
米老排林分生长规律及生物量分布格局研究
2016-11-17 01:32徐洪芳刘子斐陈珍洪李铁华
湖南林业科技 2016年6期
徐洪芳, 刘子斐, 陈珍洪, 李铁华
(1. 广东省云浮市国有同乐林场, 广东 郁南 527199; 2.中南林业科技大学 林学院, 湖南 长沙 410004)
米老排林分生长规律及生物量分布格局研究
徐洪芳1, 刘子斐2, 陈珍洪1, 李铁华2
(1. 广东省云浮市国有同乐林场, 广东 郁南 527199; 2.中南林业科技大学 林学院, 湖南 长沙 410004)
通过选定同乐林场米老排林分作为研究对象,采用样地调查与解析木对该树种进行生长特性研究。结果表明: 0~6年是米老排幼林生长的阶段,树高、胸径生长均较为旺盛,6~12年材积与胸径生长迅速,而树高生长速度呈现下降趋势, 12~16年是米老排生长树高、胸径、材积这三个指标的速生时期,28年以后树高生长速度逐渐下降,材积与胸径生长还呈现上升趋势。米老排的单株和林分生物量排序为干>枝>根>皮>叶。
米老排; 生长规律; 生物量; 生物量结构
米老排(Mytilarialaosensis)又名壳菜果、三角枫,为金缕梅科常绿乔木树种,天然分布于我国广东、广西和云南省区以及越南和老挝等地,具有生长快、成材早、适应性较广、抗性强、造林简易、成活率高、萌芽力强、落叶层厚、林地保湿效果好等优点,在提高土壤肥力、涵养水源、防火等方面具有较好的效能。米老排干形通直,材质较好,色泽美观,经久耐用,适于制作家具、胶合板、客车车箱、纸浆等,同时也是水土保持、土壤改良、混交造林以及生物防火的优良树种[1]。米老排叶子肥大嫩绿,营养成分含量高,是一种饲料资源;所分泌树脂的化学成分主要是脂类物质,其种仁含油量高达36.8%,也是重要的油脂资源[2]。
对于米老排,前人做了一些研究,主要集中在米老排防火特性[3]、米老排土壤与水源涵养特性[4-6]、米老排木材性质[7-9]、群落物种多样性[10-11],对米老排林分生长规律及生物量分布格局研究较少。笔者通过研究,探讨米老排林分生长规律及生物量分布格局特征,为提高林分的经营管理水平收益提供科学依据。
1 研究区概况
同乐林场位于广东省云浮市郁南县东北部,总面积 7453hm2(含同乐大山保护区),距郁南县城约20 km,距广西梧州约30 km,跨越两广交界。林场属低山丘陵地带,以低山为主,海拔多在100~691 m之间,地貌组成的岩性以变质岩和砂页岩为主,位于北回归线以南,属南亚热带季风气候,年均气温 21.4 ℃,年均降水量为 1433 mm。林场境内森林茂密,但由于人为活动的影响,现有原生森林植被已经很少,现在的次生植被大部分是人工杉木和马尾松、湿地松。
2 研究方法
2.1样地设置与调查
同乐林场米老排林分为人工纯林,在林分内设置面积为25.82 m×25.82 m的标准地3个,在样地内进行每木检尺,测定胸径、树高、材积等。根据每木检尺结果,选取平均标准木做解析木,伐倒并记录解析木所处的立地条件、林分状况、冠幅和相邻树木之间的关系等生境因子,通过解析木分析米老排的生长过程和样木的生物量格局情况。在标准地内四个角和中间各设置一个1 m×1 m的草本样方,进行草本、灌木的记录采样。
2.2林分生物量测定
米老排生物量的测定采用收获法测定样木的生物量,分别实测干、枝、叶、根等的鲜重。对标准木采用分层切割法进行取样,分别采集各器官部分样品皮、枝、叶各500 g,根1000 g,干1200 g,带回室内至于恒温(其中叶的烘干温度设为80 ℃,其余各部分为103 ℃)条件下烘干,叶烘干时间为24 h,其他各组分烘至恒重。在实验室获得各组分样品烘干重量(即生物量)后,通过比例推断出各组分的全株生物量:
单木各器官生物量=
各器官鲜重×(1-含水率);
对于灌木和草本,全部收获称重并取样带回实验室烘干,然后换算成林分单位面积生物量。
3 结果与分析
3.1米老排生长规律
3.1.1 样木高生长规律 从图1和图2可以看出,样木米老排树高生长过程中,连年生长波动较大,在0~6年时,连年生长量随年龄的增加呈现较大的增长趋势,8~12年,高生长相对缓慢,随后出现了快速生长。分别在6年和14年时,连年生长量达到高峰,连年生长量达到了1 m/年,表明米老排林分一个是在幼林生长的阶段,树高生长较快,另一个是在林分郁闭后林木为了争夺上方光照而迅速进行高的生长,从第14年开始,高生长量下降迅速,从第14年的1 m/年下降到了第16年的0.5 m/年,第16年后,树高生长比较缓慢,变化也不明显,维持在0.35 m/年左右,说明米老排的树高前期生长快,主要集中在前面16年(12.6 m),占总生长量(20.45 m)的61.6%以上。树高平均生长量在第6年达到最大值后,逐渐下降,曲线比较平缓。

图1 树高生长曲线(35年)Fig.1 Growth curve of tree height (35 a)

图2 树高生长量随年龄的变化(35年)Fig.2 Change of tree height increment with tree age (35 a)
3.1.2 样木胸径生长规律 从图3和图4可以看出,米老排样木林分胸径的连年生长量和平均生长量都随着龄阶的增长而增大,连年生长量在第4年开始缓慢上升,在第10年达到最高值0.97 cm,然后缓慢下降,在第16年开始下降比较迅速,之后到第20年又开始缓慢上升,一直持续到第34年。整体来看,16年前,连年生长量都在0.8 cm以上;平均生长量随年龄的增加而上升,在第12年时达到最高值0.76 cm,然后缓慢下降,整体过程变化幅度不明显,在10~16年之间生长最快,都在0.72 cm以上。连年生长量与平均生长量相交于16~18年之间,此期间是胸径生长旺盛的时期,随后胸径的生长量逐渐减缓,20年之后可能由于抚育间伐营林措施等原因,使得连年生长逐渐增大,在28~30年之间又出现平均生长和连年生长再次相交,连年生长量继续上升。

图3 胸径生长曲线(35年)Fig.3 Growth curve of DBH (35 a)

图4 胸径生长量随年龄的变化(35 a)Fig.4 Change of DBH increment with tree age(35 a)
3.1.3 样木材积生长规律 从图5和图6可以看出,随着树龄的增长,米老排材积的连年生长量基本呈现增长的态势,表明米老排的生长越来越快,30年后是材积生长的速生期,第35年,材积的生长量达到最大值(0.028 2 m3/年),样地内米老排没有出现连年生长率明显降低的趋势,平均生长量也是随着年龄的增加而上升,且连年生长量曲线没有与平均生长量曲线相交,说明林木还未达到数量成熟,米老排的数量成熟年龄应在35年后。

图5 材积生长曲线(35年)Fig.5 Growth curve of wood quantity (35 a)

图6 材积生长量随年龄的变化(35年)Fig.6 Change of wood quantity increment with tree age(35 a)
3.2单木及林分生物量
米老排单株和林分的各器官生物量如表1。

表1 米老排各器官组分生物量比较表(35年)Tab 1 Analyticalwoodcomparisonofbiomassineachorgan(35a)器官生物量(kg)各组分生物量百分比(%)林分生物量(t/hm2)树干495.7973.49268.31树枝81.7112.1144.19叶8.451.254.57皮38.125.6520.62根50.577.527.35总生物量674.65100.00364.87
从表1可以看出, 35年生米老排林分生物量干、枝、皮、叶、根所占比例分别为73.49%、12.11%、1.25%、5.65%、7.5%。各组分生物量均存在干>枝>根>皮>叶的规律,这种生物量分配规律基本符合乔木树种生物学特性。标准木的树干生物量皆超过总生物量的三分之二,远大于其他各组分,由此可以得出,米老排是具有较大的木材收益的珍贵树种。
3.3生物量结构特征
所谓生物量结构特征是指林分各组分质量之间或各组分质量与总生物量的比值[12]。包括:
(1) 枝叶比(BNR)=Wb/Wl
(2) 枝叶指数(BNI)=(Wb+Wl)/Wt
(3) 光合器官与非光合器官比值(FC)=
Wl/Wt
(4) 干材与地上部分生物量比值(STR)=
Ws/Wt
由表2可知,光合器官与非光合器官比值(FC)为0.01,FC值较低 ,表明林分光合作用较弱,不能充分利用光照进行光合作用;干材与地上部分生物量比值(STR)为0.73,说明树干这一组的生物量占林分生物量的比例较大;枝叶比(BNR)值为9.67。研究表明,当BNR值小而FC值高时,表明林分结构合理。因为FC值高,且BNR值小,表明林木叶面积大,可充分利用光照,通过光合作用为林木提供更加充足的养料,以满足其生长,进而有利于提高林分的总生物量,以取得更好的经济、生态、社会效益。而从表2可知,样地的枝叶比(BNR)值均较大,而FC值小,说明样地虽立地条件良好,但林分密度偏小,分布不合理,林分生产力较低,在调查时也发现林分内林木生长层次不齐,覆盖度较小,必须利用补植、补种以及补播等方法加以改造,以使林分结构协调互利,增大其生产潜力。

表2 生物量结构特征Tab 2 Biomassstructurecharacteristics35年BNRBNIFCSTR9 670 130 010 73
3.4林分生产力
通常用“年平均净生产量”作为生产力的估测指标,即各组分的生物量与林龄之比值,用下式表示:
ΔW=(Wa-Wa-n)/n(t/hm2.a)
当a=33时,n取33,当a=35时,n取35,当a=38时,n取38,米老排林分净生长量的计算结果如表3:
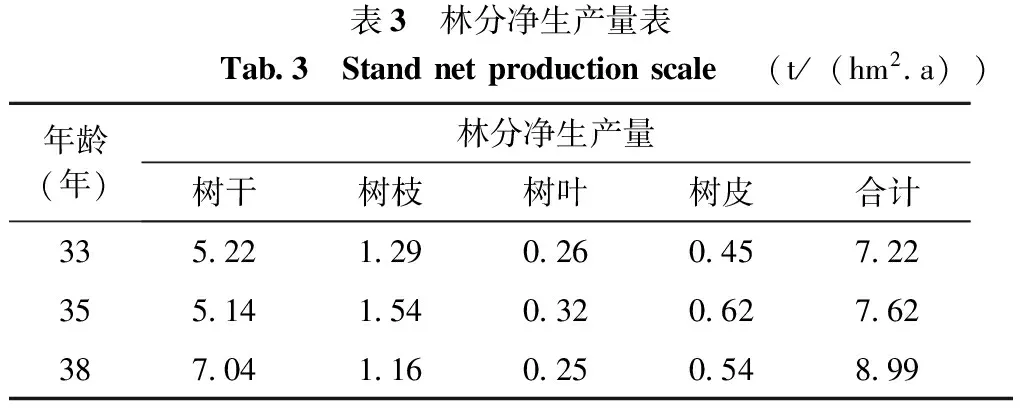
表3 林分净生产量表Tab 3 Standnetproductionscale (t/(hm2 a))年龄(年)林分净生产量树干树枝树叶树皮合计335 221 290 260 457 22355 141 540 320 627 62387 041 160 250 548 99
从表3可以看出,不同年龄阶段米老排林分净生产量有一定差异,随着林龄逐步增大,米老排的平均生产力也随之增大,38年生大于35年生大于33年年生林分。其中树干的净生产力最大,分别为5.22,5.14 t/(hm2.a)以及7.04 t/(hm2.a)。树叶的净生产力最小,分别为0.26,0.32,0.25(t/hm2.a),树叶的净生产力较低。
4 结论与讨论
(1) 米老排的树高、胸径和材积总生长量都是随着年龄的增大而增大。8~16年是胸径生长的旺盛时期,连年生长与平均生长均在这段时间达到最大值,并相交于16年左右,随后在30年左右可能由于抚育等措施连年生长再次上升与平均生长相交。0~6年是幼林生长的阶段,树高生长较快,12~14年是树高生长的旺盛时期,28年以后树高增速逐渐下降。材积的连年生长量和平均生长量在35年之前两曲线未相交,说明该米老排林分在35年之前未达到数量成熟龄。据郭文福等[13]在广西大青山对米老排人工林生长规律的研究,米老排林分树高和胸径连年生长与平均生长在第 3~4 年最快,在第 15~17年林分进入材积的数量成熟期,树高生长规律与本研究基本相符,胸径与材积与本研究存在差异,本研究分别延后5~10年和10~15年,造成这一差异的原因一方面可能是两地的气候条件不一样,导致米老排生长过程不同,另一方面可能是由于本研究的样地长期疏于管理,而且本研究的解析木年龄较长,造成生长情况出现了偏差。
(2) 米老排标准木,各器官生物量由高到低顺序:干>枝>根>皮>叶,叶生物量占林分总生物量较少,样地林木不能充分利用光照进行光合作用,可以采取相应营林措施提高FC值与BNI值,保证BNR值小,使林木叶面积增大,即可充分利用光照,通过光合作用为林木提供更加充足的养料,以满足其生长,进而有利于提高林分的总生物量,以取得更好的经济、生态、社会效益,增大其生产潜力。据蔡学林等[14]对杉木标准木各器官生物量分布研究,杉木各器官生物量排序为干>根>枝>叶>皮,树干生物量均占据优势,这表明米老排与杉木一样是优良的用材树种,本研究树根与树叶生物量较小的原因主要可能是由于树种不同造成各器官生物量的不同。
(3) 米老排各器官生产力由高到低顺序:干>枝>皮>叶,树叶的生产力较低。据丁贵杰等[15]对马尾松人工林各器官生产力的研究,马尾松各器官生产力排序为干>叶>枝>皮,除本研究树叶生产力较低意外,其他各器官基本相同,造成差异的主要原因可能是由于本研究树叶收集范围较小所造成的。
[1] 陈志云.杉木-米老排混交林生态系统生产力及养分特征研究[D].福州:福建农林大学,2010.
[2] 洪永辉,林能庆,廖柏林,等.米老排优树选择技术研究[J].林业勘察设计(福建),2015(1):1-5.
[3] 吕福如.防火生土带改建米老排防火林带的林木生长与地力维持效果[J]. 河北林果研究,2002,17(4):317-322.
[4] 王光玉.杉木混交林水源涵养和土壤性质研究[J].林业科学,2003,39(S1):15-20.
[5] 姚庆端.不同杉木混交模式土壤肥力及土壤蓄水量研究[J]. 福建林学院学报,1996,16(3):282-286.
[6] 郭友林.米老排杉木混交林的水源涵养功能研究[J].林业科技开发,2002,16(s):30-31.
[7] 陈存及,陈伙法.阔叶树种栽培[M].北京:中国林业出版社,2000.
[8] 梁善庆,罗建举.人工林米老排木材化学成分及其在树干高度上的变异[J].中南林学院学报,2004,24(5):28-31.
[9] 梁善庆,罗建举.人工林米老排木材的物理力学性质[J].中南林业科技大学学报:自然科学版,2007,27(5):97-100,116.
[10] 龚辉.杉木米老排人工群落乔木层结构与动态研究[J].江西农业大学学报,2000,22(1):46-49.
[11] 王小燕,薛杨,宿少锋,等.文昌沿海地区林下套种米老排的适应性分析[J].热带林业,2016,44(1):4-5,11.
[12] 严仁发,朱守谦.马尾松人工林生物量测定方法比较[J].贵州农学院丛刊,1984(4): 101-105.
[13] 郭文福,蔡道雄,贾宏炎,等.米老排人工林生长规律的研究[J].林业科学研究,2006,19(5):585-589.
[14] 蔡学林,张志云,欧阳勋志,等.杉木人工林生物量的研究[J].江西农业大学学报,1997,19(6):138-145.
[15] 丁贵杰,王鹏程.马尾松人工林生物量及生产力变化规律研究II.不同林龄生物量及生产力[J].林业科学研究,2002,15 (1):54-60.
StudyongrowthlawsandbiomassdistributionpatternofMytilarialaosensis
XU Hongfang1, LIU Zifei2, CHEN Zhenhong1, LI Tiehua2
(1.Tongle State-owned Forest Farm of Yunfu City in Guangdong Province,Yunan 527199,China;2.Central South University of Forestry and Technology, Changsha 41004, China)
Through survey to the growth regularity and biomass structure ofMytilarialaosensisforest stand in Tongle State-owned Forest Farm of Guangdong Province, and stem analysis of sample trees to turns out that the general growth pattern ofM.laosensisis as follow: the height, DBH and volume ofM.laosensisall increase with age: 0~6a is growth stage of young stand and the increase of height and DBH is comparatively fast; in 6~12a, the volume and DBH increase rapidly but tree height growth shows a downward trend; 12~16a is the surging growth period of the three indexes ofM.laosensis:tree height, DBH and volume; After 28a, growth speed of tree height gradually decreases but the growth of volume and DBH still presents a rising tendency. The biomass of each single tree andM.laosensisstand are sorted into: trunk >branch > root > bark> leaf.
Mytilarialaosensis; growth pattern; biomass; biomass structure
2016-09-14
徐洪芳(1964-),广东省郁南市人,大专,林业工程师,主要从事营造杉木丰产林技术的研究。
Q 949.751.4
A
1003 — 5710(2016)06 — 0051 — 05
10. 3969/j. issn. 1003 — 5710. 2016. 06. 011
(文字编校:杨 骏)
猜你喜欢
广西林业科学(2022年3期)2022-07-08
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
江苏农业科学(2018年14期)2018-08-08
现代园艺(2018年1期)2018-03-15
亚热带资源与环境学报(2015年3期)2015-01-22
