
Precipitation as a control of vegetation phenology for temperate steppes in China
2016-11-23 02:37:02ZHOUYaoZhiandJIAGenSuo
ZHOU Yao-Zhiand JIA Gen-Suo
aInstitute of Tibetan Plateau Ecology, Agricultural and Animal Husbandry College of Tibet University, Nyingchi, Tibet, China;bKey Laboratory of Regional Climate—Environment for East Asia, Institute of Atmospheric Physics, Chinese Academy of Sciences, Beijing, China
Precipitation as a control of vegetation phenology for temperate steppes in China
ZHOU Yao-Zhia,band JIA Gen-Suob
aInstitute of Tibetan Plateau Ecology, Agricultural and Animal Husbandry College of Tibet University, Nyingchi, Tibet, China;bKey Laboratory of Regional Climate—Environment for East Asia, Institute of Atmospheric Physics, Chinese Academy of Sciences, Beijing, China
Using the NDVI ratio method, the authors extracted phenological parameters from NOAA-AVHRR NDVI time-series data (1982—2008). The start of the growing season (SOS) and the date of maximum NDVI (Peak-t) correlated signifcantly with the mean annual precipitation along regional gradients of the steppes. Along the south transect (located at a lower latitude with a higher annual mean temperature) there was a positive correlation between the end of the growing season (EOS) and the mean annual precipitation along precipitation gradients (R2= 0.709, p < 0.0001). However,along the north transect (located at higher latitude with lower annual mean temperature), the EOS was slightly negatively related with the mean annual precipitation (R2= 0.179, p < 0.1). There was positive correlation between the length of the growing season and the annual precipitation along two transects (R2= 0.876, p < 0.0001 for the south transect; R2= 0.290, p < 0.01 for the north transect). Thus, for the Inner Mongolian steppe, it is precipitation rather than temperature that determines the date of the SOS.
ARTICLE HISTORY
Revised 30 December 2015
Accepted 1 February 2016
Phenology; precipitation;
steppe; remote sensing
已有的物候研究主要针对温度的影响,而降水量的物候效应研究相对较少。文中用NDVI比率法分析内蒙古草原两个同温样带的年降水量与草原物候之间的关系。结果表明,在中国温带草原区,年降雨量增加能显著促进春季植物的萌芽和夏季最高生物量日期的到来,而年平均温度起抑制的作用。在低纬度的同温样带(内蒙古中部草原),年降水量增加能延迟生长期结束日期,而在高纬度地区(呼伦贝尔草原)年降水量增加会轻微提前结束生长期。因此在内蒙古中部草原年降水量增加能明显延长草原植被的生长期。
1. Introduction
Climate change over the last several decades has substantially altered the arrival of biological spring and winter (Brown and de Beurs 2010). The changes in the annual cycles of plants and the lengthening of the growing season have had considerable infuence on natural and social systems (Morisette et al. 2009). Moreover, shifts in vegetative phenology have, in turn, fed back to climate change through vegetation—atmosphere interactions (Finzi et al. 2011). Vegetative phenology, therefore, is an important dimension of ecological research for understanding future global change (Cong et al. 2012)
On regional and global scales, temperature and precipitation have been widely documented as major factors infuencing vegetative phenology (Morisette et al. 2009). Previous research has focused on the efects of temperature on vegetative phenology, and temperature is widely considered to be a dominant control over phenology. However, more research is needed into the efects of other important factors on vegetative phenology, especially precipitation in arid and semi-arid regions (Cleland et al. 2006). So far, we have very limited understanding of the infuence that complex environmental factors have on shifts in key vegetative phenological parameters. This limited understanding has hindered simulations and predictions of future changes in vegetative phenology (Richardson et al. 2012). As a result,spatial heterogeneity of shifting vegetation phenology and its key drivers are so far poorly understood.
Steppes in arid and semi-arid regions are more sensitive to climate variations, especially precipitation shifts,than many other terrestrial ecosystems (Knapp and Smith 2001). Precipitation is considered as the dominant factor that determines the emergence of green leaves, the duration of the vegetation growing season, and the primary production in arid and semi-arid areas (Soudani et al. 2012). Therefore, investigating the efects of precipitationon vegetative phenology in steppes is critical to better understand the seasonal interactions between ecosystems and the atmosphere.
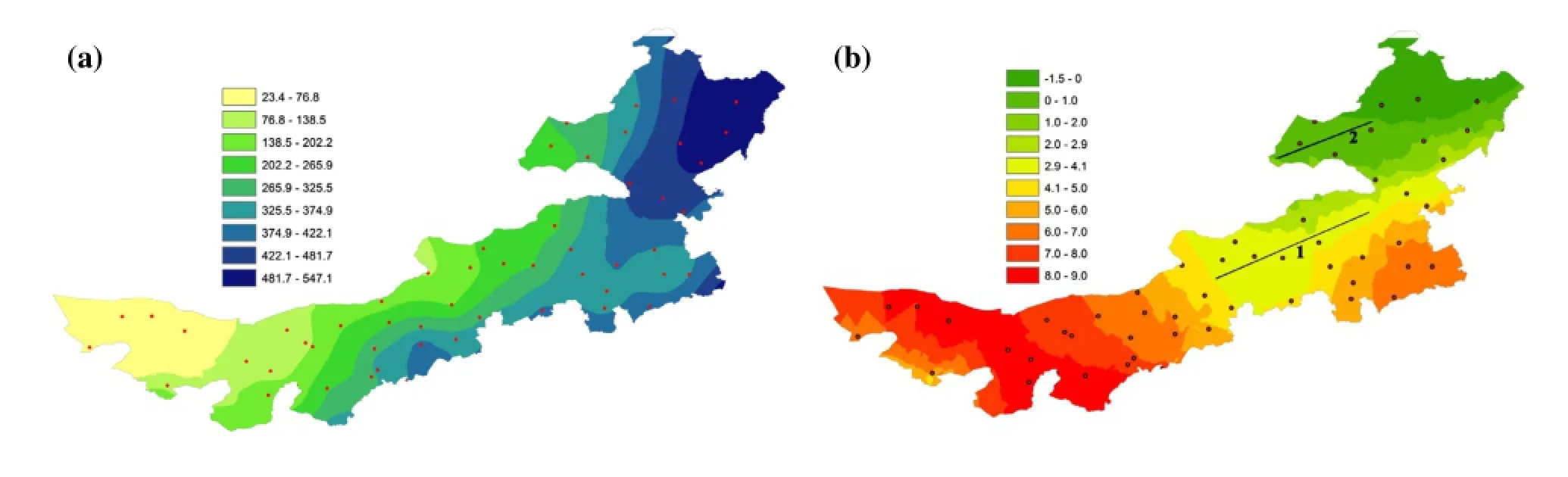
Figure 1.Mean annual (a) precipitation (units: mm) and (b) temperature (units: °C) in Inner Mongolia Autonomous Region steppes.
Regional environmental transects facilitate a unique approach to explore ideas and models about ecosystem processes, to assess their level of generality, and to project them at the global scale (Burke 2000). However, since most environmental factors (e.g. temperature and precipitation)that control ecosystem processes vary together across landscapes, it is difcult to isolate the infuence of specifc factors.
The climate of China features a summer monsoon and a strong annual cycle caused by the diferences in heat reserves between the world's largest continent (Eurasia)and the world's largest ocean (the Pacifc). For the east and central Inner Mongolian Steppe (IMS), the presence of an annual monsoon causes temperature gradients to vary along latitude, and for precipitation gradients to vary along longitude (Figure 1). This provides an opportunity to isolate the infuences of temperature and precipitation,the two main controlling factors of vegetative phenology.
The paper focuses on the efects of precipitation on steppe vegetative phenology. In carrying out the study,we expected that, in areas of similar temperature: (1) there would be negative correlation between the start of the growing season (SOS) and both Peak-t (the date of peak NDVI) and mean annual precipitation; and (2) that an increase in precipitation would cause a delay in the end of the growing season (EOS) and an increase in the length of the growing season (LOS).
2. Data and methods
2.1. Study area
We chose the temperate steppe (the east and center of Inner Mongolia Autonomous Region) in China as the study area. The majority of annual precipitation (100—400 mm) falls during the summer (June—September), coinciding with peak high temperatures. Severe drought frequently occurs in spring and early summer. In the two study areas(the north transect and the south transect), the main plant species include Stipa grandis, Stipa krylovii, Leymus chinensis, Agropyron cristatum, Artemisia frigida, Cleistogenes squarrosa, Koeleria cristata, Poa sphondylodes, and Carex duriuscula. The topsoil in the two study areas is chestnut soil. The dominant plant species in the part of the two study areas close to Daxing'an Mountains include Stipa baicalensis, Carex pediformis, and Spiraea salici olia; and in the part of the two study areas close to desertifed steppe they include S. krylovii, Allium polyrhizum, and Allium mong olicum. Detailed information about the study area is available in Lee et al. (2002) and Yu et al. (2003).
We calculated annual mean temperature and precipitation of all 52 weather stations in Inner Mongolia, and then generated gridded mean annual precipitation and mean annual temperature maps for the study region using the Kriging interpolation method over meteorological records(Figure 1). The annual mean temperatures along the south transect and along the north transect were 2.9—4.1 °C and 0—1.0 °C, respectively. The south transect, located in the Xilingol steppe, had an annual mean precipitation of 130—400 mm, while the annual mean precipitation along the north transect, located in the Hulunbeier steppe, ranged from 230 to 360 mm. Along each transect, the mean annual temperature had little spatial variation (0—1.0 °C for the north transect and 2.9—4.1 °C for the south transect),while the annual precipitation varied greatly (Figure 1(a)). This made it possible to analyze the efects of annual precipitation on vegetative phenology without the efects of temperature. Considering relatively homogeneous mean annual temperature within each of the two transects, we used data from 14 meteorological stations in the IMS, and vegetation pixels close to those weather stations (distance of less than 15 km), to examine the relationship betweenthe vegetation phonological parameters and mean annual temperature.
2.2. Data
Long-term satellite records of vegetation greenness at a spatial resolution of 8 km × 8 km and a 15-day interval were acquired from the global inventory monitoring and modeling studies (GIMMS) group derived from the NOAA/AVHRR series satellites during 1982—2008. The GIMMS NDVI data have been corrected to remove some calibration errors caused by sensor degradation,clouds, and stratospheric aerosol loadings from volcanic eruptions (Tucker et al. 2001). Climate datasets came from the China Meteorological Administration, and included 27 years (1982—2008) of continuous records from 52 weather stations throughout Inner Mongolia Autonomous Region. The objective of the study is to examine the response of key vegetation phenological parameters to annual precipitation and mean temperature, in order to identify controlling factors behind changing seasonality over the IMS.
2.3. Methods
Phenological analysis of a small area consisting of homogenous vegetation can avoid the infuence of mixed signals of natural vegetation, crop farming, urbanization, and forestation on the phenological measurements of vegetation. Previous studies on vegetative phenology based on remote sensing have used homogeneous subsets (4—9 pixels) of NDVI data in order to achieve better estimates of phenological parameters (de Beurs and Henebry 2005; Jia,Epstein, and Walker 2003; Jobbagy, Sala, and Paruelo 2002).
We selected our subset areas and discarded pixels occupied by forest, or that contained a high percentage of wetlands, water bodies, cities, farmlands or rock outcrops with fner resolution satellite data. We obtained average NDVI values from the two study areas along the two transect lines in Figure 1(a). The sampling area was 256 km2.
In order to quantify changes in the vegetative phenology of the IMS along precipitation gradients, we obtained the mean annual precipitation values for each 8 km × 8 km pixel using available precipitation records and geographical interpolation models calibrated for the whole study region.
Various methods have been proposed for retrieving phenology parameters from the NDVI time series data. In our study, we used NDVI ratios because they are considered the simplest and most efective method for phenological studies (White et al. 2009; Yu, Luedeling, and Xu 2010).
According to previous studies (Chen and Li 2009; Gu et al. 2012) and climate data (daily temperature and precipitation records during 1982—2008) for the IMS, we selected an NDVI ratio threshold of 0.2—0.4 for the SOS; and a drop in the NDVI ratio below 0.4—0.6 was interpreted to signify the EOS. An advantage to the NDVI ratio method is that a change, within a certain range, in the NDVI ratio threshold, has little infuence on vegetative phenological season dates (White, Thornton, and Running 1997). Generally, the NDVI ratio threshold value was lower in the east than in the west along each transect. Using the NDVI thresholds,the SOS, EOS, Peak-t, and LOS were estimated for each sampling pixel of the study area for each year on record. In sparsely vegetated areas, such as desert steppe, the SOS may not occur in years of low precipitation. If the SOS could not be detected before late August, the LOS was set as 0, and a randomly selected pseudo SOS (250 days) in autumn, was assigned to the specifc pixels at the particular year for further analysis (Yu et al. 2003).
3. Results
3.1. SOS and annual precipitation
The SOS correlated signifcantly with the mean annual precipitation along the transects (R2= 0.897, p < 0.0001 for the south transect; R2= 0.522, p < 0.0005 for the north transect,Figure 2(a)). In other words, drier sites started their growing season later than wetter ones. The SOS date ranged widely from early May at wetter sites (meadow steppe) to early July at drier sites (desert steppe). The ftting functions were different between the two transects. The SOS date was spatially advanced by 24.9 days for the south transect and 10.4 days for the north transect, with an increase of 100 mm in annual precipitation. The diferent response of the SOS to precipitation might be explained by a diference in annual mean temperature, which caused a diference in the amount of evapotranspiration between the two transects with similar annual precipitation. Due to the relatively higher annual temperature and a severe lack of water in the winter and spring,precipitation would strongly determine the dates of the SOS along the south transect.
3.2. Peak-t and annual precipitation
There was negative correlation between the Peak-t and mean annual precipitation along annual precipitation gradients in both transects (R2= 0.405, p < 0.0001 for the south transect; R2= 0.562, p < 0.0001 for the north transect, Figure 2(b)). The Peak-t along the south transect advanced by 4.2 days per increase of 100 mm of precipitation, while the Peak-t along the north transect advanced by 5.7 days per increase of 100 mm of precipitation. ThePeak-t demonstrated clear trends along both precipitation gradients along both transects.
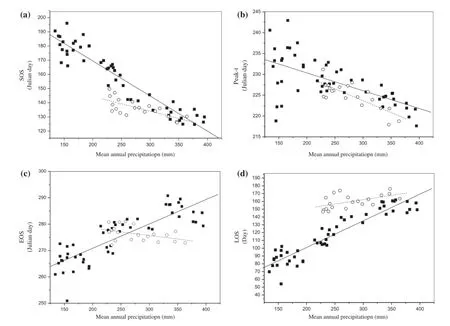
Figure 2.Relationship between the (a) SOS, (b) Peak-t, (c) EOS, (d) LOS9 and mean annual precipitation (solid squares denote the south transect and open circles denote the north transect).
3.3. EOS and annual precipitation
Along the south transect, there was positive correlation between the date of the EOS and mean annual precipitation (R2= 0.709, p < 0.0001, Figure 2(c)). The EOS occurred in mid-October at wetter sites (meadow steppe),in late September at drier sites (desert steppe), and in early October in typical steppe (average wetness) that received moderate precipitation between wetter and drier sites. Along the south transect, the EOS occurred 9.2 days later with every 100 mm increase in annual precipitation. Along the north transect there was negative correlation between the date of the EOS and mean annual precipitation (R2= 0.179, p < 0.1, Figure 2(c)). Due to the higher latitude and cooler temperatures of the north transect, precipitation in late autumn could lead to lower soil temperatures during the late vegetation growing season (Piao et al. 2006), which may explain the earlier date of the EOS accompanied by an increase in precipitation(Figure 2(c)).
3.4. LOS and annual precipitation
There was positive correlation between the LOS and mean annual precipitation along both transects (R2= 0.876,p < 0.0001 for the south transect; R2= 0.290, p < 0.05 for north transect, Figure 2(d)). Along precipitation gradients, the LOS increased from 76 days at the driest end(desert steppe) to 162 days at the wettest end (meadow steppe). The average LOS in a typical steppe with moderate hydrothermal (water and heat) conditions was 120 days(Figure 2(d)). For every 100 mm increase in precipitation,the LOS extended by 34.1 days in the south transect and by 7.7 days in the north transect. There are two reasons that may explain why there were diferences in the response of LOS to annual precipitation between the two transects. First, annual precipitation infuenced the SOS along the south transect more than along the north transect. The negative correlation between the date of SOS andthe mean annual precipitation implies increased precipitation may have contributed to the advance in the date of the SOS along the south transect. Secondly, the EOS was strongly controlled by precipitation along the south transect (R2= 0.709, p < 0.0001), so increases in the mean annual precipitation may have delayed the EOS. In the north transect, however, there was negative correlation(R2= 0.424, p < 0.1) between the EOS and mean annual precipitation, indicating that precipitation could cause an earlier EOS date.
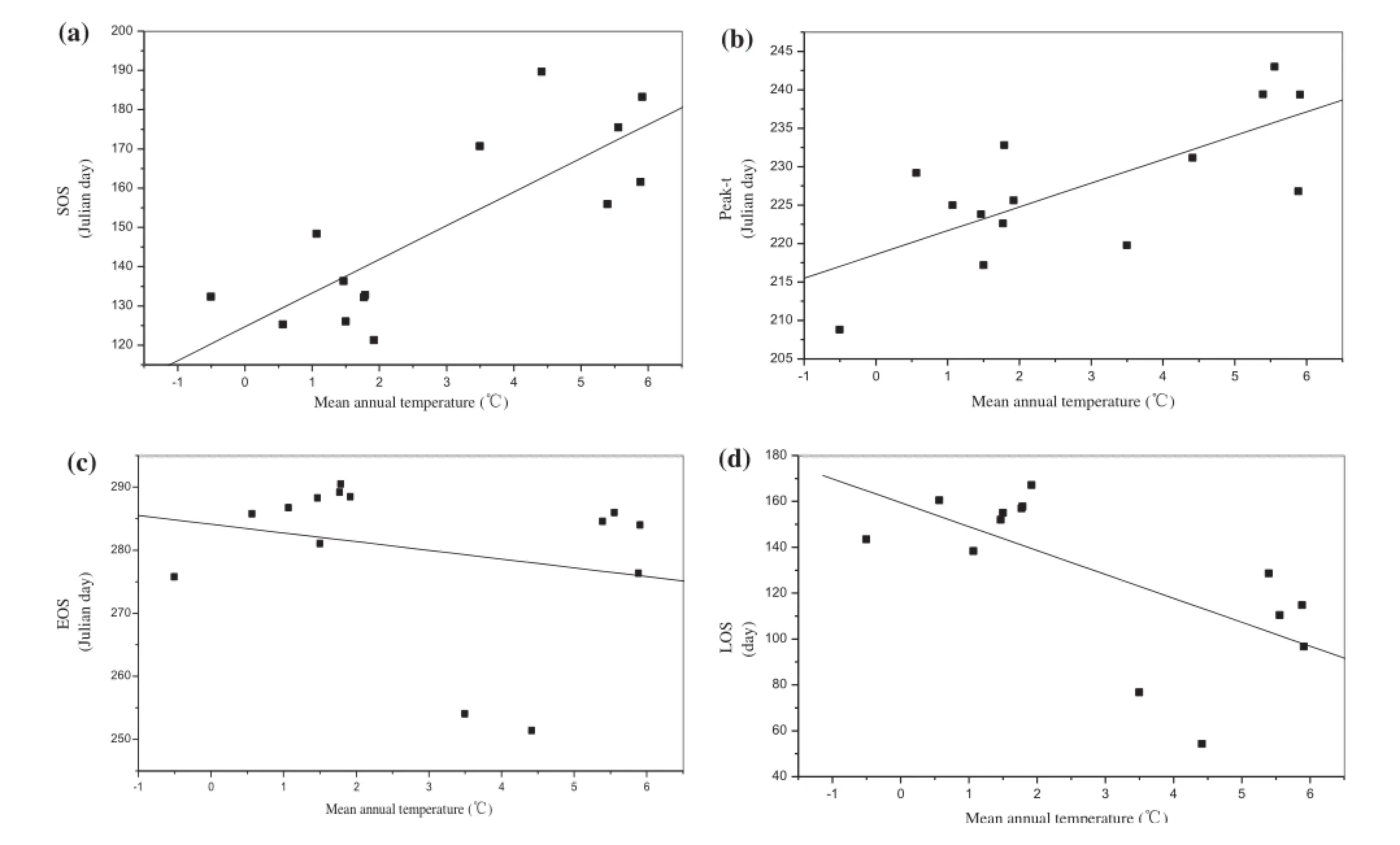
Figure 3.Relationship between the (a) SOS, (b) Peak-t, (c) EOS, (d) LOS and mean annual temperature.
3.5. Phonological parameters and mean annual temperature
There was positive correlation between the SOS, Peak-t,and mean annual temperature in the IMS (R2= 0.647,p < 0.001 for SOS, Figure 3(a); R2= 0.522, p < 0.005 for Peak-t, Figure 3(b)). In other words, warmer sites started their growing season later than cooler ones. The SOS and Peak-t occurred 8.6 and 3.1 days later with every 1 °C increase in mean annual temperature, respectively. In the IMS, however, there was negative correlation between the EOS, LOS, and mean annual temperature (R2= 0.543,p < 0.5 for EOS, Figure 3(c); R2= 0.435, p < 0.05 for LOS,Figure 3(d)). This meant that warmer sites ended their growing season earlier than cooler ones, and therefore had shorter LOS.
4. Discussion
The results supported our hypothesis that there would be negative correlation between the SOS date and annual precipitation. In other words, an increase in annual precipitation causes the advancement of the SOS date for the IMS. In the Patagonian steppe, however, the SOS date was found to be independent of the mean annual precipitation along precipitation gradients (Jobbagy, Sala,and Paruelo 2002). One possible cause of this diference is the efect of the temporal distribution of annual precipitation on vegetation activity. In the IMS, most of the annual precipitation falls during the summer, and severe water defciencies frequently occur in the spring and early summer. During these months, precipitation becomes a major control over vegetation growth, infuencing the SOS date by afecting water availability in the region. For the Patagonian steppe, more than 70% of the annual precipitation occurs during the fall and winter. As a result, during the spring, the soil has enough water to support vegetation growth. Thus,it is very likely that temperature, rather than precipitation,controls spring vegetation growth in the Patagonian steppe.
The typical steppes along both transects in the present study had similar mean annual precipitation values but diferent mean annual temperatures (Figure 1). The north transect, located in a region of higher latitude, had a lower mean annual temperature, and thus had lower rates of evapotranspiration and more available water than the south transect. The SOS date for typical steppes occurred earlier along the north transect than along the south transect (Figure 2(a)), indicating that temperature plays a minor (or even negative) role in regulating plant growth in steppe ecosystems that receive less than 400 mm of annual precipitation (Epstein, Lauenroth, and Burke 1997; Figure 3(a)). If temperature has a determining (positive) role in steppe ecosystem SOS dates, the SOS date along the south transect would have occurred earlier than along the north transect, due to its location at a lower latitude with higher temperatures. Based on temporal analysis, Lee et al. (2002)suggested that the SOS date for a year of high precipitation and low temperatures in the central IMS occurred earlier in the year than it did during a year of low precipitation and high temperatures. Thus, for the IMS, it is precipitation rather than temperature that determines the timing of the SOS date. Based on spatial analysis, Shen et al. (2014) found a comparable pattern. The areas with a greater increase in pre-season temperature did not necessarily experience a stronger advance of green-up date. In contrast, the areas with increasing pre-season precipitation overlapped with the areas having an advancing SOS, while the areas with declining pre-season precipitation experienced a delayed SOS in the Qinghai—Tibetan Plateau.
There was positive correlation between the EOS date and mean annual precipitation along the south transect. This supported our hypothesis that the EOS date would be delayed as precipitation increases for this semi-arid biome. The occurrence of a delay in the EOS date due to an increase in precipitation did not occur along the north transect. Instead, the EOS date occurred earlier with increasing precipitation. The phenological patterns of the north transect concurred with the results of Gu et al.(2012), who analyzed phenological ground observations and climate data in the IMS and concluded that an increase in precipitation during autumn could advance the EOS date for meadows (located between steppe and forest). Due to the higher latitude and cooler environment along the north transect, temperature is the limiting factor for vegetation growth (Piao et al. 2006). An increase in precipitation (usually accompanied by an increase in cloud cover and a reduction in incoming solar radiation) can signifcantly reduce soil temperature, preventing the root system from absorbing water from near-frozen soil during the late vegetation growing season (Piao et al. 2006). Furthermore,low temperatures inhibit plant photosynthesis by reducing photosynthesis-related enzyme activity, causing structural damage to chloroplasts (organelles necessary for photosynthesis) and to the bio-membrane, resulting in metabolic disequilibrium of reactive oxygen in plants.
Disclosure statement
No potential confict of interest was reported by the authors.
Funding
This study was jointly supported by the National Basic Research Program of China [973 Program, 2012CB956202] and the Collaborative Innovation Center of Research and Development on Tibetan Characteristic Agricultural and Animal Husbandry Resources.
Notes on contributors
Zhou Yao-Zhi is an associate professor at the Agricultural and Animal Husbandry College of Tibet University. His main research interests are climate change impact and adaptation, ecosystem dynamic, and grassland ecology. His recent publications include papers in Environmental Science, Arid Land Geography, and other journals.
Jia Gen-Suo is a professor at IAP, CAS. His main research interests are ecosystem dynamic, vegetation process—atmosphere environment interaction, land use and land cover change,climate change impact and adaptation. His recent publications include papers in Journal of Geophysical Research-Atmosphere, Climatic Change, Theoretical and Applied Climatology, Advances in Atmospheric Sciences, Remote Sensing of Environment, and other journals.
References
de Beurs, K. M., and G. M. Henebry. 2005. “Land Surface Phenology and Temperature Variation in the International Geosphere-Biosphere Program High-Latitude Transects.”Global Change Biology 11 (5): 779—790.
Brown, M. E., and K. M. de Beurs. 2010. “The Response of African Land Surface Phenology to Large Scale Climate Oscillations.”Remote Sensing of Environment 114 (10): 2286—2296.
Burke, I. C. 2000. “Landscape and Regional Biogeochemistry: Approaches.” In Methods in Ecosystem Science, edited by O. E. Sala, R. B. Jackson, H. A. Mooney, and R. W. Howarth, 277—288. New York: Springer Verlag.
Chen, X., and J. Li. 2009. “Relationship between Leymus chinensis Phenology and Meteorological Factors in Inner Mongolia Steppes.” Acta Ecologica Sinica 29 (10): 5280—5290.
Cleland, E. E., N. R. Chiariello, S. R. Loarie, H. A. Mooney, and C. B. Field. 2006. “Diverse Responses of Phenology to Global Changes in a Grassland Ecosystem.” Proceedings of the National Academy of Sciences 103 (37): 13740—13744.
Cong, N., S. Piao, A. Chen, X. Wang, X. Lin, S. Chen, S. Han, G. Zhou,and X. Zhang. 2012. “Spring Vegetation Green-up Date in China Inferred from SPOT NDVI Data: A Multiple Model Analysis.”Agricultural and Forest Meteorology 165 (15): 104—113.
Epstein, H. E., W. K. Lauenroth, and I. C. Burke. 1997. “Efects of Temperature and Soil Texture on ANPP in the US Great Plains.”Ecology 78 (8): 2628—2631.
Finzi, A. F., A. T. Austin, E. E. Cleland, S. D. Frey, B. Z. Houlton, and M. Wallenstein. 2011. “Responses and Feedbacks of Coupled Biogeochemical Cycles to Climate Change: Examples from Terrestrial Ecosystems.” Frontiers in Ecology and the Environment 9 (1): 61—67.
Gu, R. Y., W. C. Zhou, M. L. Bai, X. C. Li, R. Q. Di, and J. Yang. 2012.“Impacts of Climate Change on Phenological Phase of Herb in the Main Steppe in Inner Mongolia.” Acta Ecological Sinica 32 (3): 767—776.
Jia, G. J., H. E. Epstein, and D. A. Walker. 2003. “Greening of Arctic Alaska, 1981—2001.” Geophysical Research Letters 30 (20),2067: 1—4. doi:10.1029/2003GL018268.
Jobbagy, E. G., O. E. Sala, and J. M. Paruelo. 2002. “Patterns and Controls of Primary Production in the Patagonian Steppe: A Remote Sensing Approach.” Ecology 83 (2): 307—319.
Knapp, A. K., and M. D. Smith. 2001. “Variation among Biomes in Temporal Dynamics of Aboveground Primary Production.”Science 291 (5503): 481—484.
Lee, R., F. Yu, K. P. Price, J. Ellis, and P. Shi. 2002. “Evaluating Vegetation Phenological Patterns in Inner Mongolia Using NDVI Time-Series Analysis.” International Journal of Remote Sensing 23 (12): 2505—2512.
Morisette, J. T., A. D. Richardson, A. K. Knapp, J. I. Fisher, E. A. Graham, J. Abatzoglou, B. E. Wilson, et al. 2009. “Tracking the Rhythm of the Seasons in the Face of Global Change: Phenological Research in the 21st Century.” Frontiers in Ecology and the Environment 7 (5): 253—260.
Piao, S. L., J. Y. Fang, L. M. Zhou, P. Ciais, and B. Zhu. 2006. “Variations in Satellite-Derived Phenology in China's Temperate Vegetation.”Global Change Biology 12 (4): 672—685.
Richardson, A. D., R. S. Anderson, M. A. Arain, A. G. Barr, G. Bohrer,G. S. Chen, J. M. Chen, et al. 2012. “Terrestrial Biosphere Models Need Better Representation of Vegetation Phenology: Results from the North American Carbon Program Site Synthesis.”Global Change Biology 18 (2): 566—584.
Shen, M. G., G. X. Zhang, N. Cong, S. P. Wang, W. D. Kong, and S. L. Piao. 2014. “Increasing Altitudinal Gradient of Spring Vegetation Phenology during the Last Decade on the Qinghai-Tibetan Plateau.” Agricultural and Forest Meteorology 189—190: 71—80. doi:10.1016/j.agrformet.2014.01.003.
Soudani, K., G. Hmimina, N. Delpierre, J. Y. Pontailler, M. Aubinet,D. Bonal, B. Caquet, et al. 2012. “Ground-Based Network of NDVI Measurements for Tracking Temporal Dynamics of Canopy Structure and Vegetation Phenology in Diferent Biomes.” Remote Sensing of Environment 123: 234—245. doi:10.1016/j.rse.2012.03.012.
Tucker, C. J., D. A. Slayback, J. E. Pinzon, S. O. Los, R. B. Myneni,and M. G. Taylor. 2001. “Higher Northern Latitude NDVI and Growing Season Trends from 1982 to 1999.” International Journal of Biometeorology 45 (4): 84—190.
White, M. A., K. M. de Beurs, K. Didan, D. W. Inouye, A. D. Richardson, O. P. Jensen, J. O'Keefe, et al. 2009.“Intercomparison, Interpretation, and Assessment of Spring Phenology in North America Estimated from Remote Sensing for 1982—2006.” Global Change Biology 15 (10): 2335—2359. doi:10.1111/j.1365-2486.2009.01910.x.
White, M. A., P. E. Thornton, and S. W. Running. 1997. “A Continental Phenology Model for Monitoring Vegetation Responses to Interannual Climatic Variability.” Global Biogeochemical Cycles 11 (2): 217—234.
Yu, H. Y., E. Luedeling, and J. C. Xu. 2010. “Winter and Spring Warming Result in Delayed Spring Phenology on the Tibetan Plateau.” Proceedings of the National Academy of Sciences 107(51): 22151—22156.
Yu, F. F., K. P. Price, J. Ellis, and P. Shi. 2003. “Response of Seasonal Vegetation Development to Climatic Variations in Eastern Central Asia.” Remote Sensing of Environment 87 (1): 42—54.
物候; 降水量; 草原; 遥感
2 October 2015
CONTACT JIA Gen-Suo jiong@tea.ac.cn
© 2016 The Author(s). Published by Taylor & Francis.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
猜你喜欢
草原歌声(2018年2期)2018-12-03 08:14:06
草原歌声(2018年2期)2018-12-03 08:13:56
中成药(2018年6期)2018-07-11 03:01:34
中成药(2018年1期)2018-02-02 07:20:05
中国公路(2017年16期)2017-10-14 01:04:48
中成药(2017年6期)2017-06-13 07:30:35
中央民族大学学报(自然科学版)(2017年1期)2017-06-11 07:13:18
中成药(2017年4期)2017-05-17 06:09:36
文学港(2016年7期)2016-07-06 15:22:08
小学教学研究(2016年36期)2016-05-17 05:52:05
 Atmospheric and Oceanic Science Letters2016年3期
Atmospheric and Oceanic Science Letters2016年3期
- Atmospheric and Oceanic Science Letters的其它文章
- Discrepancies in boreal summer monsoon rainfall between GPCP and CMAP products during 1979-2014
- Analysis of ‘9.4’ unusual rainfall in Beijing during autumn 2015
- Characteristics of pressure gradient force errors in a terrain-following coordinate
- Improved simulation of the East Asian winter monsoon interannual variation by IAP/LASG AGCMs
- The relationship between the Arctic Oscillation and ENSO as simulated by CCSM4
- Estimation of the surface heat budget over the South China Sea