
南方型紫花苜蓿叶片盐胁迫应答转录因子鉴定与分析
2016-10-31 04:35裴翠明张振亚
浙江农业学报 2016年4期
裴翠明,张振亚,马 进
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
南方型紫花苜蓿叶片盐胁迫应答转录因子鉴定与分析
裴翠明,张振亚,马进*
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
以南方型紫花苜蓿‘Millennium’为材料,以正常培养(WT_CK2)和250 mmol·L-1NaCl胁迫下72 h时(WT_N2)条件下的2个样品叶片进行转录组分析,鉴定紫花苜蓿叶片盐胁迫应答转录因子基因。同时随机挑选4个转录因子差异表达基因进行实时荧光定量qRT-PCR验证RNA-Seq结果的可靠性。紫花苜蓿叶片在250 mmol·L-1NaCl胁迫下72 h时,检测到表达量差异达到2倍以上的基因共7 497个,其中包括隶属于46个转录家族474个转录因子在盐胁迫下的差异表达,上调242个,下调232个。bHLH,NAC,WRKY和C2H2转录家族成员基因70%以上表达上调。实时荧光定量PCR分析表明4个随机选择的基因在胁迫前后的表达特点与表达谱测序结果一致。此外,还发现大量紫花苜蓿盐胁迫应答的转录因子候选基因如MsERF110,MsERF071,MsbHLH36,MsZFP,MsHSFB-3,MsMYB,MsNAC和MsWRKY等。同时,也揭示了紫花苜蓿叶片对盐胁迫响应可能是多种转录因子家族共同参与的应答过程。
南方型紫花苜蓿;转录因子;叶片转录组;盐胁迫
植物盐胁迫是一种综合胁迫,包括生理性干旱胁迫、渗透胁迫以及离子毒害胁迫,涉及到信号传递、多基因的协同控制、蛋白质翻译与表达等一系列复杂的分子调控网络。然而由于对植物耐盐机理的认识不够,且现有可用耐盐基因的种类较少,限制了植物耐盐育种的发展。基因对盐胁迫应答主要表现在自身遗传物质的基因控制,其中部分差异表达基因在耐盐应激调控过程中发挥重要作用。
紫花苜蓿(MedicagosativaL.)主要分布在温带地区,除了作为牧草外,也是一种很好的水土保持绿化植物。近年来,国外高秋眠级紫花苜蓿品种引进解决了南方的种植难题,但紫花苜蓿耐盐能力有限,限制了在南方盐碱地大面积种植,因此,发掘参与其盐逆境响应中耐盐功能基因,并分析其耐盐分子机制,有助于利用基因工程技术培育适合南方盐碱地种植的紫花苜蓿新品种。
目前已经鉴定出部分苜蓿耐盐基因,Miller等[1]克隆出紫花苜蓿MsPDH和MsP5CDH两个脯氨酸脱氢酶基因,该基因具有增加游离脯氨酸积累功能;Luo等[2]利用RT-PCR方法从紫花苜蓿中分离得到受胁迫诱导表达的解旋酶基因MH1,该基因可提高活性氧(ROS)的清除和提高植物渗透调节能力;Long等[3]从紫花苜蓿中盐胁迫诱导中筛选出富含甘氨酸蛋白基因(MsGRP),该基因具有抑制种子萌发和拟南芥(Arabidopsisthaliana)幼苗生长的功能;Ma等[4]从紫花苜蓿盐胁迫中成功克隆一个MsGME基因,该基因对抗坏血酸合成起着重要的作用。蒺藜苜蓿(Medicagotruncatula)作为豆科模式植物,与苜蓿亲缘关系很近,与四倍体苜蓿基因组具有很高的同源性。Zahaf等[5]对豆科模式植物蒺藜苜蓿盐胁迫下全基因组分析,为开展紫花苜蓿耐盐分子机制研究提供了重要信息,但紫花苜蓿为同源四倍体,异花授粉植物,其基因组非常复杂,仍然难以揭示紫花苜蓿耐盐的分子机理。
转录因子作为调控下游基因表达的重要元件,调节着各种生理活动的关键环节,一些调节逆境应答的转录因子基因是改良植物耐盐性的重要靶标基因,因此,对逆境应答转录因子的研究是现代植物分子生物学研究的热点。目前采用高通量技术从转录组水平上鉴定和分析紫花苜蓿转录因子盐胁迫应答研究还鲜见报道。本研究利用RNA-Seq技术对盐胁迫处理前后的南方型紫花苜蓿幼苗叶片进行转录组测序,鉴定叶片响应盐胁迫应答转录因子及表达特性,以期更好地理解南方型紫花苜蓿对盐胁迫响应的分子机制,为进一步鉴定和克隆其重要的耐盐基因,提高南方型紫花苜蓿耐盐性状奠定基础。
1 材料与方法
1.1材料及处理
选择耐盐性相对较强的高秋休眠级南方型紫花苜蓿‘Millennium’为材料[6],将种子先用75%酒精消毒10 min,然后用灭菌水清洗3次,灭菌水中浸泡2 h于20 ℃生长箱内发芽。7 d后,挑选生长一致小苗转移至塑料盆,盆内装满细沙和珍珠岩(体积比3∶1),每天浇霍格兰营养液。30 d后加入250 mmol·L-1NaCl至营养液中作为盐胁迫处理,选取胁迫处理0和72 h的紫花苜蓿地上部叶片组织混合于液氮中保存备用。
1.2RNA的提取及检测
利用TRNzol Reagent试剂盒分别提取处理与对照组幼叶片RNA,提取过程按试剂盒说明进行。所提取的RNA经电泳检测合格后送至深圳华大基因科技有限公司进一步确认质量后,将同一样本所提取的幼叶RNA进行高通量测序。
1.3Illumina 测序
将盐处理组与对照组总RNA由华大基因科技有限公司完成文库构建与测序,将构建好的文库进行其质量和产量检测。对质量检测合格后的文库采用IlluminaHiSeqTM2000进行测序。将经Base calling测序产生的原始图像转化为序列数据(Raw data),继而转换为Clean data。采用短reads比对软件SOAPaligner/SOAP2将Clean reads分别与参考基因组蒺藜苜蓿进行比对。
1.4差异基因的选择
测序后使用RPKM法计算基因表达量,根据基因表达量(RPKM值)计算其在处理与对照中的差异表达倍数。差异表达倍数大于2倍以上(含2倍)的基因认定为差异表达基因。
1.5荧光定量RT-PCR
以分别提取的处理组与对照组叶片RNA为材料,采用TaKaRa公司的PrimeScript RTreagent Kit With cDNA Eraser试剂盒反转录合成cDNA,作为实时荧光PCR反应的模板。根据测序的基因表达谱结果随机挑取选出4个盐胁迫转录因子基因,Primer Premier 5.0软件设计荧光定量PCR引物(表1)。利用Primer 5.0 软件设计qRT-PCR引物,以苜蓿GAPDH(GenBank登录号为MTR_4g131180)为内参,引物为5′-GTGGTGC-CAAGAAGGTTGTTAT-3′和5′-CTGGGAATGATGTTGAAGGAAG-3′。各处理均做3次重复计算基因的相对表达量。
表1验证DEG数据准确性的qRT-PCR引物
Table 1The primers to detect the accuracy of DEG data by qRT-PCR
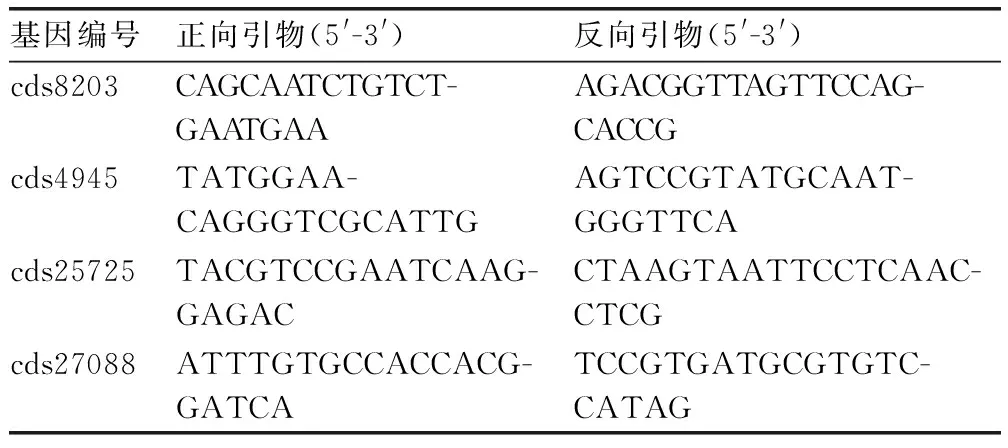
基因编号正向引物(5'-3')反向引物(5'-3')cds8203CAGCAATCTGTCT-GAATGAAAGACGGTTAGTTCCAG-CACCGcds4945TATGGAA-CAGGGTCGCATTGAGTCCGTATGCAAT-GGGTTCAcds25725TACGTCCGAATCAAG-GAGACCTAAGTAATTCCTCAAC-CTCGcds27088ATTTGTGCCACCACG-GATCATCCGTGATGCGTGTC-CATAG
2 结果与分析
2.1测序质量分析
对测得的reads分析表明:对照样本(WT_CK2)和盐胁迫处理样本(WT_N2) raw reads在去除接头序列、低质量的序列等后,得到高质量的reads。由于遗传背景的差异和注释信息量的限制,对照样本clean reads中近52.47%可被参考基因组注释,47.53%的clean reads未能注释到对应的基因(表2),盐胁迫处理样本(WT_N1)样本的reads注释情况相近。此外, 测序随机性分析表明:reads在基因中的分布均一,说明测序样品的mRNA完整性和片段化均一性很好,序列分析中没有mRNA降解和偏向性的影响。
表2DEG样品测序数据统计结果
Table 2Statistics of sequencing data of the DEG samples
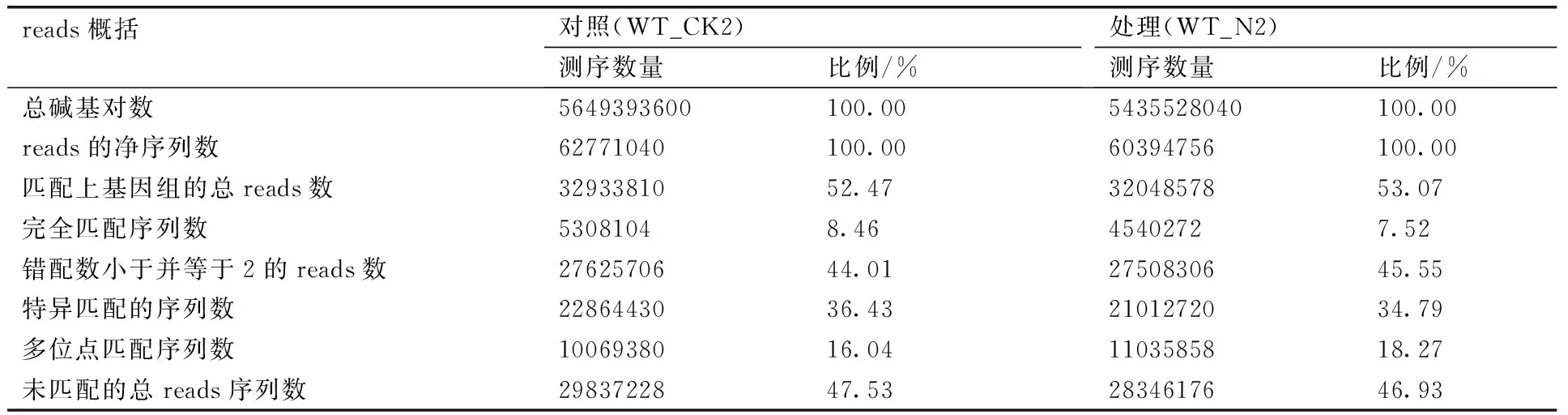
reads概括对照(WT_CK2)测序数量比例/%处理(WT_N2)测序数量比例/%总碱基对数5649393600100.005435528040100.00reads的净序列数62771040100.0060394756100.00匹配上基因组的总reads数3293381052.473204857853.07完全匹配序列数53081048.4645402727.52错配数小于并等于2的reads数2762570644.012750830645.55特异匹配的序列数2286443036.432101272034.79多位点匹配序列数1006938016.041103585818.27未匹配的总reads序列数2983722847.532834617646.93
2.2紫花苜蓿叶片盐胁迫应答转录因子基因分析
将盐处理组与对照组总RNA由华大基因科技有限公司完成文库构建与测序,以差异检验FDR值≤0.001且差异倍数不低于2倍为差异表达基因的选择标准,紫花苜蓿叶片在250 mmol·L-1NaCl胁迫下72 h时,共检测到31 372个基因表达量发生了改变,对照样本(WT_CK2)和盐胁迫处理样本(WT_N2)有7 497个基因表现为差异表达,其中4 078个DEGs在盐胁迫响应中表达上调,3 419个DEGs表达下调。南方型紫花苜蓿叶片盐响应下共有隶属于46个转录因子家族474个转录因子差异表达,占总差异表达基因的6.32%,其中242个上调、232个下调(图1)。
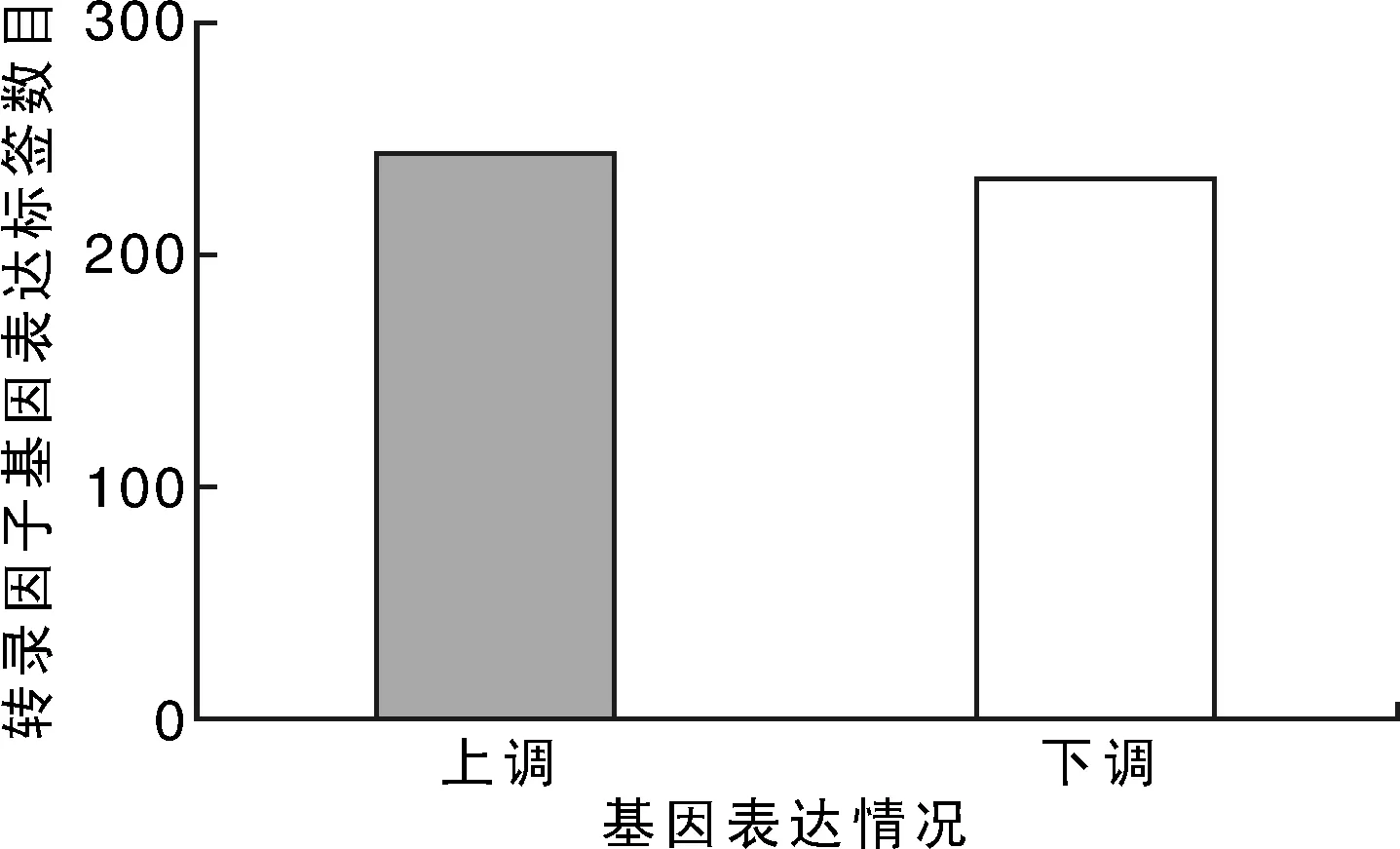
图1 正常和盐胁迫下紫花苜蓿叶片转录因子基因数目Fig.1 Distribution of differentially expressed genes and transcription factor number in the leaves of alfalfa under control and salt stress
图2可以看出:转录因子基因表达差异不一,Log2ratio基因表达量变化15倍以内,占差异表达转录因子基因总数的99.64%。其中,表达量上、下调1~2倍的基因分别占差异表达上、下调基因总数的42.56%和57.326%,表达量上、下调2~5倍的基因分别占37.60%和30.60%,表达量上、下调5~10倍的基因分别占5.37%和2.16%,表达量上、下调10~15倍以上的基因分别占14.05%和9.91%,表达量上调15倍以上的基因占0.41%。相比而言,上调基因的表达量增强幅度更大。
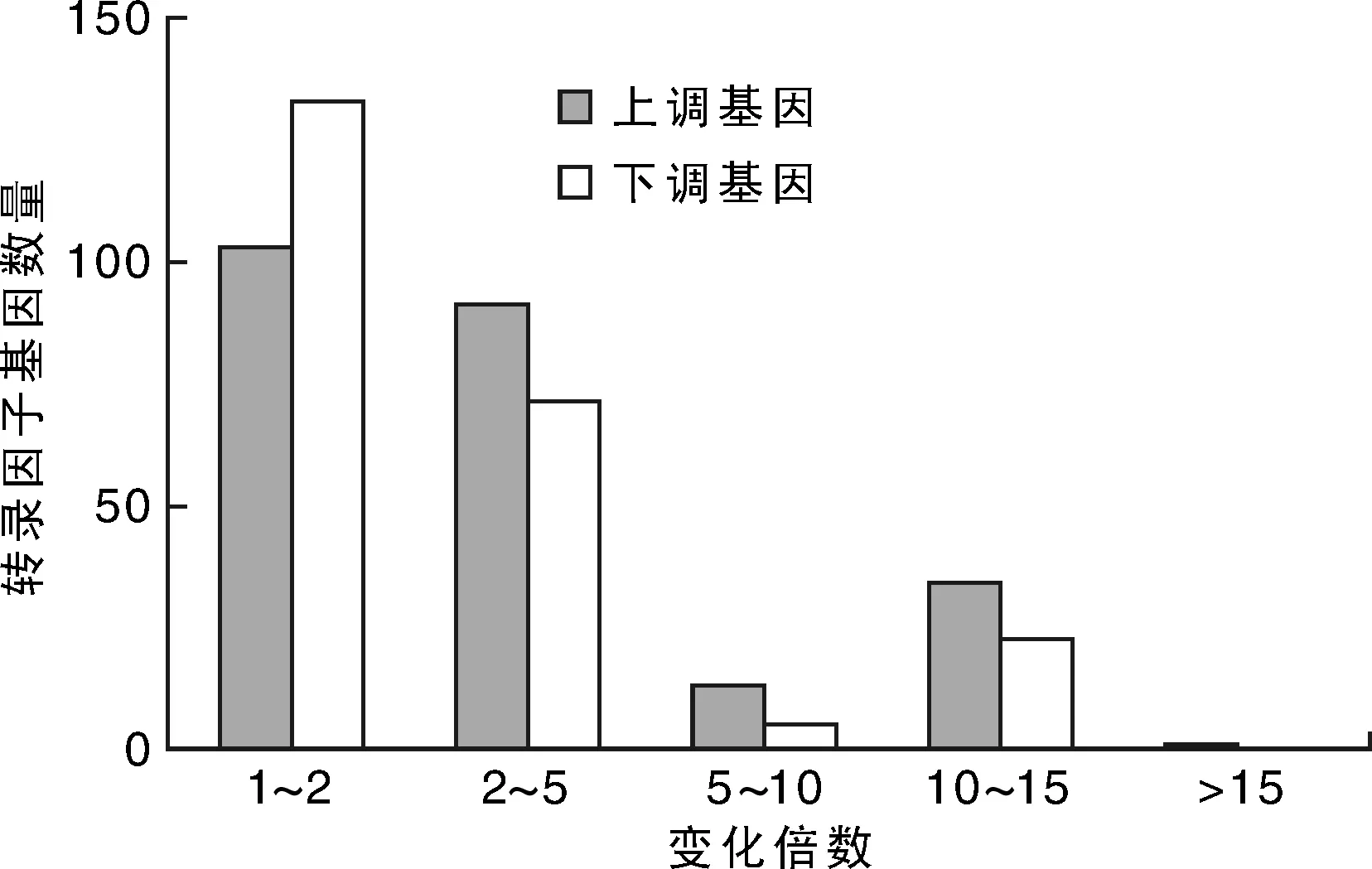
图2 紫花苜蓿叶片响应盐胁迫转录因子差异基因倍数变化Fig.2 Fold change of differentially expressed genes of transcription factor in the leaves of alfalfa under control and salt stress
图3可以看出:紫花苜蓿叶片响应盐胁迫下有46个转录因子家族的差异表达。其中差异表达显著且包含转录因子家族数量较多的家族有MYB上调表达的转录因子最多为51个,且下调表达转录因子也最多为39个。E2F-DP,FAR1,GeBP和PLATZ转录因子家族基因全部上调,BES1,C2C2-GATA,RWP-RK,S1Fa-like,SBP,Sigma70-lik,SRS和TUB转录因子家族基因全部下调。其他的转录因子家族基因如AP2-EREBP,bHLH,NAC,MADS,Trihelix和WRKY等转录家族成员基因既有转录水平上调的,也有转录水平下调的,暗示着转录家族在盐响应中调控方式的复杂性。
2.3紫花苜蓿叶片盐胁迫应答重要转录因子基因分析
转录因子激活或抑制逆境胁迫相关基因的表达。目前研究已证实bZIP,WRKY,AP2,NAC,bHLH,DERB,MYB和C2H2等家族基因呈现在盐胁迫应答中起着重要作用。根据BLASTP 结果对表达DEGs 进行功能注释(表3),发现许多盐逆境相关的转录因子基因显著上调或下调表达,上调如乙烯响应因子MsERF110(gi|357443595和gi|357464465)、MsDREB(gi|357444089)和MsERF071(gi|357438137)、bHLH转录因子MsbHLH36(gi|357474369)、锌指转录因子MsZFP(gi|357510471和gi|357503025)、MYB转录因子(gi|657379544,gi|657378081和gi|357476493)、NAC转录因子MsNAC(gi|657404698,gi|657387760和gi|657373824)、GRAS转录因子MsGRAS(gi|357472773和gi|657400127)、Trihelix转录因子MsMGT-3a(gi|657378410)和WRKY转录因子MsWRKY(gi|657389642,gi|357512541,gi|657393579,gi|357437277,gi|657390965和gi|657375305)等。下调如Alfin-like转录因子MsAlfin-like(gi|657396279)、bZIP转录因子MsbZIP48(gi|657376782)、GRAS转录因子MsGRAS(gi|357488941和gi|657390120)、MYB转录因子MsMYB(gi|657404680,gi|657400077和gi|657370137)。
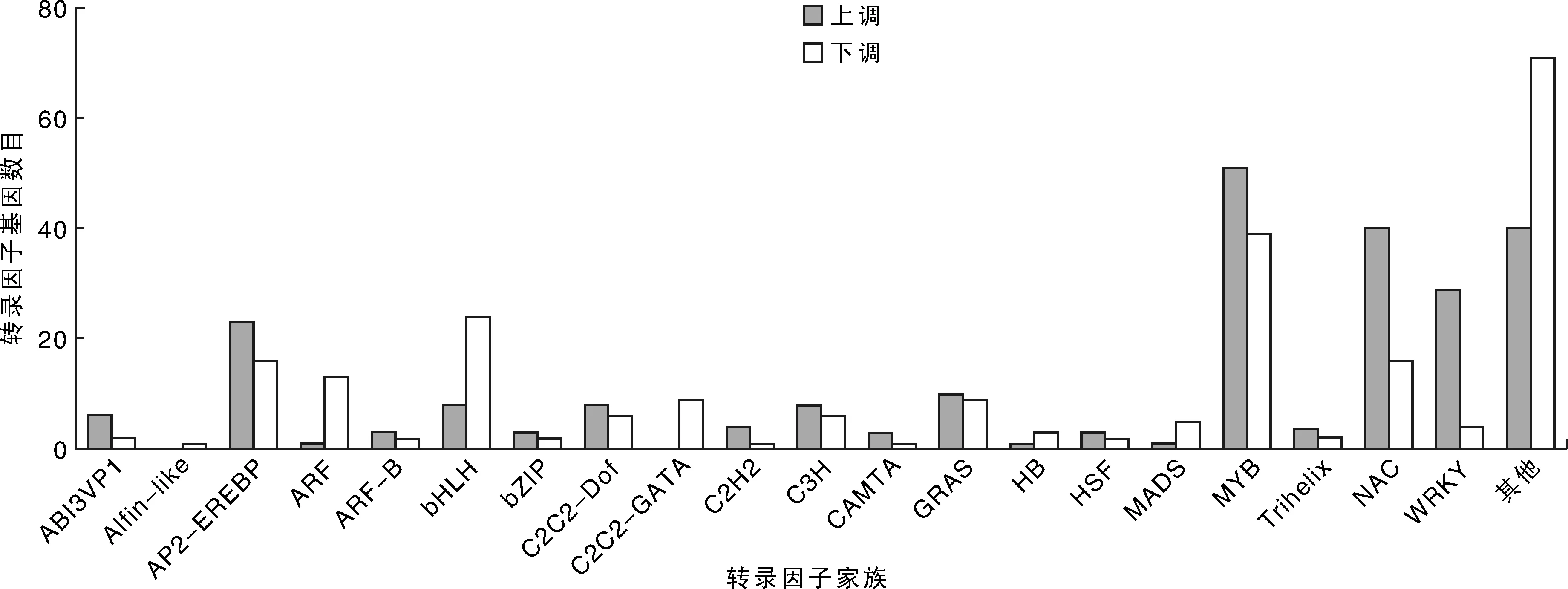
图3 紫花苜蓿叶片响应盐盐胁迫转录因子家族分布Fig.3 Distribution of transcription factor family in the leaf of alfalfa under control and salt stress
表3紫花苜蓿叶片响应盐胁迫诱导表达重要转录因子
Table 3Salt-stress induced key transcription factors in the leaf of alfalfa
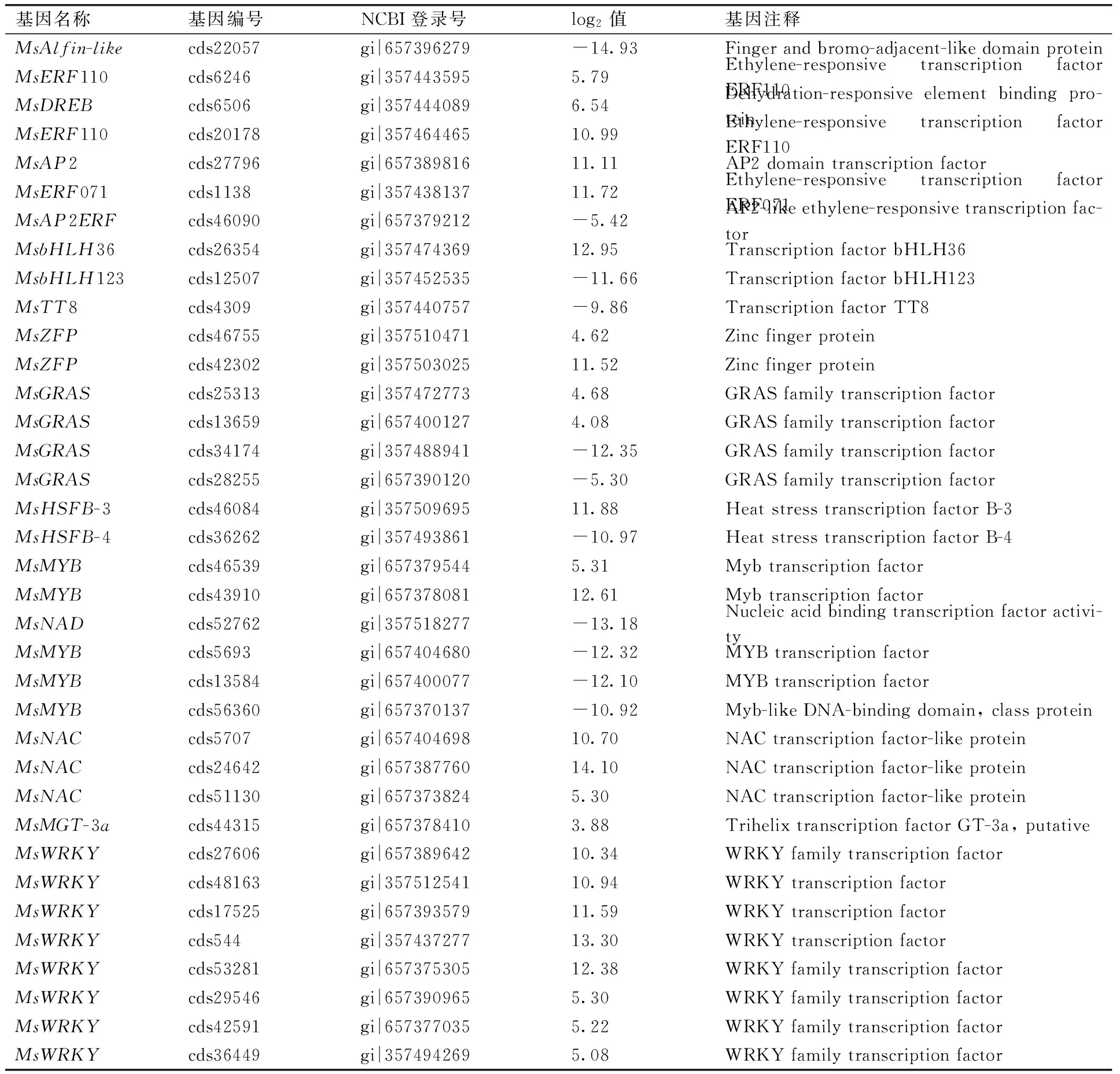
基因名称基因编号NCBI登录号log2值基因注释MsAlfin-likecds22057gi|657396279-14.93Fingerandbromo-adjacent-likedomainproteinMsERF110cds6246gi|3574435955.79Ethylene-responsivetranscriptionfactorERF110MsDREBcds6506gi|3574440896.54Dehydration-responsiveelementbindingpro-teinMsERF110cds20178gi|35746446510.99Ethylene-responsivetranscriptionfactorERF110MsAP2cds27796gi|65738981611.11AP2domaintranscriptionfactorMsERF071cds1138gi|35743813711.72Ethylene-responsivetranscriptionfactorERF071MsAP2ERFcds46090gi|657379212-5.42AP2-likeethylene-responsivetranscriptionfac-torMsbHLH36cds26354gi|35747436912.95TranscriptionfactorbHLH36MsbHLH123cds12507gi|357452535-11.66TranscriptionfactorbHLH123MsTT8cds4309gi|357440757-9.86TranscriptionfactorTT8MsZFPcds46755gi|3575104714.62ZincfingerproteinMsZFPcds42302gi|35750302511.52ZincfingerproteinMsGRAScds25313gi|3574727734.68GRASfamilytranscriptionfactorMsGRAScds13659gi|6574001274.08GRASfamilytranscriptionfactorMsGRAScds34174gi|357488941-12.35GRASfamilytranscriptionfactorMsGRAScds28255gi|657390120-5.30GRASfamilytranscriptionfactorMsHSFB-3cds46084gi|35750969511.88HeatstresstranscriptionfactorB-3MsHSFB-4cds36262gi|357493861-10.97HeatstresstranscriptionfactorB-4MsMYBcds46539gi|6573795445.31MybtranscriptionfactorMsMYBcds43910gi|65737808112.61MybtranscriptionfactorMsNADcds52762gi|357518277-13.18Nucleicacidbindingtranscriptionfactoractivi-tyMsMYBcds5693gi|657404680-12.32MYBtranscriptionfactorMsMYBcds13584gi|657400077-12.10MYBtranscriptionfactorMsMYBcds56360gi|657370137-10.92Myb-likeDNA-bindingdomain,classproteinMsNACcds5707gi|65740469810.70NACtranscriptionfactor-likeproteinMsNACcds24642gi|65738776014.10NACtranscriptionfactor-likeproteinMsNACcds51130gi|6573738245.30NACtranscriptionfactor-likeproteinMsMGT-3acds44315gi|6573784103.88TrihelixtranscriptionfactorGT-3a,putativeMsWRKYcds27606gi|65738964210.34WRKYfamilytranscriptionfactorMsWRKYcds48163gi|35751254110.94WRKYtranscriptionfactorMsWRKYcds17525gi|65739357911.59WRKYtranscriptionfactorMsWRKYcds544gi|35743727713.30WRKYtranscriptionfactorMsWRKYcds53281gi|65737530512.38WRKYfamilytranscriptionfactorMsWRKYcds29546gi|6573909655.30WRKYfamilytranscriptionfactorMsWRKYcds42591gi|6573770355.22WRKYfamilytranscriptionfactorMsWRKYcds36449gi|3574942695.08WRKYfamilytranscriptionfactor
2.4差异表达基因的qRT-PCR荧光定量验证
为了验证数字基因表达谱数据的可靠性,随机筛选4个差异表达基因,其中2个基因上调cds8203和cds4945,2个下调基因cds25725和cds27088,以苜蓿GAPDH(GenBank登录号为MTR_4g131180)为内参,进行qRT-PCR验证。结果表明,4个基因在盐胁迫处理后发生不同程度的表达(图4),但胁迫诱导表达的变化趋势相同,说明基因表达谱的分析结果可靠。
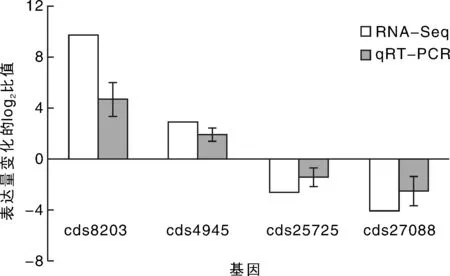
qRT-PCR 中的误差线表示平均值的标准差(n=3)。图4 DEGs 的qRT-PCR 验证Fig.4 Validation of DEGS data by qRT-PCR
3 讨论
转录因子在逆境胁迫下可以来激活或抑制下游基因的转录表达,在植物对逆境胁迫应答中充当着重要角色。目前已发现MYB,NAC,bHLH,DERB,C2H2,HD-Zip/b-ZIP和WRKY等盐胁迫相关的转录因子。本研究发现,紫花苜蓿叶片在250 mmol·L-1NaCl胁迫下72 h时,表达量差异达到2倍以上的基因共7 497个,其中包含46个转录家族474个转录因子,242个上调、232个下调,这也暗示着紫花苜蓿叶片盐胁迫应答各类转录因子调控方式的复杂性。
AP2/EREBP家族转录因子在植物中极其广泛,参与多种生理生化反应的信号诱导相关。目前已经证实AP2/EREBP家族中DREB类和ERF类转录因子在逆境抗性中起重要作用。如过表达TSRF1可提高水稻(Oryzasativa)对渗透和干旱胁迫的耐受性[7]。过表达AhDREB1可提高烟草(Nicotianatabacum)耐高盐胁迫能力[8]。但也有报道,ERFs类转录因子在植物逆境调控起负作用,如过表达SlERF3可降低番茄(Lycopersiconesculentum)对盐的耐受性[9]。本研究发现39个AP2/EREBP基因参与紫花苜蓿叶片相应盐胁迫应答过程,其中23个上调,16个下调。在盐胁迫下,AP2/EREBP基因表现上调和下调现象,说明不同的基因参与盐胁迫应答调控方式并不相同。
NAC是植物特有的转录因子基因家族,已有研究表明,NAC转录因子在植物发育、逆境应答、衰老和次生壁合成等众多的生物学过程中发挥着重要作用[10-11]。大量研究表明:NAC家族转录因子存在正负两种调控机制,过表达AtNAC2的转基因降低拟南芥的耐盐性[12],过表达ONAC045可提高水稻转基因植株抗旱和耐盐性[13]。本研究也发现:56个受盐胁迫诱导NAC家族转录因子,40个上调,16个下调,其中gi|657404698和gi|657387760上调10倍以上,这可能在紫花苜蓿叶片对盐胁迫应答反应中起重要的作用。
MYB转录因子是植物最大的转录因子家族之一,在植物非生物胁迫应答中具有重要作用。研究表明:超量表达JAmyb可增加水稻转基因植株的耐盐性[14],过表达MdSIMYB1可增强苹果和烟草对高盐、干旱和冷胁迫的耐受性[15]。在本研究中,我们也发现90个MYB转录家族成员中50%以上成员上调明显,推测它在紫花苜蓿响应盐胁迫过程中可能发挥着重要作用。紫花苜蓿叶片盐响应下WRKY家族转录因子中33个得到表达,其中29个WRKY基因表达上调、4个下调。WRKY基因家族各成员参与调控植物的抗逆反应及其信号转导途径的建立,在植物的生长发育和耐逆抗病过程中都发挥着极其重要的调控作用。现已证实WRKY家族转录因子存在正负两种调控机制,过表达拟南芥中的AtWRKY25和AtWRKY33能够提高植物对高盐的耐性[16],过表达棉花GhWRKY39-1能够提高转基因烟草对盐的耐受性[17],但过表达OsWRKY45-1可降低水稻对高盐的耐受性[18]。因此,WRKY基因参与盐胁迫具有复杂性,可能在紫花苜蓿盐胁迫应答中起重要作用。
bZIP转录因子家庭在植物防卫反应和逆境胁迫应答过程中起着十分重要的作用。据报道bZIP家族转录因子在植物逆境胁迫应答过程存在着正负两种调控机制,如OsbZIP12,OsbZIP46和OsbZIP71在ABA介导的干旱和盐耐受性扮演着一个重要的角色[19],但GmbZIP44,GmbZIP62和GmbZIP78作为负调控因子参与大豆的盐胁迫响应[20]。本研究发现5个bZIP转录因子被调控,其中3个上调和2个下调。
C2H2型锌指蛋白基因是植物中最重要的转录调节子家族之一,已在拟南芥和水稻基因组中分别发现C2H2型家族的基因成员有176个和189个。C2H2型转录家族因子参与植物生长发育和胁迫应答的多种细胞生化过程。如ZFP182编码一个具有转录激活特性的锌指蛋白,过表达ZFP182能够提高转基因烟草和水稻的耐盐水平[21]。过表达ZFP245和ZFP179可以使植物抗氧化能力显著增强,活性氧ROS清除能力提高,植物体内对盐胁迫应答中发挥重要作用[22-23]。本研究发现5个C2H2转录因子被调控,其中4个上调和l个下调,其中gi|357503025上调11倍以上。
bHLH转录因子家庭是植物中仅次于MYB转录因子的第二大转录因子超家族。拟南芥和水稻中发现有162和167个bHLH转录家族成员。bHLH转录因子能与E-box顺式作用元件特异性结合,调控胁迫应答相关基因的表达。过表达bHLH122和OrbHLH001分别可增加拟南芥和水稻的耐盐性[24-25],共表达AtWRKY28和AtbHLH17能显著提高植物对多种非生物逆境胁迫的耐受性[26]。本研究发现32个bHLH转录因子参与紫花苜蓿叶片响应盐胁迫应答过程,8个上调,24个下调,其中gi|357474369上调12倍以上,推测这些bHLH转录因子可能在紫花苜蓿盐胁迫应答过程中起重要调控作用。
Trihelix转录因子家族是一类最近才被引发关注的基因家族,拟南芥和水稻中发现有30个和31个Trihelix转录家族成员。研究显示Trihelix转录因子在非生物逆境应答中发挥重要作用[27-28]。本研究中也发现6个Trihelix转录因子差异表达。热激转录因子HSF是一类已知的有抗热功能的基因,可调节热击基因的表达,在植物对环境胁迫的分子响应中发挥重要作用。Friedberg等[29]首次克隆出MsHSFA4基因,其表达受热和冷胁迫诱导。过表达胡杨(Populuseuphratica)PeHSF可增强转基因烟草幼苗耐盐性[30]。本研究检测到3个HSF转录因子上调表达,推测HSF家族的转录因子在盐胁迫应答中发挥着一定的作用。
本研究利用RNA-Seq鉴定出474个转录因子参与南方型紫花苜蓿叶片盐胁迫应答过程。转录因子基因是改良植物耐盐性的重要靶标基因,但目前只有少数几个紫花苜蓿耐盐转录基因被克隆,因此,挖掘并克隆转录因子耐盐关键优异基因,并明确其控制下游靶基因及其在信号传导通路中作用机制还需要进一步深入的研究。
[1]MILLER G, STEIN H, HONIG A. Responsive modes ofMedicagosativaproline dehydrogenase genes during salt stress and recovery dictate free proline accumulation [J].Planta, 2005,222(1):70-79.
[2]LUO Y, LIU Y B, DONG Y X, et al. Expression of a putative alfalfa helicase increases tolerance to abiotic stress inArabidopsisby enhancing the capacities for ROS scavenging and osmotic adjustment [J].JournalofPlantPhysiology, 2009,166(4): 385-394.
[3]LONG R C, YANG Q C, KANG J M, et al. Overexpression of a novel salt stress-induced glycine-rich protein gene from alfalfa causes salt and ABA sensitivity inArabidopsis[J].PlantCellReports, 2013,32(8): 1289-1298.
[4]MA L C, WANG Y, LIU W, et al. Overexpression of an alfalfa GDP-mannose 3, 5-epimerase gene enhances acid, drought and salt tolerance in transgenicArabidopsisby increasing ascorbate accumulation [J].BiotechnologyLetters, 2014,36(11): 2331-2341.
[5]ZAHAF O, BLANCHET S, ZELICOURT A, et al .Comparative transcriptomic analysis of salt adaptation in roots of contrastingMedicagotruncatulagenotypes [J].MolecularPlant, 2012,5(5):1-14.
[6]马进, 郑钢,蔡建国. 南方型紫花苜蓿基因型在组织培养条件下对NaCl的耐性[J]. 浙江农业学报, 2011, 23(4):782-786.
[7]QUAN R D, HU S J, ZHANG Z L, et al. Overexpression of an ERF transcription factor TSRF1 improves rice drought tolerance[J].PlantBiotechnologyJournal, 2010, 8(4): 476-488.
[8]SHEN Y G, ZHANG W K, YAN D Q, et al. Characterization of a DRE-binding transcription factor from a halophyteAtriPlexhortensis[J].TheoreticalandAppliedGeneitics, 2003, 107(1): 155-161.
[9]PAN I C, LI C W, SU R C,et al. Ectopic expression of an EAR motif deletion mutant of SlERF3 enhances tolerance to salt stress andRalstoniasolanacearumin tomato [J].Planta, 2010, 232(5):1075-1086.
[10]PURANIK S, SAHU P P, SRIVASTAVA P S,et al. NAC proteins: regulation and role in stress tolerance [J].TrendsinPlantScience, 2012, 17 (6):369-381.
[11]NAKASHIMA K, TAKASAKI H, MIZOI J, et al. NAC transcription factors in plant abiotic stress responses [J].BiochimicaetBiophysicaActa, 2012, 1819 (2): 97-103.
[12]BALAZADEH S, SIDDIQUI H, ALLU A D, et al. A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence [J].ThePlantJournal, 2010, 62(2): 250-264.
[13]SONG S Y, CHEN Y, CHEN J, et al. Physiological mechanisms underlyingOsNAC5-dependent tolerance of rice plants to abiotic stress [J].Planta, 2011, 234(2): 331-345.
[14]YOKOTANI N, ICHIKAWA T, KONDOU Y, et al.Role of the rice transcription factorJAmybin abiotic stress response [J].JournalofPlantResearch, 2013, 126(1): 131-139.
[15]WANG R K, CAO Z H, HAO Y J. Overexpression of a R2R3 MYB geneMdSIMYB1 increases tolerance to multiple stresses in transgenic tobacco and apples [J].PhysiologiaPlantarum, 2014, 150(1): 76-87.
[16]JIANG Y Q, DEYHOLOS M K. Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses [J].PlantMolecularBiology, 2009, 69(1-2): 91-105.
[17]SHI W, HAO L, LI J, et al. TheGossypiumhirsutumWRKY geneGhWRKY39-1 promotes pathogen infection defense responses and mediates salt stress tolerance in transgenicNicotianabenthamiana[J].PlantCellReports, 2014, 33(3): 483-498.
[18]TAO Z, KOU Y J, LIU H B, et al.OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice [J].JournalofExperimentalBotany, 2011, 62(14): 4863-4874.
[19]LIU C T, MAO B G, OU S J, et al.OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice [J].PlantMolecularBiology, 2014, 84(1-2): 19-36.
[20]LIAO Y, ZOU HF , WANG H W, et al. SoybeanGmMYB76,GmMYB92 andGmMYB177 genes confer stress tolerance in transgenicArabidopsisplants [J].CellResearch,2008, 18(10): 1047-1060.
[21]HUANG J, YANG X, WANG M M, et al. A novel rice C2H2-type zinc finger protein lacking DLN-box/EAR-motif plays a role in salt tolerance [J].BiochimicaetBiophysicaActa, 2007, 1769(4): 220-227.
[22]HUANG J, SUN S J, XU D Q, et al. Increased tolerance of rice to cold, drought and oxidative stresses mediated by the overexpression of a gene that encodes the zinc finger protein ZFP245[J].BiochemicalandBiophysicalResearchCommunications, 2009,389(3): 556-561.
[23]SUN S J, GUO S Q, YANG X, et al. Functional analysis of a novel Cys2/His2-type zinc finger protein involved in salt tolerance in rice [J].JournalofExperimentalBotany, 2010, 61(10): 2807-2818.
[24]LIU W, TAI H, LI S, et al. bHLH122 is important for drought and osmotic stress resistance inArabidopsisand in the repression of ABA catabolism [J].NewPhytologist, 2014,201(4): 1192-204.
[25]LI F, GUO S, ZHAO Y, et al. Overexpression of a homopeptide repeat-containing bHLH protein gene (OrbHLH001) from Dongxiang Wild Rice confers freezing and salt tolerance in transgenicArabidopsis[J].PlantCellReports,2010, 29(9): 977-86.
[26]BABITHA KC, RAMU SV, PRUTHVI V, et al. Co-expression ofAtbHLH17 andAtWRKY28 confers resistance to abiotic stress inArabidopsis[J].TransgenicResearch,2013, 22 (2): 327-341.
[27]FANG Y J, XIE K B, HOU X, et al. Systematic analysis of GT factor family of rice reveals a novel subfamily involved in stress responses [J].MolecularGeneticsGenomics, 2010, 283(2): 157-169.
[28]WANG X H, LI Q T, CHEN H W, et al. Trihelix transcription factor GT-4 mediates salt tolerance via interaction with TEM2 inArabidopsis[J].BMCPlantBiology, 2014, 14∶339-345.
[29]FRIEDBERG J N, BOWLEY S R, MCKERSIE B D. Isolation and characterization of class A4 heat shock transcription factor from alfalfa [J].PlantScience,2006, 171(3):332-344.
[30]SHEN Z D, DING M Q, SUN J, et al. Overexpression ofPeHSFmediates leaf ROS homeostasis in transgenic tobacco lines grown under salt stress conditions [J].PlantCell,TissueandOrganCulture,2013,115(3):299-308.
(责任编辑张韵)
Identification and analysis of salt stress-responsive transcription factor in leaf of southern type alfalfa
PEI Cui-ming, ZHANG Zhen-ya, MA Jin*
(SchoolofLandscapeArchitecture,ZhejiangAofFUniversity,Lin’an311300,China)
Illumina RNA-sequencing was performed in two alfalfa samples of control (WT_CK2) and NaCl-treated samples(WT_N2)in order to estimate a broad spectrum of transcription factor genes affected by salt stress,and the real-time fluorescent qRT-PCR technique was used to verify the expression characteristics of random selected 4 genes. These results indicated that 7 497 of them showed two fold higher or lower difference under NaCl stress from the control, including 474 transcription factors (TFs) belonging to 46 transcription family, among which 242 genes were up-regulated and 232 were down-regulated. Transcription factor families including bHLH,NAC,WRKY and C2H2, among which more than 70% genes were up-regulated. Expression patterns of random selected 4 genes confirmed the results of the digital gene expression profile. In addition, this study also identified a large number of candidate functional transcription factor genes such asMsERF110,MsERF071,MsbHLH36,MsZFP,MsHSFB-3,MsMYB,MsNAC,MsWRKYand so on, which appeared to be constitutively involved in the response to salt stress. This study revealed that many transcription factor family were involved in response to salt stress in leaf of southern type alfalfa.
southern type alfalfa; transcription factor; leaf transcriptome; salt stress
10.3969/j.issn.1004-1524.2016.04.02
2015-09-18
国家自然科学基金资助项目(31272494)
裴翠明 (1991—),女,河南商丘人,硕士研究生,研究方向为园林植物研究和应用。E-mail: 1158862079@qq.com
,马进,E-mail: majinzjl@163.com
S542
A
1004-1524(2016)04-0550-08
裴翠明,张振亚,马进. 南方型紫花苜蓿叶片盐胁迫应答转录因子鉴定与分析[J].浙江农业学报,2016,28(4): 550-557.
猜你喜欢
做人与处世(2022年6期)2022-05-26
疯狂英语·新阅版(2022年5期)2022-04-29
今日农业(2021年21期)2021-11-26
河南农业科学(2020年2期)2020-03-11
上海农业学报(2016年2期)2016-10-27
浙江农业学报(2016年7期)2016-06-15
化工进展(2015年6期)2015-11-13
高中生学习·高三版(2014年3期)2014-04-29
食品工业科技(2014年15期)2014-03-11
中国糖料(2013年4期)2013-03-02
