kinase in most eukaryotes. Fab1/PIKfyve is thought to be involved in the regulation of membrane homeostasis, and defect in its function lead to abnormal vacuole formation in both yeast and mammalian cells. Moreover, loss of the PIKfyve gene is embryonically lethal, suggesting that PtdIns(3,5)P2produced by Fab1/PIKfyve plays a vital role in maintaining membrane homeostasis. The PtdIns(3,5)P2was initially reported to be implicated in the regulation of ESCRTIII complex. After phosphorylation of PtdIns(3)P by Fab1 in yeast, PtdIns(3,5)P2associates with Vps24, which is an ESCRTIII subunit, followed by the recruitment of additional subunits of ESCRTIII to sort cargo proteins into MVBs. Another effector called Ent3p functions in MVBs sorting via an ENTH domain, which is a phosphatidylinositol phosphate-binding motif. However, the molecular mechanisms by which PtdIns(3,5) P2regulates MVBs is still controversial (Michell et al., 2006). Additionally, PtdIns(3,5)P2has been implicated in the autophagy pathway. Atg18/WIPIs (Atg18; yeast or WIPI1, WIPI2, WIPI3, and WIPI4; mammals), which are essential proteins in autophagy, recognize both PtdIns(3,5)P2and PtdIns(3)P. Interestingly, it was reported that Atg18 negatively regulates PtdIns(3,5)P2level in yeast. Moreover, PtdIns(3,5)P2has been suggested to be a potential sensor of intracellular amino acids, followed by the regulation of target of rapamycin complex 1 (TORC1) activity (Jin et al., 2014). These observations imply that both Atg18/WIPIs and PtdIns(3,5)P2are mediators that link amino acids levels to TORC1 signaling in cells. PtdIns(3,5)P2regulates vesicle motility: Recently, we have found a novel functions of PtdIns(3,5)P2in neurons. Previously, we reported that PtdIns(3,5)P2produced by PIKfyve plays a significant role in the protection of excitotoxic neuronal cell death via regulation of the levels of the voltage-gated calcium channel CaV1.2 (Tsuruta et al., 2009). In neurons, intracellular Ca2+concentration is responsible for the coordination of neuronal activity being tightly regulated through control of both N-methyl-D-aspartate receptor (NMDAR) and voltage-gated calcium channel, CaV1.2. Our studies indicated that bath glutamate application promotes the internalization of CaV1.2 channels from the plasma membrane leading to lysosomal degradation. Under this condition, PIKfyve associates with CaV1.2 and synthesizes PtdIns(3,5)P2on CaV1.2-containing vesicles, resulting in the effective transition from endosomes to lysosomes. Notably, inhibition of PIKfyve, which causes a decrease in PtdIns(3,5)P2levels, suppresses glutamate-induced internalization of surface CaV1.2, leaving neurons vulnerable to excitotoxicity. As defects in both membrane trafficking and Ca2+homeostasis are implicated in many neurodegenerative disorders, PIKfyve may link between impaired trafficking systems and neurological disorders. The mechanism by which PIKfyve mediates lysosomal degradation of cargo proteins in neurons has been a long-standing unsolved question. In our recent study, we were able to shed light on the function of PIKfyve (Tsuruta and Dolmetsch, 2015). We observed that endolysosomal compartments are highly motile in neuronal dendrites. Thus, we speculated that PtdIns(3,5)P2are involved in the regulation of vesicle motility. In fact, knockdown of PIKfyve in neurons significantly decreased a motility of both endosomes and lysosomes, demonstrating that PIKfyve is required to control their motility in neurons.
The remaining question is the molecular mechanisms by which PtdIns(3,5)P2mediates vesicle movements in neurons. So far, several studies have reported that other PIs on endolysosomal compartments are associated with their own motility (Hirokawa et al., 2009). For instance, kinesin-3 family proteins, KIF1A and KIF1Bβ have a PH domain located at the C-terminal tail, which directly interacts with PtdIns(4,5)P2and regulates the motility of PtdIns(4,5)P2-containing vesicles, including synaptic vesicles in neurons. In addition, guanylate kinase-associated kinesin (GAKIN), classified as a kinesin-3 protein KIF13B, interacts with PtdIns(3,4,5)P3-containing vesicles and transports them to the edge of axons, followed by selective activation of PI3K pathways. Furthermore, the kinesin-3 family member KIF16B directly binds to PtdIns(3)P on early endosomes and transports early endosomes to the microtubule plus ends. Hence, it is plausible that PIs function as a recognition target of either motor proteins or their adaptor proteins, and that PtdIns(3,5)P2could be associated with either motor proteins or adaptor proteins, which mediates vesicle transport in neurons (Figure 1).
PIKfyve might also control membrane trafficking through the binding proteins. Recent studies have shown that members of JNK-interacting proteins (JIPs) are involved in axonal transport regulated by KIF5 proteins. Mutation in JIP homologs in both Drosophila and Caenorhabditis elegans impair axonal transport. JIP4, a novel member of the JIP family, binds to PIKfyve, and is involved in the regulation of membrane trafficking (Ikonomov et al., 2009). Therefore, PIKfyve binding proteins such as JIP4 could control vesicle movements in neurons. Apart from JIP4, other target proteins that alter the functions of either motor or adaptor proteins are thought to be potent candidates. It has been known that Fab1/PIKfyve activity is regulated by protein complex formation on vesicle, including Fig4 (Fig4; yeast or Fig4/Sac3; mammals) and Vac14 (Vac14; yeast or Vac14/Ar-PIKfyve; mammals). Fig4, which is a lipid phosphatase, dephosphorylates PtdIns(3,5) P2at 5-position and synthesizes PtdIns(3)P. Interestingly, Fig4 also functions as a positive regulator of PIKfyve, demonstrating that not only PIKfyve but also Fig4 is essential to adjust the balance between PtdIns(3,5)P2and PtdIns(3)P on vesicles. Vac14 is composed of many HEAT domains and acts as a scaffold to form theprotein complex. As PtdIns(3,5)P2turnover is dynamic and rapid, these protein complexes may monitor PIs levels on vesicles spatiotemporally and activate downstream signaling pathway, resulting in the alternation of motor protein functions. Taken together, it is possible that PtdIns(3,5)P2on vesicles is associated with the function of motor protein either directly or indirectly.
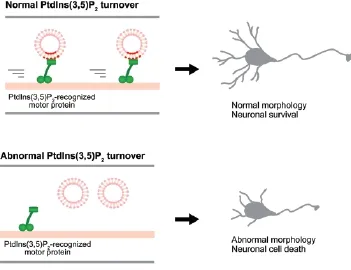
Figure 1 Putative model for the mechanisms that underlie PtdIns(3,5)P2-dependent vesicle motility.
Defect of PtdIns(3,5)P2effectors and neurodegeneration: Recent study have demonstrated that mutation in Fig4 causes neurological disorders (Chow et al., 2007), such as Charcot-Marie-Tooth disease and amyotrophic lateral sclerosis (ALS). Presently, it is known that two candidates, WIPI4 and SNX14, are the PtdIns(3,5)P2effector proteins involved in neurodegeneration. Mutations in WIPI4, the mammalian homolog of Atg18, were reported to be associated with a severe neurodegenerative disorder-Static encephalopathy of childhood with neurodegeneration in adulthood (SENDA). SENDA is a genetic disorder characterized by abnormal accumulation of iron in the basal ganglia (Saitsu et al., 2013). Recently, several groups have reported that accumulation of ferritin clusters, iron storage structures, is observed in autophagy-deficient cells. Furthermore, NCOA4, a ferritinopathy specific autophagy cargo receptor associated with ferritin heavy chain 1, was identified. These findings implied that defects in autophagy causes impairment of ferritinophagy and abnormal accumulation of iron in cells (Kaur and Debnath, 2015). In addition, recent exome sequencing studies revealed that mutation in SNX14, which is part of a family of sorting nexins, is associated with cerebellar atrophy (Akizu et al., 2015). SNX14 contains a PX domain, which recognizes PtdIns(3,5)P2on endolysosomal membrane. Defect in SNX14 exhibits large lysosomes and impairs effective autophagosome clearance. As the SNX family consists of a diverse group of membrane trafficking proteins, impaired PtdIns(3,5)P2may either affect the sorting ability of SNX14 or the properties of endolysosomal compartments. Interestingly, the phenotype showing larger lysosome was similar to that of PIKfyve knockdown in our experiments, implying that SNX14 could be a target of PtdIns(3,5)P2associated with the regulation of vesicle motility.
Conclusions: The PtdIns(3,5)P2turnover are crucial for maintaining the neuronal functions including vesicle transport. Aberrant PtdIns(3,5)P2metabolism leads to neurodegenerative disorders. As defects in either PIKfyve or Fig4 expression lead to abnormal neuronal properties including neuronal morphology and survival, misregulation of PtdIns(3,5)P2may alter the normal tuning of neural circuit and brain homeostasis. Although the molecular mechanism that connect impaired PtdIns(3,5)P2to pathogenesis of neurodegenerative disorders has not been elucidated, this question is important and will need to be addressed in the future. Furthermore, it might be interesting to investigate whether control of PtdIns(3,5)P2levels by pharmacological compounds in neurons are effective against several diseases such as Charcot-Marie-Tooth disease and ALS.
We apologize to the many authors whose papers could not be cited due to space limitations. I would like to thank Dr. Ricardo Dolmetsch and the members of his laboratory for their helpful discussion, Drs. Sergiu Pasca, Yu Kigoshi, and Takuma Aihara for critical reading of this paper.
Fuminori Tsuruta*
Graduate School of Life and Environmental Sciences, University of Tsukuba, Ibaraki, Japan
*Correspondence to: Fuminori Tsuruta, Ph.D., tsuruta.fuminori.fn@u.tsukuba.ac.jp.
Accepted: 2015-11-16
Akizu N, Cantagrel V, Zaki MS, Al-Gazali L, Wang X, Rosti RO, Dikoglu E, Gelot AB, Rosti B, Vaux KK, Scott EM, Silhavy JL, Schroth J, Copeland B, Schaffer AE, Gordts PL, Esko JD, Buschman MD, Field SJ, Napolitano G, et al. (2015) Biallelic mutations in SNX14 cause a syndromic form of cerebellar atrophy and lysosome-autophagosome dysfunction. Nat Genet 47:528-534.
Chow CY, Zhang Y, Dowling JJ, Jin N, Adamska M, Shiga K, Szigeti K, Shy ME, Li J, Zhang X, Lupski JR, Weisman LS, Meisler MH (2007) Mutation of FIG4 causes neurodegeneration in the pale tremor mouse and patients with CMT4J. Nature 448:68-72.
Hirokawa N, Noda Y, Tanaka Y, Niwa S (2009) Kinesin superfamily motor proteins and intracellular transport. Nat Rev Mol Cell Biol 10:682-696.
Ikonomov OC, Fligger J, Sbrissa D, Dondapati R, Mlak K, Deeb R, Shisheva A (2009) Kinesin adapter JLP links PIKfyve to microtubule-based endosome-to-trans-Golgi network traffic of furin. J Biol Chem 284:3750-3761.
Jin N, Mao K, Jin Y, Tevzadze G, Kauffman EJ, Park S, Bridges D, Loewith R, Saltiel AR, Klionsky DJ, Weisman LS (2014) Roles for PI(3,5)P2 in nutrient sensing through TORC1. Mol Biol Cell 25:1171-1185.
Kaur J, Debnath J (2015) Autophagy at the crossroads of catabolism and anabolism. Nat Rev Mol Cell Biol 16:461-472.
McCartney AJ, Zhang Y, Weisman LS (2014) Phosphatidylinositol 3,5-bisphosphate: low abundance, high significance. BioEssays 36:52-64.
Michell RH, Heath VL, Lemmon MA, Dove SK (2006) Phosphatidylinositol 3,5-bisphosphate: metabolism and cellular functions. Trends Biochem Sci 31:52-63.
Saitsu H, Nishimura T, Muramatsu K, Kodera H, Kumada S, Sugai K, Kasai-Yoshida E, Sawaura N, Nishida H, Hoshino A, Ryujin F, Yoshioka S, Nishiyama K, Kondo Y, Tsurusaki Y, Nakashima M, Miyake N, Arakawa H, Kato M, Mizushima N, et al. (2013) De novo mutations in the autophagy gene WDR45 cause static encephalopathy of childhood with neurodegeneration in adulthood. Nat Genet 45:445-449, 449e441.
Shisheva A (2012) PIKfyve and its Lipid products in health and in sickness. Curr Top Microbiol Immunol 362:127-162.
Tsuruta F, Dolmetsch RE (2015) PIKfyve mediates the motility of late endosomes and lysosomes in neuronal dendrites. Neurosci Lett 605:18-23.
Tsuruta F, Green EM, Rousset M, Dolmetsch RE (2009) PIKfyve regulates CaV1.2 degradation and prevents excitotoxic cell death. J Cell Biol 187:279-294.
10.4103/1673-5374.177727 http://www.nrronline.org/
How to cite this article: Tsuruta F (2016) New insights into the functions of PtdIns(3,5)P2in the pathogenisis of neurodegenerative disorders. Neural Regen Res 11(2):240-241.
- 中国神经再生研究(英文版)的其它文章
- Tissue-type plasminogen activator is a modulator of the synaptic vesicle cycle
- Impaired consciousness caused by injury of the lower ascending reticular activating system: evaluation by diffusion tensor tractography
- Considering calcium-binding proteins in invertebrates: multi-functional proteins that shape neuronal growth
- Cardiovascular dysfunction following spinal cord injury
- Practical application of the neuroregenerative properties of ketamine: real world treatment experience
- Exergames: neuroplastic hypothesis about cognitive improvement and biological effects on physical function of institutionalized older persons