
茶树3类渗透调节物质与冬春低温相关性及其品种间的差异评价
2016-10-13 20:39薄晓培王梦馨王金和韩宝瑜
中国农业科学 2016年19期
薄晓培,王梦馨,崔 林,王金和,韩宝瑜
茶树3类渗透调节物质与冬春低温相关性及其品种间的差异评价
薄晓培1,王梦馨1,崔 林1,王金和2,韩宝瑜1
(1中国计量大学浙江省生物计量及检验检疫技术重点实验室,杭州310018;2江苏吟春碧芽股份有限公司,江苏丹阳212345)
【目的】探讨2个茶树品种可溶性糖、脯氨酸和游离氨基酸等3类渗透调节物质含量变化及其与冬春期间气温的相关性,评价其对低温的响应程度及其品种间的差别。【方法】选早芽品种‘平阳特早’和‘中茶102’,连续2年于10月21日至翌年4月5日逐日记载茶梢间气温,每天8:00、14:00、20:00记录干湿球温度计的温度,每日3次测得的每块茶园3支干球温度计的平均值即为当日茶梢生境气温;测温期间每月5日、15日、25日采摘成叶,以蒽酮比色法测定茶鲜叶可溶性糖含量,将剩余擦拭干净的鲜叶蒸青、烘干制成蒸青茶样,用SDE-HPLC方法测定茶干叶脯氨酸和游离氨基酸含量。【结果】根据茶树的发育起点温度和休眠温度,将试验阶段分为越冬期(2013/10/21—2014/3/5,2014/10/21—2015/3/5)和早春期(2014/3/6—2014/4/5,2015/3/6—2015/4/5)2个阶段。①连续2年试验的越冬期间(2013/10/21—2014/3/5,2014/10/21—2015/3/5),气温先逐渐下降,再持续低温;2个品种成叶的可溶性糖含量相应地升高、再升高,与旬平均气温显著负相关;脯氨酸含量升高、保持,与旬平均气温显著负相关;游离氨基酸含量则为下降、保持,与旬平均气温显著正相关;当温度出现最低值时,可溶性糖含量和脯氨酸含量相应出现最高值,游离氨基酸含量也跟随着出现最低值,而当温度出现最高值时,可溶性糖含量和脯氨酸含量随后也就相应地出现最低值,游离氨基酸含量随之也相应地出现最高值。②2年试验的早春期间(2014/3/6—2014/4/5,2015/3/6—2015/4/5)气温渐升,可溶性糖含量趋于减少,脯氨酸、游离氨基酸含量趋于增加;③茶树抗冻性与温度密切相关,茶鲜叶可溶性糖、脯氨酸和游离氨基酸含量与茶树的抗冻性密切相关,2年越冬期间,与‘中茶102’相比,在遭受冬季低温胁迫时,‘平阳特早’3类渗透调节物质对低温的响应性、响应幅度分别更灵敏、更大,‘平阳特早’3类渗透调节物质含量稍多。【结论】深秋至清明期间茶树易受冻害和倒春寒,其中越冬期间可溶性糖、脯氨酸和游离氨基酸皆能敏感地响应低温变化,三者含量分别与旬平均气温显著相关,是茶树的重要抗冻指标;‘平阳特早’品种对于低温响应程度大于‘中茶102’。
低温;茶树;冻害;可溶性糖;脯氨酸;游离氨基酸
0 引言
【研究意义】近年来,中国江北和江南茶区茶园面积持续扩展,而深秋至早春的极度严寒和“倒春寒”不期而至,常常导致大片茶园受冻[1-2]。早春日平均气温高于10℃时,茶芽开始萌动,代谢活动增强,假以时日即可采制高档茶叶。然而萌动了的茶芽抗寒性减弱,若气温骤降,则会受冻而死,延迟大批茶芽的萌发并降低萌发率,进而导致春茶减产和品质降低[3-4]。【前人研究进展】茶树具备抗冻生理生化机制,茶鲜叶脯氨酸、可溶性蛋白和可溶性糖含量随气温的降低皆呈先升后降的变化趋势,但不同茶树品种响应低温胁迫的温度范围有所不同[5]。有研究者认为可溶性糖含量高低可作为判断茶树品种或新梢抗寒性强弱的一个指标[6]。也有研究者指出冷驯化中茶树体内脯氨酸含量变化是对外界条件变化的一种综合反应,很难说明单一的脯氨酸含量与抗寒力之间的因果关系[7]。还有研究者认为可溶性糖和脯氨酸都是植物体内重要的抗寒物质[8-10]。【本研究切入点】关于脯氨酸含量与茶树抗寒性的关系,目前的研究还存在异议;而且关于游离氨基酸含量与茶树抗寒性的研究较少;茶树抗寒特早生品种的选育一直成为炙手可热的话题。为探讨茶树抗冻的物质基础,田野等[11]从2013年深秋至2014年早春研究了‘乌牛早’和‘茂绿’品种抗冻性,检测了成叶中可溶性糖、脯氨酸、游离氨基酸含量与冬春气温相关性,发现这3类物质与低温密切相关,可作为茶树“抗冻指标”,且可溶性糖含量与温度相关性更密切,‘乌牛早’抗性强于‘茂绿’。【拟解决的关键问题】本研究于2013年10月21日至2014年4月5日、2014年10月21日至2015年4月5日连年检测早芽种[12-13]‘平阳特早’和‘中茶102’可溶性糖、脯氨酸和游离氨基酸含量,分析这3类物质分别对于低温响应程度和含量变化,以及其在这2个早芽种成叶中的含量差异,比较2个品种对于低温的敏感性。
1 材料与方法
1.1 试验茶园与茶梢生境气温调查方法
2013年10月21日至2014年4月5日、2014年10月21日至2015年4月5日在中国计量大学试验茶园(北纬30°19′29.90″,东经120°21′19.16″,海拔11 m)选‘平阳特早’和‘中茶102’2个茶树品种,树龄6年,树高95 cm、树幅宽100 cm。每个品种园约为80 m2正方形,在每块试验茶园单对角线的茶丛内各放3支干湿球温度计,贴近茶梢,温度计顶端与茶芽平齐。每天8:00、14:00、20:00记录干湿球温度计的温度和湿度,每日3次测得的每块茶园3支干球温度计的平均值即为当日茶梢生境气温。
1.2 茶鲜叶3类渗透调节物质的检测
1.2.1 样品可溶性糖检测 试验期间的每月5日、15日、25日采集茶芽下第2至6片叶,用干净湿润的纱布轻轻擦去叶面灰尘。称取0.500 g鲜叶,剪碎放入具塞试管中,加10 mL蒸馏水,浸入沸水中提取1 h,冷却后3 000 r/min离心10 min,上清液即为待测液。采用蒽酮比色法测定可溶性糖含量。每个样品重复6次,平均值即为当日检测值。
1.2.2 样品氨基酸检测 将剩余擦拭干净的鲜叶蒸青、烘干制成蒸青茶样。将蒸青样研磨均匀,称取2.000 g茶粉放入100 mL具塞锥形瓶中,加入75 mL蒸馏水,摇匀,于沸水中提取1 h。冷却后,2 000 r/min离心5 min,取上清液用0.45 µm微孔滤膜过滤,吸600 µL滤液至2 mL离心管,加入0.2 mol·L-1PITC-乙腈溶液、1 mol·L-1三乙胺-乙腈溶液各200 µL,混匀,室温放置1 h。加正己烷400 µL,涡旋振荡30 s,静置10 min,取下层溶液至棕色小瓶,即为待测液。每个样品重复3次。采用高效液相色谱法测定脯氨酸和17种游离氨基酸含量。色谱条件参照田野等[11]的方法并改进。
氨基酸定量分析:用50%的乙腈将氨基酸标准品原液进行不同浓度梯度的稀释,分别稀释10倍、20倍、50倍、100倍,用高效液相色谱法测定峰面积,将不同浓度梯度的标准品按其峰面积和浓度制做成标准曲线,根据测得样品的峰面积,通过标准曲线计算出样品中氨基酸的含量。
1.3 数据处理
茶树发育起点温度是10℃,严冬来临时日平均温度低于10℃则开始休眠,早春时节高于10℃茶树便开始发芽[14],因此,将试验阶段分为越冬期(2013/10/21—2014/3/5,2014/10/21—2015/3/5)和早春期(2014/3/6—2014/4/5,2015/3/6—2015/4/5)2个阶段。
以ORIGIN软件作出渗透调节物质含量(g·g-1)-气温关系图,观察各类渗透调节物质含量随着气温的变化趋势,比较2个供试品种之间渗透调节物质含量的差异。为便于数值分析,文中各类物质含量皆用百分含量;以旬平均气温为自变量,以各类渗透调节物质含量分别为因变量,用DPS软件求出相关系数并检验显著性,建立回归方程。
2 结果
2.1 越冬期和早春期2个品种茶鲜叶可溶性糖含量动态及其与气温关系
依据茶树发育起点为10℃的标准,将越冬期分为第1阶段(2013/10/21—2013/12/5和2014/10/21—2014/12/5)和第2阶段(2013/12/6—2014/3/5和2014/12/6—2015/3/5),其中第1阶段为深秋至初冬时期,气温持续下降,日均温≥10℃;第2阶段为寒冷的严冬期间,日均温≤10℃。2013—2014年严冬期间,气温变幅剧烈,还有3 d日平均气温低于0℃,即-0.38℃(2013/12/27)、-0.93℃(2013/12/28)和-0.09℃(2014/2/10),平均气温6.42℃;2014—2015年严冬期间气温变化相对平缓且日均温均在0℃ 以上,最低温度为1.97℃(2015/1/1),平均温度6.98℃。早春期(2014/3/6—2014/4/5和2015/3/6—2015/4/5)温度呈上升趋势,日均温≥10℃,茶芽逐渐萌发(图1)。
2013—2014年越冬期间,‘中茶102’可溶性糖平均百分含量(4.19%)低于‘平阳特早’(4.68%)。第1阶段(2013/10/21—2013/12/5),‘中茶102’和‘平阳特早’可溶性糖含量逐渐上升,该阶段‘中茶102’从0.0374 g·g-1(2013/10/25)增加为0.0467 g·g-1(2013/12/5),上升了24.87%,‘平阳特早’从0.0283 g·g-1(2013/10/25)增加为0.0478 g·g-1(2013/12/5),上升了68.90%。第2阶段(2013/12/6—2014/3/5),‘中茶102’平均含量为0.0446 g·g-1,比第1阶段平均含量0.0370 g·g-1上升了20.54%,‘平阳特早’ 平均含量为0.0519 g·g-1,比第1阶段平均含量0.0374 g·g-1上升了38.77%。当最低气温在2013/12/27、2013/12/28和2014/2/10出现,分别为-0.38℃、 -0.93℃和-0.09℃时,相应地‘中茶102’可溶性糖百分含量在2014/1/5出现1个峰值(4.93%)、在2014/2/15再出现1个峰值(4.95%);而‘平阳特早’在2014/2/15也出现1个峰值(7.95%)。在2014年1月底气温突然升高时,‘中茶102’可溶性糖百分含量在2014/2/5出现低谷(3.42%),‘平阳特早’在2014/2/5也出现低谷(3.23%)。
早春期间(2014/3/6—2014/4/5),‘中茶102’可溶性糖平均含量(4.53%)高于‘平阳特早’(4.19%)(图1)。
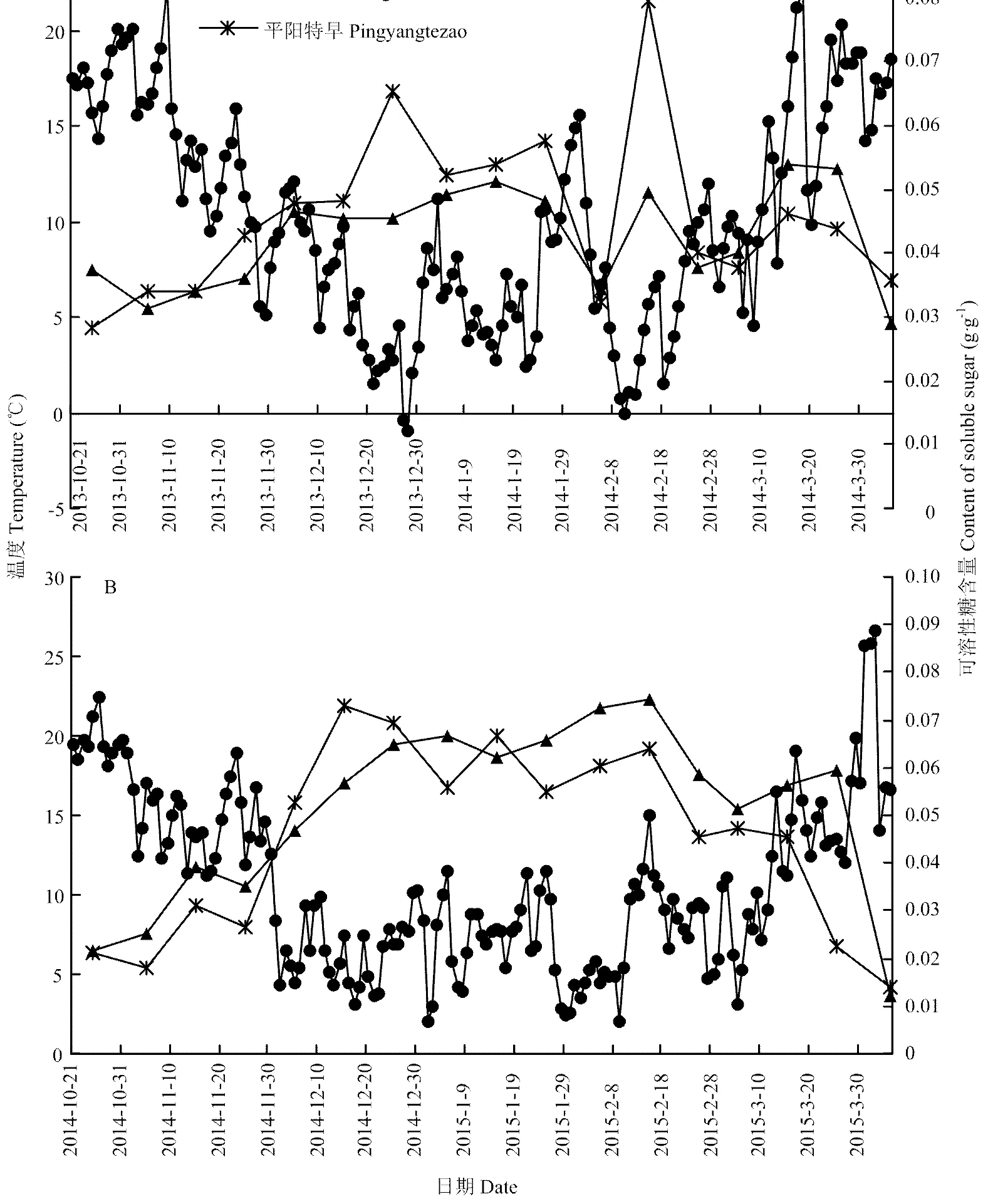
图1 越冬期和早春期2个品种茶鲜叶可溶性糖含量波动及其与温度关系
2014—2015年越冬期间,‘中茶102’可溶性糖平均百分含量(5.30%)高于‘平阳特早’可溶性糖平均百分含量(4.91%)。第1阶段(2014/10/21—2014/12/5),‘中茶102’可溶性糖含量从0.0215 g·g-1(2014/10/25)升至0.0470 g·g-1(2014/12/5),升高了1.2倍;‘平阳特早’的可溶性糖含量从0.0211 g·g-1(2014/10/25)上升至0.0528 g·g-1(2014/12/5),升高了1.5倍;第2阶段(2014/12/6—2015/3/5),‘中茶102’平均含量为0.0637 g·g-1,比第1阶段平均含量0.0337 g·g-1上升了89.13%;‘平阳特早’平均含量为0.0597 g·g-1,比第1阶段平均含量0.0300 g·g-1上升了99.13%。在2015年2月15日温度骤然升高,2种茶树的可溶性糖含量在2月25日分别下降了21.47%(‘中茶102’)和29.13%(‘平阳特早’)。
在早春阶段(2015/3/6—2015/4/5),‘中茶102’可溶性糖的平均百分含量(4.26%)高于‘平阳特早’(2.73%)。
2.2 越冬期和早春期2个品种茶鲜叶脯氨酸含量动态及其与气温关系
2013—2014年越冬期间(图2),‘中茶102’的脯氨酸平均百分含量0.13%高于‘平阳特早’平均百分含量0.07%。在越冬期第1阶段(2013/10/21—2013/12/5),‘中茶102’的脯氨酸含量由0.0034 g·g-1(2013/10/25)逐渐降至0.0015 g·g-1(2013/12/5),降低了55.88%,‘平阳特早’脯氨酸含量由0.0009 g·g-1(2013/10/25)下降至0.0006 g·g-1(2103/12/5),降低了33.33%。
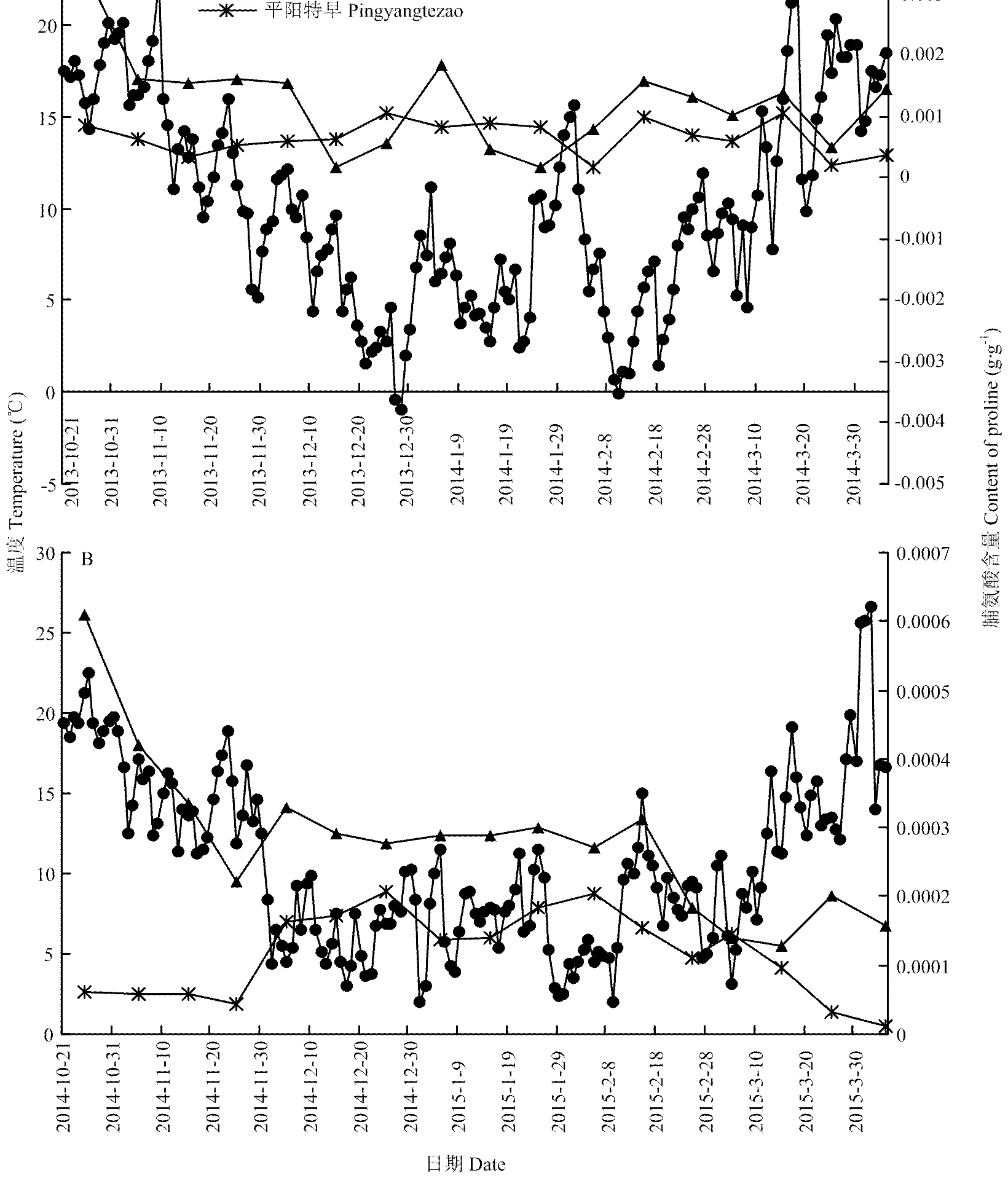
图2 越冬期和早春期2个品种茶叶脯氨酸含量波动及其与温度关系
越冬期第2阶段(2013/12/6—2014/3/5),‘中茶102’脯氨酸平均含量为0.0009 g·g-1,比第1阶段平均含量0.0019 g·g-1下降了52.63%;‘平阳特早’脯氨酸平均含量为0.0007 g·g-1,比第1阶段平均含量0.0006 g·g-1升高了16.67%。当最低气温在2013/12/27、2013/12/28和2014/2/10出现,分别为-0.38℃、-0.93℃和-0.09℃时,‘中茶102’脯氨酸含量在2014/1/5出现1个峰值0.0019 g·g-1;‘平阳特早’脯氨酸含量在2014/2/15出现1个峰值0.0010 g·g-1。在2014年1月底温度突然升高时,2个品种茶树的脯氨酸含量在2014/1/25同时出现低谷,皆为0.0002 g·g-1。
早春期间(2014/3/6—2014/4/5),‘中茶102’的脯氨酸平均百分含量为0.11%,是‘平阳特早’百分含量0.05%的2倍。
2014—2015年越冬期间‘中茶102’的脯氨酸平均百分含量为0.03%,高于‘平阳特早’平均百分含量0.01%。在越冬期第1阶段(2014/10/21—2014/12/5),中茶102脯氨酸百分含量由0.06%(2014/10/25)逐渐降低至0.03%(2014/12/5),下降了50.00%;平阳特早脯氨酸百分含量由0.0060%(2014/10/25)升至0.0162%(2014/12/5),升高了1.70倍。在越冬期第2阶段(2014/12/6—2015/3/5),‘中茶102’脯氨酸平均百分含量0.0262%,比第1阶段脯氨酸平均百分含量0.0383%下降了31.59%;‘平阳特早’脯氨酸平均百分含量0.0162%,比第1阶段平均百分含量0.0077%升高了1.10倍。2015年2月15日温度骤然升高,2种茶树的脯氨酸含量在2014/2/25分别下降了41.07%(‘中茶102’)和31.26%(‘平阳特早’)。早春阶段(2014/3/6—2014/4/5),脯氨酸含量有所下降,‘中茶102’的脯氨酸平均百分含量为0.0162%,比‘平阳特早’(0.0047%)高了3倍。
2.3 越冬期和早春期2个品种茶鲜叶游离氨基酸含量动态及其与气温关系
2013—2014年越冬期间,‘中茶102’游离氨基酸平均百分含量为0.0017 g·g-1,低于‘平阳特早’的游离氨基酸平均百分含量0.0027 g·g-1。在越冬期第1阶段(2013/10/21-2013/12/5),‘中茶102’游离氨基酸含量由0.0029 g·g-1(2013/10/25)降至0.0019 g·g-1(2013/12/5),下降了34.48%,‘平阳特早’由0.0048 g·g-1(2013/10/25)降至0.0027 g·g-1(2013/12/5),下降了43.75%。在越冬期第2阶段(2013/12/6—2014/3/5),‘中茶102’游离氨基酸的平均含量为0.0014 g·g-1,比第1阶段平均含量0.0022 g·g-1降低了36.36%;‘平阳特早’游离氨基酸的平均含量为0.0022 g·g-1,比第1阶段平均含量0.0034 g·g-1下降了35.29%。早春时期(2014/3/6—2014/4/5),2个茶树品种游离氨基酸含量逐渐上升,‘中茶102’的平均含量(0.0019 g·g-1)低于‘平阳特早’平均含量(0.0035 g·g-1)。
2014—2015年越冬期间,‘中茶102’的游离氨基酸平均含量为0.0029 g·g-1,低于‘平阳特早’平均含量0.0035 g·g-1。越冬期第1阶段(2014/10/21—2014/12/5),‘中茶102’的游离氨基酸含量由0.0043 g·g-1(2014/10/25)降至0.0036 g·g-1(2014/12/5),下降了16.28%;‘平阳特早’游离氨基酸平均含量由0.0046 g·g-1(2014/10/25)降至0.0036 g·g-1(2014/12/5),下降了21.74%。在越冬期第2阶段(2014/12/6—2015/3/5),2个茶树品种的游离氨基酸含量较低,‘中茶102’的游离氨基酸平均含量为0.0026 g·g-1,比第1阶段的平均含量0.0034 g·g-1下降了23.53%;‘平阳特早’游离氨基酸的平均含量为0.0029 g·g-1,比第1阶段平均含量0.0044 g·g-1下降了34.09%。在早春阶段(2015/3/6—2015/4/5),游离氨基酸含量开始上升,‘中茶102’游离氨基酸的平均含量(0.0034 g·g-1)低于‘平阳特早’平均含量(0.0041 g·g-1)。
2.4 越冬期和早春期2个品种茶鲜叶3种渗透调节物质含量与气温的数值相关性
以旬平均气温为自变量,以渗透调节物质含量为因变量,检验抗冻物质含量与旬平均气温的相关性,并建立线性回归方程(表1)。在2年试验越冬期间第1阶段和第2阶段:‘中茶102’和‘平阳特早’的可溶性糖含量分别与旬平均气温显著负相关;‘中茶102’和‘平阳特早’的脯氨酸含量与旬平均气温显著负相关;‘中茶102’和‘平阳特早’的游离氨基酸含量与旬平均气温显著正相关(表1)。
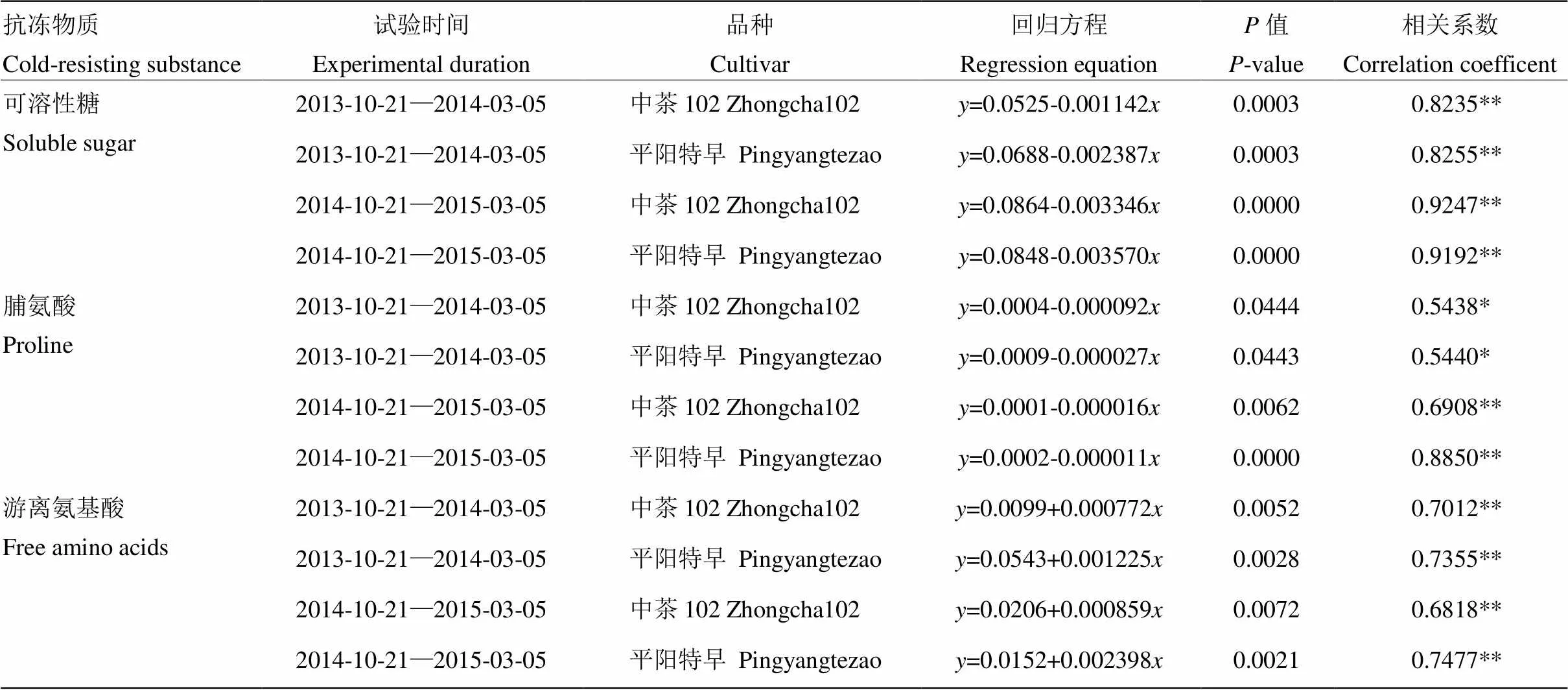
表1 越冬期间2个品种茶鲜叶3种渗透调节物质与气温的相关性
*表示在<0.05水平上差异显著,**表示在<0.01水平上差异显著 * significantly different at<0.05, ** extremely significant different at<0.01
2014/3/6—2014/4/5和2015/3/6—2015/4/5分别为2年试验的早春期间,气温逐渐回升,可溶性糖含量趋向于减少(图1),脯氨酸、游离氨基酸含量则分别趋向于增加(图2和图3)。
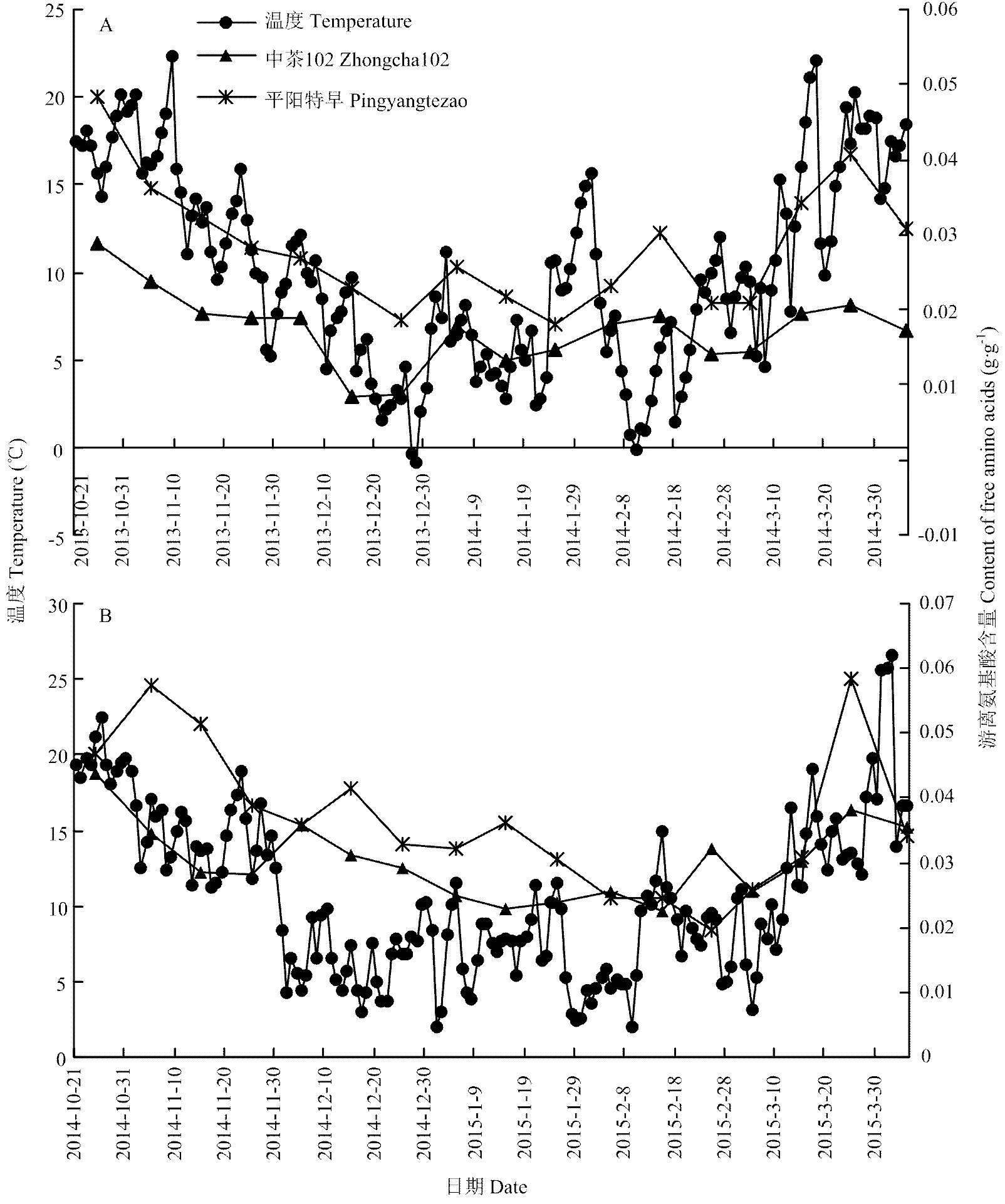
图3 越冬期和早春期2个品种茶叶游离氨基酸含量波动及其与温度关系
3 讨论
3.1 渗透调节——可溶性糖
糖在植物生长中具有重要的调节作用,当植物受到环境胁迫时,可溶性糖含量增加并通过渗透调节增强植物的抵抗力[15-17]。植物受低温胁迫一段时间之后,膜系统发生一系列生理生化变化,通透性增大,膜内可溶性物质外渗,细胞失水,植物组织便会受到损伤以致于死亡[18]。可溶性糖可调节细胞渗透压,增大保水能力,能够供给碳源和底物,诱导其他抗寒生理生化反应以提高植物的抗寒性;还可以保护蛋白质,避免蛋白质遭受低温而发生凝固,以增强植物抗寒性[19-20]。本研究发现:在越冬期第1阶段,可溶性糖含量伴随着温度的下降而上升;在越冬期第2阶段,可溶性糖含量继续升高,最低温出现之后,可溶性糖含量峰值随后也就出现;当温度突然升高时,可溶性糖含量快速下降。说明可溶性糖灵敏地响应温度变化;回归分析表明越冬期可溶性糖含量与气温显著负相关。这些与前人研究结果一致[21-23],也与田野对于‘乌牛早’和‘茂绿’品种的研究结果一致[11]。在早春时期,温度持续上升,‘中茶102’和‘平阳特早’的可溶性糖含量相比越冬期间略有下降,且与此时期的温度无显著相关性,‘中茶102’可溶性糖平均含量高于‘平阳特早’。
3.2 渗透调节——脯氨酸
在越冬期和植物萌芽期,脯氨酸的累积与植物生长密切相关[24-25]。脯氨酸作为植物体内一种渗透调节物质,当植物受到低温胁迫时,游离脯氨酸能够促进蛋白质的水合作用,保护酶结构的稳定,从而保护细胞免受伤害[19]。植物叶片的脯氨酸含量与植物抗寒力有显著相关性,抗寒力强的植物在受到一定范围的低温胁迫之后,游离脯氨酸显著增多[8, 10, 26-27]。但也有报道认为植物体内脯氨酸含量与植物抗寒力无显著相关性[28]。也有研究表明脯氨酸的累积与植物对环境胁迫的响应无显著相关性[29-30]。本研究表明,越冬期间,2个供试茶树品种鲜叶游离脯氨酸含量变化虽没有可溶性糖明显,但2013—2014年、2014—2015年越冬期间2个品种茶树脯氨酸含量皆分别与旬平均气温显著负相关;并且在越冬期第2阶段出现最低温度时,2个品种茶树脯氨酸含量随着都有明显升高,在2014年1月底气温突然升高时2个品种茶树脯氨酸含量也随着出现低谷,比较灵敏地响应温度的变化。认为脯氨酸也是一种重要的渗透调节物质,促进茶树抗冻,与前人研究结果一致[8, 10, 26-27]。在早春期间,‘中茶102’的脯氨酸含量高于‘平阳特早’,两者的脯氨酸含量与气温无显著相关性。
3.3 渗透调节——游离氨基酸
植物抗寒性与可溶性蛋白含量正相关[31],植物遭受低温胁迫后,体内可溶性蛋白遭到破坏而降解,造成氨基酸的累积。在低温处理过程中,稻叶内游离氨基酸大量积累,与可溶性蛋白含量的下降极显著相关[32]。另有研究表明植物在冷训化过程中游离氨基酸含量增加[33]。游离氨基酸含量增加,加大植物C、N源的供给,增强蛋白对于水分子束缚,调节渗透性,也就增强了抗冻性,其抗冻机理尚待深入研究。
但本研究结果表明,越冬期茶树鲜叶的游离氨基酸含量变化趋势与气温一致;随着温度最低值的出现,游离氨基酸含量也相应的达到低谷,温度升高时,游离氨基酸含量也逐渐升高,说明游离氨基酸含量比较敏感地响应温度变化;茶鲜叶游离氨基酸含量与气温极显著正相关。田野等[11]也发现2013年10月20日到翌年4月5日‘乌牛早’和‘茂绿’品种茶鲜叶游离氨基酸含量与气温正相关,但未达到显著水平,可能是品种之间有差异,试验时间也较短。也有研究证实霜冻胁迫会显著影响茶叶游离氨基酸含量,在遭受霜冻胁迫时,游离氨基酸含量下降[34]。早春期气温升高时,游离氨基酸含量也相应地升高,明显高于越冬期含量,‘中茶102’游离氨基酸含量低于‘平阳特早’。
3.4 三类渗透调节物质抗冻性能的初步评判以及‘平阳特早’和‘中茶102’抗冻性比较
优良的渗透调节物质,不仅具有增强茶树免于冻害的生物效应,而且从深秋至严冬乃至早春倒春寒侵袭时能够灵敏地响应低温变化,具有生物统计学意义的相关性。有研究认为,茶树抗冻性与温度密切相关,茶鲜叶可溶性糖、脯氨酸和可溶性蛋白含量与茶树的抗冻性密切相关[35]。本研究发现,可溶性糖、脯氨酸和游离氨基酸在茶树体内具有一定含量,2013—2014、2014—2015年越冬期间这3类物质与气温显著相关,对于温度的响应程度较大。在越冬期第1阶段和第2阶段,‘平阳特早’可溶性糖含量的升高幅度皆大于‘中茶102’的;在越冬期第2阶段‘平阳特早’的脯氨酸含量相比第1阶段显著升高,而‘中茶102’的脯氨酸含量相比第1阶段略有降低;‘平阳特早’游离氨基酸含量在越冬期第1阶段下降幅度高于‘中茶102’,而在第2阶段的平均含量高于中茶102。与‘中茶102’相比,在遭受冬季低温胁迫时,‘平阳特早’3类渗透调节物质对低温的响应性、响应程度分别更灵敏、更大,‘平阳特早’3类渗透调节物质含量稍多。
即便如此,茶树品种抗冻性评价是一个复杂问题,还要结合电导率、酶活等可信的生理生化指标[6, 8, 10]、遗传特性和田间冻害指数[36]等进行综合评价,可溶性糖、脯氨酸和游离氨基酸含量只是其中的几个参考指标。
4 结论
越冬期间可溶性糖、脯氨酸和游离氨基酸皆能敏感地响应空气温度的变化,三者含量分别与旬平均气温显著负相关、显著负相关和显著正相关,被认为是茶树的重要渗透调节物质,可作为抗冻指标。
越冬期间,‘平阳特早’可溶性糖含量、游离氨基酸含量明显大于‘中茶102’,‘平阳特早’可溶性糖、脯氨酸和游离氨基酸对于低温的变幅大于‘中茶102’的,因此,‘平阳特早’对低温响应程度大于‘中茶102’。
References
[1] Hwang J G, Kim Y D. A survey low temperature damage of tea tree at South Korea in 2011., 2012, 14(4): 246-253.
[2] 杨俊虎, 张行才, 王超, 陆小强. 气象因子与春茶及中高档春茶产量的灰色关联分析. 山西农业科学, 2012, 40(1): 53-55.
Yang J H, Zhang H C, Wang C, Lu X Q. Grey connection analysis of spring tea and high-middle grade spring tea yield with meteorological factors., 2012, 40(1): 53-55. (in Chinese)
[3] 朱秀红, 袁洪刚, 郑海涛. 近45年山东茶树冻害气候原因分析. 中国茶叶, 2012(3): 11-13.
Zhu X H, Yuan H G, Zheng H T. Analysis on frozen injury weather of Shandong tea plants., 2012(3): 11-13. (in Chinese)
[4] 吴华玲, 陈栋, 李家贤. 广东茶区“倒春寒”冻害情况的调查与反思. 广东农业科学, 2010(7): 39-41.
Wu H L, Chen D, Li J X. Investigation and reflection of late spring coldness frozen injury from Guangdong tea-producing area., 2010(7): 39-41. (in Chinese)
[5] 黄海涛, 余继忠, 王贤波, 张伟, 周铁锋, 敖存. 不同抗寒性茶树品种秋季新梢的生理特性研究. 浙江农业学报, 2014, 26(4): 925-928.
Huang H T, Yu J Z, Wang X B, zhang w, zhou t f, ao c. Study on physiological characters of new shoot in different cold-resistant tea varieties in autumn., 2014, 26(4): 925-928. (in Chinese)
[6] 李叶云, 庞磊, 陈启文, 周月琴, 江昌俊. 低温胁迫对茶树叶片生理特性的影响. 西北农林科技大学学报(自然科学版), 2012, 40(4): 134-138.
Li Y Y, Pang L, Chen Q W, Zhou Y Q, Jiang C J. Effects of low temperature stress on physiological characteristics of tea leaves (L.)., 2012, 40(4): 134-138. (in Chinese)
[7] 孙海伟, 曹德航, 尚涛, 张虹, 刘静, 谢忠琴, 王相涛. 茶树抗寒育种及转基因研究进展. 山东农业科学, 2013, 45(6): 119-122, 129.
Sun H W, Cao D H, Shang T, Zhang H, Liu J, Xie Z Q, Wang X T. Advances in research of cold-resistant breeding and transgene of tea plant., 2013, 45(6): 119-122, 129. (in Chinese)
[8] 张婷, 车凤斌, 潘俨, 胡柏文, 许娟, 李萍. 低温胁迫对核桃枝条几个抗寒生理指标的影响. 新疆农业科学, 2011, 48(8): 1428-1433.
Zhang T, Che F B, Pan Y, Hu B W, Xu J, Li P. Influence of low temperature stress on several cold resistance indexes of the primary branches of walnuts., 2011, 48(8): 1428-1433. (in Chinese)
[9] 李叶云, 舒锡婷, 周月琴, 江昌俊. 自然越冬过程中3个茶树品种的生理特性变化及抗寒性评价. 植物资源与环境学报, 2014, 23(3): 52-58.
Li Y Y, Shu X T, Zhou Y Q, Jiang C J. Change in physiological characteristics and cold resistance evaluation of three cultivars ofduring natural overwintering period., 2014, 23(3): 52-58. (in Chinese)
[10] 裴文, 李鹏, 裴海潮, 刘增喜, 杨秋生. 低温条件下9种木兰科植物抗寒性研究. 河南农业科学, 2014, 43(4): 101-105.
Pei W, Li P, Pei H C, Liu Z X, Yang Q S. Comparative analysis of physiological index related to cold resistance of nine magnoliaceae plants under natural and artificial low temperature conditions., 2014, 43(4): 101-105. (in Chinese)
[11] 田野, 王梦馨, 王金和, 韩宝瑜. 茶鲜叶可溶性糖和氨基酸含量与低温的相关性. 茶叶科学, 2015, 35(6): 567-573.
Tian Y, Wang M X, Wang J H, Han B Y. Correlation of low temperature with soluble sugar and amino acid content in fresh tea leaves., 2015, 35(6): 567-573. (in Chinese)
[12] 林平, 赵东. 平阳特早茶研究. 茶叶, 1998, 24(4): 200-204.
Lin P, Zhao D. A study of Pingyang Tezaocha (L.)., 1998, 24(4): 200-204. (in Chinese)
[13] 杨亚军, 杨素娟, 曾建明, 孙涛, 王玉书. 茶树新品种中茶102选育研究报告. 中国茶叶, 2004(6): 14-15.
Yang Y J, Yang S J, Zeng J M, Sun T, Wang Y S. A study of Zhongcha 102 (L.)., 2004(6): 14-15. (in Chinese)
[14] 陈宗懋. 中国茶叶大辞典. 北京: 中国轻工业出版社, 2000: 117.
Chen Z M.. Beijing: China Light Industry Press, 2000: 117. (in Chinese)
[15] LASTDRAGER J, HANSON J, SMEEKENS S. Sugar signals and the control of plant growth and development., 2014,65(3): 799-807.
[16] QAYYUM A. Water stress causes differential effects on germination indices, total soluble sugar and proline content in wheat (L.) genotypes., 2011, 10(64): 14038-14045.
[17] MA Y Y, ZHANG Y L, LU J, SHAO H B. Roles of plant soluble sugars and their responses to plant cold stress., 2009, 8(10): 2004-2010.
[18] 骆耀平. 茶树冻害的发生及防御. 中国茶叶, 2008(1): 30-31.
Luo Y P. Introduction to frozen injury of tea plants and their prevention measures., 2008(1): 30-31. (in Chinese)
[18] 邱乾栋, 吕晓贞, 臧德奎, 张雷, 张立才, 杜淑辉. 植物抗寒生理研究进展. 山东农业科学, 2009, 8: 53-57.
Qiu Q D, Lv X Z, Zang D K, Zhang L, Zhang L C, Du S H. Research progress on plant physiology of cold resistance., 2009, 8: 53-57. (in Chinese)
[20] Yue C, Cao H L, Wang L. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season., 2015, 88(6): 591-608.
[21] 朱政, 蒋家月, 江昌俊, 李雯. 低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响. 安徽农业大学学报, 2011, 38(1): 24-26.
Zhu Z, Jiang J Y, Jiang C J, Li W. Effect of low temperature stess on SOD activity, soluble protein content and soluble sugar content inleaves., 2011, 38(1): 24-26. (in Chinese)
[22] 赵明明, 周余华, 彭方仁, 郝明灼, 梁有旺, 任莺, 华宏. 低温胁迫下冬青叶片细胞内Ca2+水平及可溶性糖含量的变化. 南京林业大学学报(自然科学版), 2013, 37(5): 1-5.
Zhao M M, Zhou Y H, Peng F R, Hao M Z, Liang Y W, Ren Y, Hua H. Changes of Ca2+level and soluble sugar content in cells ofL. leaflets under low temperature stress., 2013, 37(5): 1-5. (in Chinese)
[23] 姚远, 闵义, 胡新文, 李开绵, 郭建春. 低温胁迫对木薯幼苗叶片转化酶及可溶性糖含量的影响. 热带作物学报, 2010, 31(4): 556-560.
Yao Y, Min Y, Hu X W, Li K M, Guo J C. Effect of low temperature stress on the activity of invertase and soluble sugar content in leaves of Cassava seedlings., 2010, 31(4): 556-560. (in Chinese)
[24] Sagisaka S, Araki T. Amino acid pools in perennial plants at the wintering stage and at the beginning of growth., 1983, 24(3): 479-494.
[25] Delauney A J, Verma D P S. Proline biosynthesis and osmoregulation in plants., 1993, 4(2): 215-223.
[26] 罗丹, 张喜春, 田硕. 低温胁迫对番茄幼苗脯氨酸积累及其代谢关键酶活性的影响. 中国农学通报, 2013, 29(16): 90-95.
Luo D, Zhang X C, Tian S. Effect of low temperature stress on proline accumulation and the activities of the key enzymes involved in the proline metabolism in leaves of tomato seedling., 2013, 29(16): 90-95. (in Chinese)
[27] 陈璇, 李金耀, 马纪, 张富春. 低温胁迫对春小麦和冬小麦叶片游离脯氨酸含量变化的影响. 新疆农业科学, 2007, 44(5): 553-556.
Chen X, Li J Y, Ma J, Zhang F C. Effect of low temperature stress on change of free proline content in the leaves of spring and winter wheat., 2007, 44(5): 553-556. (in Chinese)
[28] 杨亚军, 郑雷英, 王新超. 冷驯化和ABA对茶树抗寒力及其体内脯氨酸含量的影响. 茶叶科学, 2004, 24(3): 177-182.
Yang Y J, Zheng L Y, Wang X C. Effect of cold acclimation and ABA on cold hardiness, contents of proline in tea plants., 2004, 24(3): 177-182. (in Chinese)
[29] BORGO L, MARUR C J, Vieira L G E. Effects of high proline accumulation on chloroplast and mitochondrial ultrastructure and on osmotic adjustment in tobacco plants., 2015, 37(2): 153-175.
[30] LÜ B S, MA H Y, LI X W, Wei, Li X, LÜ H Y, YANG H Y, JIANG C J, LIANG Z W. Proline accumulation is not correlated with saline-alkaline stress tolerance in rice seedlings., 2015, 107(1): 51-60.
[31] 梁锁兴, 孟庆仙, 石美娟, 罗钢铁, 王贵珠. 平欧榛枝条可溶性蛋白及可溶性糖含量与抗寒性关系的研究. 中国农学通报, 2015, 31(13): 14-18.
Liang S X, Meng Q X, Shi M J, Luo G T, Wang G Z. Reasearch on relationship between cold-resistance and soluble protein and sugar of×branches., 2015, 31(13): 14-18. (in Chinese)
[32] 龚明, 刘友良, 朱培仁. 低温下稻苗叶片中蛋白质及游离氨基酸的变化. 植物生理学通讯, 1989(4): 18-22.
Gong M, Liu Y L, Zhu P R. Changes of proteins and free amino acids in leaves of rice seedlings during chilling stress., 1989(4): 18-22. (in Chinese)
[33] KOVACS Z, Simon-Sarkadi L, Sovany C, Kirsch K, Galiba G, Kocsy G. Differential effects of cold acclimation and abscisic acid on free amino acid composition in wheat., 2011, 180(1): 61-68.
[34] 郭湘, 唐茜, 许燕, 陈玖琳, 王自琴. 早春霜冻对不同茶树品种芽叶的生化成分及制茶品质的影响. 云南大学学报(自然科学版), 2015(6): 930-938.
Guo X, Tang Q, Xu Y, Chen J L, Wang Z Q. Effect of frost in early spring on biochemical composition and sensory quality of new shoots in different tea cultivars., 2015(6): 930-938. (in Chinese)
[35] 林郑和, 钟秋生, 陈常颂. 茶树抗性育种研究进展. 福建茶叶, 2015(4): 2-4.
Lin Z H, Zhong Q S, Chen C S. Research progress on resistance breeding of tea plant., 2015(4): 2-4. (in Chinese)
[36] 韩宝瑜, 章正和, 房国宾, 黄安智. 茶树益微增强茶树抗寒和复壮效能的试验研究. 茶叶, 1996, 12(1): 18-19.
Han B Y, Zhang Z H, Fang G B, Huang A Z. Study on enhancing cold-resistant of tea plant by tea beneficial microbes., 1996, 12(1): 18-19. (in Chinese)
(责任编辑 李莉)
Evaluation on Correlations of Three Kinds of Osmoregulation Substances in Tea Fresh Leaves with Low Temperature during Winter and Spring Respectively and Their Difference among Cultivars
BO Xiao-pei1, WANG Meng-xin1, CUI Lin1, WANG Jin-he2, HAN Bao-yu1
(1Zhejiang Provincial Key Laboratory of Biometrology and Inspection & Quarantine, China Jiliang University, Hangzhou 310018;2Jiangsu Yinchunbiya Tea Co. Ltd, Danyang 212345, Jiangsu)
【Objective】 The content dynamics of three types of osmoregulation substances in fresh leaves of two cultivars of tea plants and their correlations with air temperatures during winter and early spring were investigated, to evaluate the differences in their responsiveness to the low temperature between the two tea cultivars. 【Method】 Two early bud tea cultivars, ‘Pingyangtezao’ and ‘Zhongcha 102’, were chosen, and air temperatures around their tea shoots were recorded every day from Oct. 21 to April 5 for two consecutive years. The temperature of the dry and wet bulb thermometer at 8:00, 14:00 and 20:00 was daily recorded, and the average of the three dry bulb thermometers set in the experimental tea plantations three times per day standard for the temperature of tea shoot habitat. During these two testing periods adult tea leaves of each cultivar were sampled on the fifth, fifteenth and twenty-fifth days of each month. The soluble sugar contents in the tea fresh leaves were measured by anthrone colorimetric method, the remaining clean fresh leaves were processed into the steamed green tea samples, whereas the contents of proline and free amino acids were determined by SDE-HPLC method. 【Result】The test phase was divided into two stages, the overwintering period (2013/10/21-2014/3/52014/10/21-2015/3/5) and early spring period (2014/3/6-2014/4/52015/3/6-2015/4/5) according to the developmental threshold temperature and the dormancy temperature of the tea plant. Accompanied with initial gradual drops and then maintaining at low air temperatures during the two winters (2013/10/21-2014/3/5, 2014/10/21-2015/3/5), the soluble sugar contents in fresh tea leaves of the two cultivars increased continuingly, negatively correlated with the 10-day average air temperatures. The proline contents increased first and then kept at a high level, also negatively correlated with the 10-day average air temperatures. The free amino acid contents dropped first and then stayed at a low level, which was positively correlated with the 10-day average air temperatures. When the temperature appeared the lowest value, the content of soluble sugars and the content of proline also correspondingly appeared the highest values subsequently, then the content of free amino acids also correspondingly appeared the lowest value. When the temperature was the highest value, the content of soluble sugar and the content of proline also correspondingly appeared the lowest value later, the content of the free amino acids also correspondingly appeared the highest value soon afterwards. During the two spring seasons (2014/3/6-2014/4/5, 2015/3/6-2015/4/5), as air temperatures increased gradually, the soluble sugar contents decreased, while the proline and the free amino acid contents appeared to increase.The frost-resistance of tea plant closely related with air temperature, and soluble sugar, the contents of proline and free amino acids in tea leaves closely related with the frost-resistance of tea plants. During the two winter seasons, compared with ‘Zhongcha 102’, when suffered from the low temperature stress in winter, the content of each of three kinds of osmoregulation substances of ‘pingyangtezao’ more sensitively responsed to low temperature than ‘Zhongcha 102’, and the contents of the three kinds of substances in cultivar ‘pingyangtezao’ were slightly high. 【Conclusion】 Tea plants are vulnerable to frost, freeze and the late spring coldness from late autumn to Qingming Festival. As important cold-resisting indices of tea plants, the contents of soluble sugars, proline and the free amino acids varied sensitively in responses to the low temperatures, and showed significant correlations with the 10-day average air temperatures during the winter and early spring seasons. The responsiveness of cultivar ‘Pingyangtezao’ to the low temperatures was greater than that of cultivar ‘Zhongcha 102’.
low temperature;L.; cold damage; soluble sugar; proline; free amino acids
2016-03-14;接受日期:2016-05-23
国家“863”计划(2012AA10A508)
薄晓培,E-mail:514451641@qq.com。通信作者韩宝瑜,Tel:0571-86835706;E-mail:han-insect@263.net
猜你喜欢
中老年保健(2022年3期)2022-11-21
实用手外科杂志(2022年2期)2022-08-31
Plasma Science and Technology(2022年1期)2022-02-15
科学导报·学术(2020年29期)2020-10-21
边疆文学(2020年5期)2020-05-22
西夏研究(2020年1期)2020-04-01
科学与财富(2019年15期)2019-10-21
中国三峡(2017年9期)2017-12-19
湖南林业科技(2017年1期)2017-02-06
现代检验医学杂志(2015年4期)2015-02-06
