
基于F2群体的豁眼鹅豁眼性状遗传分析
2016-10-13 20:41于金成
中国农业科学 2016年19期
于金成,李 喆,于 宁,赵 辉
基于F2群体的豁眼鹅豁眼性状遗传分析
于金成,李 喆,于 宁,赵 辉
(辽宁省农业科学院草牧业研究所,沈阳 110161)
【目的】豁眼鹅产蛋性能优良,是中国宝贵的地方家禽资源。作为豁眼鹅的品种标志,豁眼性状的遗传规律有待揭示。文章通过构建鹅豁眼性状F2资源群,结合表型分析,验证决定豁眼性状基因为隐性遗传的假设,从而了解豁眼性状的遗传机制,为豁眼鹅遗传资源利用提供理论依据。【方法】选用20只豁眼公鹅和100只豁眼母鹅为亲本,组建随机交配群产生豁眼鹅纯系F1代,观察F1代中眼睑的表现和分离比例;资源群采用远交群体F2设计,选用豁眼鹅(3只,15只♀)和四川白鹅(3只,15只♀)为亲本建立资源群,正反交交配产生F1代,F1代在避开近交的前提下互交产生F2代,观察资源群F1和F2代中眼睑的表现和分离比例。【结果】豁眼鹅纯系随机交配下一代的豁眼表型比例为89%(n=444),11%(n=444)的个体为正常眼睑表型,其中,豁眼与正常表型公鹅的实际比值为7﹕1(n=238),与理论值差异不显著(χ2=2.09<χ20.05(1)=3.84),母鹅的实际比值为10﹕1(n=206),与理论值差异不显著(χ2=0.06<χ20.05(1)=3.84),表明决定豁眼性状的基因为隐性遗传的假设为正确的;同时,提示豁眼性状可能由两个基因座决定的,其中一个基因座影响眼睑形成,另一个起修饰作用的基因座影响豁眼表型的外显率。②豁眼鹅与四川白鹅的反交F1代群体中公鹅和母鹅全部为正常眼睑,表明豁眼性状相对正常眼睑为隐性遗传。③正交F1代中公鹅100%(n=71)为正常眼睑,母鹅中83%(n=76)的个体表现豁眼,17%(n=76)的个体表现正常,其中,豁眼母鹅与正常表型母鹅的实际比值为5﹕1(n=76),与理论值差异不显著(χ2=3.51<χ20.05(1)=3.84),表明豁眼性状呈伴性遗传。④正交F2群体中豁眼公、母鹅与正常眼睑实际比值分别为5﹕8(n=102)和2﹕3(n=94),与理论比值差异不显著(χ2=0.36, 0.02<χ20.05(1)=3.84);同时,反交F2群体中豁眼公、母鹅与正常眼睑的实际比值分别为0﹕1(n=61)和5﹕7(n=60),与相应的理论比值差异不显著(χ2=0.02<χ20.05(1)=3.84),正反交F2群体中公母鹅的豁眼表型分离情况进一步证实了豁眼性状呈伴性隐性遗传的遗传规律。【结论】豁眼性状相对正常眼睑为隐性遗传,且呈伴性遗传;豁眼性状的形成主要受两个基因座的影响,一个起主要作用的基因座位于Z染色体上,另一个修饰作用的基因座位于常染色体上。
豁眼性状;隐性遗传;伴性遗传;鹅F2资源群;豁眼鹅
0 引言
【研究意义】豁眼鹅是中国宝贵的地方家禽遗传资源,因优良的产蛋性能而著称于世。豁眼鹅出壳时左右双侧上眼睑残缺,终生保持不变,豁眼鹅因此而得名。豁眼虽不是与生产性能直接相关的经济性状,但作为品种标志,揭示豁眼性状的遗传规律,可为豁眼鹅的遗传资源保护、纯种选育和杂交利用提供科学的参考依据。【前人研究进展】自上世纪伊始孟德尔遗传定律被重新发现以来,众多研究者采用孟德尔的研究方法,即通过建立F2资源群以研究家禽品种特征性状的遗传规律,取得了众多的成果,包括鸡的冠型[1-2]、羽型[2]、羽色和胫色[2-7]、皮肤颜色[8-9]、蛋壳颜色[10-11],鸭的羽色[12-14]、蛋壳颜色[15-17]以及鹌鹑的羽色[18-19]等等。鹅质量性状的遗传方式分析主要有羽色和斑纹、羽毛变异和解剖形态变异等方面[20],对于鹅豁眼性状的研究鲜有报道,仅朱峰伟等[21]利用RAPD分子标记技术对五龙鹅(豁眼鹅)豁眼组和无豁眼组群体进行了遗传多样性分析,其关注点是建立五龙鹅(豁眼鹅)品种内两群体的特异性分子标记,但未对该性状的遗传方式进行研究。【本研究切入点】本质上讲,豁眼是遗传缺陷,属于质量性状。通过观察豁眼鹅上眼睑的形态结构,对照正常眼睑表型,推测豁眼是鹅在胚胎发育过程中上眼睑形成基因发生突变或其表达受到影响所致,即豁眼鹅是眼睑性状的突变体。1994年,WILKIE[22]在综述显性突变的分子机制时,提出突变体多数呈隐性遗传(90%以上)的规律。据此,假设决定豁眼性状的基因为隐性遗传,根据孟德尔遗传定律,豁眼鹅与眼睑正常的鹅交配产生的F1代均为正常眼睑。以豁眼鹅和正常眼睑的四川白鹅为亲本,采用F2群体设计建立鹅豁眼性状资源群,检验豁眼的表现和分离情况是否符合孟德尔遗传规律,从而揭示豁眼性状的遗传方式。【拟解决的关键问题】通过对豁眼鹅及其资源家系上眼睑表型的观测,找出其变化规律,阐明该性状的遗传机制,以期为豁眼鹅遗传资源利用提供理论支撑。
1 材料与方法
1.1 试验鹅群的建立
选用左右双侧上眼睑残缺(本文中简称豁眼)的豁眼鹅(Huoyan goose, H)18只(♂3,♀15)和四川白鹅(Sichuan White goose, S)18只(♂3,♀15)为亲本,正反交交配产生F1代,F1代在避开近交的前提下互交产生F2代,其中,于正交F1代中挑选带有豁眼表型的母鹅个体进行选配。此外,选用20只雄性豁眼鹅和100只雌性豁眼鹅为亲本,组建随机交配群,观察豁眼性状在本品种内的传递情况。随机交配群、资源群亲代和F1代交配的公母比为1﹕5,每组6只鹅单栏饲养。豁眼鹅纯系随机交配群F1代和豁眼性状F2资源群体的构建,于2013年3月至2015年8月在辽宁省彰武县隆江牧业种鹅场实施。
1.2 杂交方法
资源群配种时采用远交群体F2设计。亲代(P)豁眼鹅与四川白鹅按照公母比例1﹕5进行正反交交配,产生2组杂交1代(F1)代群体,每组F1代群体内避半同胞交配产生杂交2代(F2),即为最终的分离群体。
群体及大小如图1所示,以豁眼鹅和四川白鹅为亲本,豁眼♂×四川白鹅♀定为正交组,四川白鹅♂×豁眼♀则为反交组,主要用于豁眼性状分离的研究。
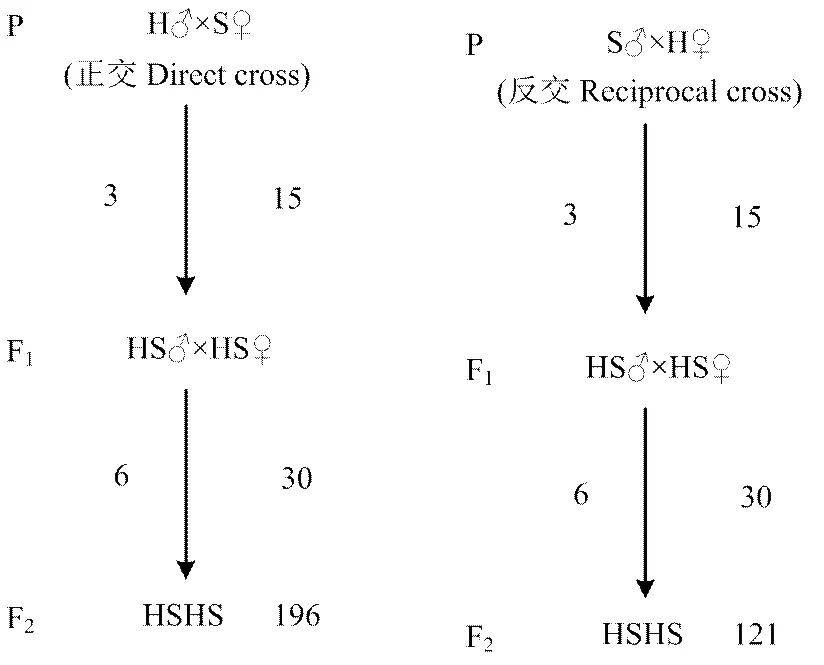
图1 资源群体继代示意图
1.3 性状测定方法
鹅在同一栋舍内分组饲养,自由采食、饮水。按常规方法进行饲养管理及免疫。出雏时即测定实验鹅的豁眼性状情况。
记录资源群中每代不同性状个体数量,Excel软件进行统计分析。性状的分析采用卡方适合性检验,用来检验性状的观察值次数与该性状的理论比率是否相符合。
当df=1,采用英国叶斯(F. Yates)提出的矫正公式。矫正后的χ2公式如下:
χ2=
其中,O:实际观察值;E:理论值;:表型数。
卡方适合性检验的无效假设:H0:实际次数和理论次数的偏差等于零,H1:实际次数和理论次数的偏差不等于零。如果>0.05,差异不显著,即理论值与实际值相符合;若0.01<<0.05,差异显著,<0.01差异极显著,说明理论值与实际值不符合。
2 结果
2.1 豁眼鹅随机交配群中豁眼性状的表现
表1显示了纯种豁眼鹅随机交配下一代群体的豁眼表型比例情况。本试验选用的亲本豁眼鹅均为双侧豁眼表型,根据豁眼基因呈隐性遗传的假设,只有豁眼基因隐性纯合体才有豁眼表型,其交配的下一代应该100%表现豁眼。本试验的结果中, F1代中89%(394/444)的豁眼鹅个体为豁眼表型,虽然比预测的豁眼比例少11%(50/444),但不能推翻决定豁眼性状的基因为隐性遗传的假设,因为从基因型到表型,会出现不完全外显的现象。
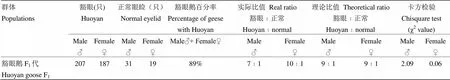
表1 豁眼鹅本品种内随机交配F1代群体中豁眼性状情况
更为重要的是,豁眼性状的不完全外显现象提示,豁眼可能是由两个基因座相互作用而导致的。两个基因座中,一个基因座影响眼睑形成,是豁眼性状形成的主要因素,将其命名为H基因座,其中,H代表正常眼睑等位基因,h代表豁眼等位基因;另一个基因座起修饰作用,是决定豁眼表型外显的成因,命名为M基因座,其中,M代表显性的豁眼表型修饰基因,m代表隐性的修饰基因。由于本试验中豁眼鹅在H基因座上是hh纯合型,所以,豁眼鹅群体中起修饰作用的M基因座上两个等位基因的频率显而易见,F1代中11%的个体为正常眼睑表型,可知m的频率约为0.33,则M等位基因的频率为0.67。据此,F1代中豁眼与正常表型鹅数的理论比值约为9﹕1,实际公鹅的比值为7﹕1,与理论值差异不显著(χ2=2.09<χ20.05(1)= 3.84);母鹅的实际比值为10﹕1,与理论值差异不显著(χ2=0.06<χ20.05(1)=3.84)。
2.2 豁眼鹅与四川白鹅正反交交配F1和F2代豁眼性状的表现
表2显示了豁眼性状F2资源群的豁眼表型比例情况。资源群亲本中豁眼鹅为豁眼表型,四川白鹅为正常眼睑,根据决定豁眼性状的基因为隐性遗传的假设,预测资源群F1代个体均为正常眼睑。杂交结果显示,豁眼鹅与四川白鹅的反交F1代群体中公鹅和母鹅全部为正常眼睑,表明豁眼性状相对正常眼睑为隐性遗传,证实了笔者关于决定豁眼性状的基因为隐性遗传的假设。
正交F1代中公鹅100%为正常眼睑,母鹅表现豁眼,表明豁眼性状呈伴性遗传。根据纯种豁眼鹅F1代中母鹅的豁眼比例情况(表1),即豁眼鹅纯系中m等位基因频率为0.33,推测正交F1代中母鹅的豁眼与正常眼睑的理论比值为9﹕1,实际比值为5﹕1,与理论比值差异不显著(χ2=3.51<χ20.05(1)=3.84)。该群体中表现正常眼睑的母鹅比例为17%,则正交F1代母鹅群体中m基因频率为0.41(),高于豁眼鹅纯系群体的m基因频率(0.33),提示四川白鹅群体的m基因频率可能高于豁眼鹅。究其原因,豁眼性状呈伴性遗传,则正交F1代中母鹅群体的H基因座都是豁眼鹅提供的h等位基因,且眼睑正常母鹅的M基因座是mm纯合型,根据群体遗传学原理,豁眼鹅和四川白鹅两群体杂交产生F1代的m基因频率为两群体该基因频率的算术平均数,已知正交F1代母鹅群体m等位基因频率为0.41,豁眼鹅纯系群体的m等位基因频率为0.33,则四川白鹅群体的m等位基因频率约为0.49(0.41×2-0.33=0.49),这可能与豁眼鹅历经多年的本品种选育有关,在逐步淘汰非豁眼鹅过程中降低了m等位基因的频率,而四川白鹅在该基因座上未受到选择。
由于正交F2群体来源于F1,则F1群体中豁眼与正常眼睑母鹅的实际比值5﹕1,可作为F2群体该指标的基础理论值,根据孟德尔遗传定律,推测,正交F2群体中豁眼与正常眼睑的公、母鹅数理论比值都应为5﹕7,而实际比值分别为5﹕8和2﹕3,经χ2检验差异不显著(χ2=0.36, 0.02<χ20.05(1)= 3.84);同时,反交F2群体中理论比值分别为0﹕1和5﹕7,实际值为0﹕1和5﹕7,经χ2检验差异不显著(χ2=0.02<χ20.05(1)=3.84)。正反交F2群体中公、母鹅的豁眼表型分离情况进一步证实了豁眼性状伴性隐性遗传的遗传规律。

表 2 豁眼鹅与四川白鹅杂交F1和F2代群体中豁眼性状分离
HS组为豁眼公鹅与四川白鹅母鹅交配组,SH组为四川白鹅公鹅与豁眼鹅母鹅交配组。HSHS为豁眼公鹅与四川白鹅母鹅交配组的F1代交配组,SHSH组为四川白鹅公鹅与豁眼鹅母鹅交配组的F1代交配组。表格中*表示不需计算卡方值
Group HS stands for that female of SichuanWhite goose mated to gander of Huoyangoose. Group SH stands for that female of Huoyan goose mated to gander of SichuanWhite goose. Group HSHS stands for that females of the Group HS mated to males of the Group HS without inbred. Group SHSH stands for that females of the Group SH mated to males of the Group SH without inbred. * stands for that value of χ2don’t need to calculate
3 讨论
3.1 豁眼性状呈隐性遗传
通过鹅豁眼性状F2资源群的建立,对其表型分离情况进行了详细的统计分析,结合豁眼鹅本品种内随机交配群体中豁眼表型数据,证实了笔者关于决定豁眼性状基因为隐性遗传的假设。本实验中豁眼鹅随机交配下一代中公、母鹅共有11%的个体表现正常眼睑表型,这可能是豁眼鹅纯系的不同群体中普遍存在的现象。2009年,为了建立五龙鹅(豁眼鹅)本品种内豁眼组和无豁眼组两群体的特异性分子标记,朱峰伟等[21]利用RAPD分子标记技术对五龙鹅(豁眼鹅)本品种内群体进行了遗传多样性分析,该研究隐含的意思是,豁眼鹅群体随机交配一代后会出现一些带有正常眼睑表型的个体,形成了五龙鹅(豁眼鹅)豁眼组群体和无豁组群体。
正常眼睑表型的豁眼鹅并非在基因型上与眼睑缺失的豁眼鹅有差别,而是基因型到表型过程中,发生了不完全外显导致的。一些人类遗传性疾病的临床表现中经常出现不完全外显现象,且多发生于由显性基因控制的疾病中,其形成的原因众多,包括前突变、限性因素、等位基因的影响、修饰基因的作用、二基因遗传(digenic inheritance)、基因印记和环境因素等[23]。但也有部分的不完全外显现象是由隐性基因控制的,其机制是主效基因突变造成基因表达异常所致[24]。豁眼性状由两个基因座相互作用而导致的,一个基因座影响眼睑形成,是豁眼性状形成的主要因素,当该基因座隐性纯合时才有可能导致眼睑发育不全,但是,另一个起修饰作用的基因座隐性纯合时却会抑制豁眼表型的形成。可见,豁眼性状形成的遗传基础复杂而有趣,其遗传机制有待于借助现代分子生物学手段来揭示。
3.2 豁眼性状呈伴性遗传
首次发现豁眼性状的伴性遗传现象。自Morgan等[25]详尽描述鸡横斑芦花伴性遗传以来,在家鸡中先后发现了金银羽、羽速、矮小型等伴性性状[26],在鹌鹑中发现羽色的伴性遗传[27],如今,这些性状广泛应用于现代蛋鸡、肉鸡和鹌鹑生产中。鹅的质量性状中,由灰雁驯化而来的一些欧洲鹅的羽色和斑纹性状呈伴性遗传[28-29],但尚未在由鸿雁驯化而来的中国地方鹅中发现该现象。豁眼鹅是中国产蛋性能最优良的鹅种,豁眼的伴性遗传在鹅的杂交利用中可用于1日龄雏鹅自别雌雄,其应用前景良好。
作为特征性状,豁眼表型曾是民间选种的依据。辽宁省昌图地区的豁眼鹅是由“闯关东”的山东移民所带来,当地群众根据“鹅有豁,产蛋多”的标准,历经20余年选育了东北地区的豁眼鹅群体[30]。民间选种是自发行为,豁眼与产蛋只是一种表型关联,并无实质联系。豁眼相对正常眼睑为隐性遗传,隐性性状作为选种的标准往往是对显性表型的淘汰来完成,这不能解释民间成功选育豁眼鹅的现象。正是豁眼的伴性遗传方式,才促使豁眼鹅种群在东北地区快速重建和扩散。因此,豁眼性状遗传规律的揭示可为豁眼鹅的遗传资源保护、纯种选育和杂交利用提供科学的参考依据。
3.3 豁眼性状遗传规律分析
豁眼性状属于质量性状,它是鹅胚在发育过程中上眼睑形成不全导致的一种表征遗传缺陷。推测,这种缺陷是上眼睑发育基因本身发生了突变或或其表达受到影响所致。本研究表明,鹅豁眼性状由隐性基因控制,呈伴性遗传;豁眼性状由两个基因座相互作用形成。因此,豁眼性状可能的分子机制是:在Z染色体上存在1个影响上眼睑发育的基因座H,该基因座上隐性等位基因纯合(hh)导致上眼睑发育基因的表达异常;在常染色体上还存在1个起修饰作用的基因座M,该基因座上隐性等位基因纯合(mm)能够影响豁眼表型的外显率。
根据孟德尔遗传规律,下面针对豁眼鹅和四川白鹅杂交F1代和F2代的眼睑性状进行遗传模式分析。豁眼鹅为眼睑部分缺失,公鹅的基因型为ZhZhMM或ZhZhMm,母鹅的基因型为ZhWMM或ZhWMm;四川白鹅为正常眼睑,公鹅基因型为ZHZHMM或ZHZHMm或ZHZHmm,母鹅基因型为ZHWMM或ZHWMm或ZHWmm。正交组合的遗传模式如图2所示,基因型为ZhZhM(M/m)的豁眼公鹅与基因型为ZHW(M/m)(M/m)的四川白鹅母鹅交配,F1代群体中,带有H等位基因的公鹅,无论M基因座是何种等位基因,均表现正常眼睑;由于豁眼鹅群体中m基因频率约为0.33,四川白鹅群体中m基因频率约为0.52,则5/6 的母鹅个体表现豁眼,其基因型为ZhWM(M/m);1/6的个体表现正常眼睑,其基因型为ZhWmm。F2代群体中,1/2 的公鹅表现正常眼睑,其基因型是ZHZh(M/m)(M/m);剩余的1/2 公鹅中,约5/6的个体表现豁眼,即5/12的个体基因型是ZhZhM(M/m),约1/6个体表现正常眼睑,即1/12的个体基因型为ZhZhmm,所以,F2代公鹅中豁眼表型分离比值在5﹕7左右;同理,F2代群体中豁眼母鹅与正常眼睑的母鹅数之比也应在5﹕7左右。

图2 豁眼鹅×四川白鹅群体眼睑性状的遗传模式
反交组合的遗传模式如图3所示,基因型为ZHZH(M/m)(M/m)的四川白鹅公鹅与基因型为ZhWM(M/m)的豁眼母鹅交配,F1代群体中,公鹅、母鹅均带有H等位基因,所以都为正常眼睑。F2代群体中,带有H等位基因的公鹅,无论M基因座是何种等位基因,均表现正常眼睑;1/2 的母鹅表现正常眼睑,其基因型是ZHW(M/m)(M/m);剩余的1/2 母鹅中,约5/6的个体表现豁眼,即5/12的个体基因型是ZhWM (M/m),约1/6个体表现正常眼睑,即1/12的个体基因型为ZhWmm,所以,F2代母鹅中豁眼表型分离比值在5﹕7左右。

图3 四川白鹅×豁眼鹅群体眼睑性状的遗传模式
4 结论
建立了鹅豁眼性状分化的F2资源群,分析了豁眼表型分离情况,结果表明:(1)豁眼性状相对正常眼睑性状为隐性遗传;(2)豁眼性状呈伴性遗传;(3)豁眼性状主要受Z染色体1个影响上眼睑发育的基因座所导致,此外,在常染色体上还存在1个起修饰作用的基因座能够影响豁眼表型的外显率。
References
[1] BATESON W.. Cambridge: Cambridge University Press, 1909: 61-68.
[2] DUNN L C, JULL M A. On the inheritance of some characters of the silky fowl.1927, 19(1): 27-63.
[3] PEARL R, SURFACE F M. On the inheritance of the barred color pattern in poultry.1910, 30(1): 45-61.
[4] KNOX C W. The genetics of plumage colour in poultry. Iowa Agricultral Experiment Station, 1926, Bullentin, No.105.
[5] KNOX C W. The inheritance of shank color in chickens.1935, 20(6): 529.
[6] NORDSKOG A W. Inheritance of shank and plumage color in the Fayoumi breed. P1970, 49(6): 1735-1737.
[7] 王存芳, 李宁, 吴常信. 基于F2群体的藏鸡羽色、胫色性状的遗传分析. 遗传, 2006, 28(7): 810-814.
WANG C F, LI N, WU C X. Genetic analysis of feather color and shank color trait based on F2resource population in Tibetan chicken.2006, 28(7): 810-814.
[8] BATESON W, PUNNETT R C. The inheritance of the peculiar pigmentation of the silky fowl.1911, 1(3): 185-203.
[9] LAMBERT W V, KNOX C W. Genetic studies in poultry II.1927, 7(1): 24-30.
[10] PUNNETT R C, BAILEY P G. Genetic studies in poultry. II. Inheritance of egg-colour and broodiness.1920, 10(4): 277-292.
[11] PUNNETT R C. Genetic studies in poultry. IX. The blue egg.1933, 27(3): 465-470.
[12] JAAP R G.. Alleles of the mallard plumage pattern in ducks.1934, 19(4): 310-322.
[13] GONG Y, YANG Q, LI S, FENG Y, GAO C, TU G, PENG X. Grey plumage coloration in the duck is genetically determined by the alleles on two different, interacting loci.2010, 41:105-108.
[14] LIN R L, CHEN H P, ROUVIER R, POIVEY J P. Selection and crossbreeding in relation to plumage color inheritance in three chinese egg type duck breeds ()., 2014, 27(8): 1069-1074.
[15] 赖垣忠,张松殊. 鸭蛋蛋壳颜色的遗传分析. 遗传, 1991, 13(2): 4-6.
LAI Y Z, ZHANG S S. Studies on egg shell color inheritance of the laying duck., 1991, 13(2): 4-6. (in Chinese)
[16] 陈晖, 檀俊秩, 曾安庆. 莆田黑鸭青壳蛋遗传规律的研究. 福建省农业科学院学报, 1991, 6(2): 71-74.
CHEN H, TAN J Z, ZENG A Q. Study on genotype of blue shell in Pu-tian black ducks.1991, 6(2): 71-74. (in Chinese)
[17] LIU H C, HSIAO M C, HU Y H, LEE S R, CHENG W T K. Eggshell pigmentation study in blue-shelled and white-shelled ducks.2010, 23: 162-168.
[18] LAUBER J K. Sex-linked albinism in the Japanese quail., 1964, 146(3646): 948-950.
[19] TRUAX R E, JOHNSON W A. Genetics of plumage color mutants in Japanese quail.1979, 58(1): 1-9.
[20] 陈国宏, 王继文, 何大乾, 王志跃, 王宝维. 中国养鹅学. 北京: 中国农业出版社, 2013: 43-44.
CHEN G H, WANG J W, HE D Q, WANG Z Y, WANG B W.Beijing: China Agricultural Press, 2013: 43-44. (in Chinese)
[21] 朱峰伟, 葛蔚, 王宝维, 朱新产. 五龙鹅豁眼性状遗传变异的RAPD 分析. 西北农业学报, 2009, 18(5): 53-57.
ZHU F W, GE W, WANG B W, ZHU X C. The genetic variation analysis of Huoyanmorphological characters in Wulong geese by RAPD marking technique.2009, 18(5): 53-57. (in Chinese)
[22] WILKIE A O. The molecular basis of genetic dominance.1994, 31: 89-98.
[23] ZLOTOGORA J. Penetrance and expressivity in the molecular age.2003, 5(5): 347-352.
[24] BARADARAN-HERAVI A, CHO K S, TOLHUIS B, SANYAL M, MOROZOVA O, MORIMOTO M, ELIZONDO L I, BRIDGEWATER D, LUBIENIECKA J, BEIRNES K, MYUNG C, LEUNG D, FAM H K, CHOI K, HUANG Y, DIONIS K Y, ZONANA J, KELLER K, STENZEL P, MAYFIELD C, LüCKE T, BOKENKAMP A, MARRA M A, VAN LOHUIZEN M, LEWIS D B, SHAW C, BOERKOEL C F. Penetrance of biallelic SMARCAL1 mutations is associated with environmental and genetic disturbances of gene expression.2012, 21(11): 2572-2587.
[25] MORGAN T H, Goodale H D. Sex-linked inheritance in poultry., 1912, 22(1): 113-133.
[26] 杨宁. 家禽生产学. 北京: 中国农业出版社, 2002: 38-39.
YANG N.Beijing: China Agricultural Press, 2002: 38-39.(in Chinese)
[27] 庞有志, 赵淑娟. 鹌鹑羽色遗传的研究及应用. 遗传, 2003, 25(4): 450-454.
PANG Y Z, ZHAO S J. The study and use on genetics of plumage color in quail.2003, 25(4): 450-454. (in Chinese)
[28] JEROME F N. Color inheritance in geese and its application to goose breeding.1953, 32(1): 159-165.
[29] JEROME F N. Color inheritance of geese: Study 2.1954, 33(3): 525-528.
[30] 国家畜禽遗传资源委员会组编. 中国畜禽遗传资源志(家禽志). 北京: 中国农业出版社, 2011: 528.
National Committee of Domestic Animal and poultry Genetic Resources, Compiled by the Committee.(). Beijing: China Agricultural Press, 2011: 528. (in Chinese)
(责任编辑 林鉴非)
Genetic Analysis of Huoyan Trait Based on F2Resource Population in Huoyan Goose
YU Jin-cheng, LI Zhe, YU Ning, ZHAO Hui
(Pratacultural & Animal Science Research Institute, Liaoning Academy of Agricultural Sciences, Shenyang 110161)
【Objective】The Huoyan breed of geese which possesses a good laying performance, is a precious native poultry resource of China. As the breed characteristics of Huoyan goose, they need to be studied about the inheritance patterns of Huoyan trait. The purpose of this study was, by establishing the goose-F2resource group with Huoyan trait records, combined with the phenotypic analysis, to test the hypothesis that the allele that determines Huoyan trait demonstrates recessive inheritance, so as to understand the genetic mechanism of Huoyan trait. Our findings of the study will lay a theoretical basis for utilization of Huoyan goose genetic resource.【Method】Twenty males and 100 females of Huoyan geese were chosen to generate the F1generation of Huoyan goose pure line by random mating, and Huoyan appearance and segregation ratio in F1were observed. By F2-design of outbreed populations, the resource population with Huoyan trait records was established, in which the 3 males and 15 females of Huoyan geese were crossed reciprocally with the 3 males and 15 females of Sichuan White geese, and inter se matings in F1were carried out to generate the F2population without inbreeding. Huoyan appearance and segregation ratio in F1and F2were observed.【Result】①Huoyan and normal eyelid phenotype in F1generation of Huoyan goose pure line by random mating was 89% (n=444) and 11% (n=444), respectively. The real ratio of Huoyan vs normal eyelid phenotype of males was 7﹕1(n=238), which was no significant with the theoretic value (χ2=2.09<χ20.05(1)=3.84), and the real ratio of Huoyan vs normal eyelid phenotype of females was 10﹕1(n=206), which was no significant with the theoretic value (χ2=0.06<χ20.05(1)=3.84). It indicated that the hypothesis of the allele that determines Huoyan trait demonstrates recessive inheritance is correct, and that Huoyan trait may be determined by two loci, one influences the formation of eyelid, and another modifies the penetrance of the Huoyan phenotype. ②All of the males and females in reciprocal cross F1populations of female Huoyan goose crossed with Sichuan White goose ganders were normal eyelid. It indicated that the Huoyan trait exhibits recessive heredity to normal eyelid trait. ③In F1populations of Huoyan goose gander crossed with female Sichuan White goose, 100% (n=71) of the males were normal eyelid, as well as the females with Huoyan and with normal eyelid phenotype were 83% (n=76), 17% (n=76), respectively. The real ratio of Huoyan vs normal eyelid phenotype of females was 5﹕1(n=76), which was of no significance with the theoretic value by the χ2test (χ2=3.51<χ20.05(1)=3.84). It indicated that the Huoyan trait demonstrates sex-linked inheritance. ④In F2populations of Huoyan goose ganders crossed with female Sichuan White goose, the real ratio of Huoyan vs normal eyelid phenotype of males and females were 5﹕8 (n=102), 2﹕3 (n=94), respectively, by the χ2test, and they were of no significance with the theoretic value (χ2=0.36, 0.02<χ20.05(1)=3.84). Meanwhile, in F2populations of reciprocal cross, the real ratio of Huoyan vs normal eyelid phenotype of males and females were 0﹕1(n=61), 5﹕7(n=60), respectively, by the χ2test, they were of no significance with the theoretic value (χ2=0.02<χ20.05(1)=3.84). The separation results of Huoyan phenotype in F2populations reciprocally further proved that the Huoyan trait demonstrates recessive sex-linked inheritance【Conclusion】The Huoyan trait exhibits recessive heredity to normal eyelid trait, and demonstrates sex-linked inheritance. The Huoyan trait is determined by two loci, one gene is on the Z chromosome, and another modifier gene exists on the autosome.
Huoyan trait; recessive inheritance; sex-linked inheritance; goose-F2resource population; Huoyan goose
2016-02-19;接受日期:2016-08-05
国家水禽产业技术体系专项资金(CARS-43-23)、辽宁省博士启动基金(20141164)
于金成,Tel:024-31029891;13898156386;E-mail:yujincheng_pi@126.com。通信作者赵辉,E-mail:zhaohui_sy@126.com
猜你喜欢
意林(2022年7期)2022-04-19
科普童话·学霸日记(2021年5期)2021-09-05
现代畜牧科技(2021年2期)2021-03-19
医学与法学(2020年3期)2020-09-18
刑事技术(2018年2期)2018-05-05
作文周刊·小学三年级版(2016年30期)2017-04-15
农村百事通(2017年6期)2017-03-30
小学生导刊(高年级)(2016年12期)2016-12-07
乡村科技(2015年1期)2015-02-22
中国司法鉴定(2014年2期)2014-04-19
