
猪瘟病毒石门株在ST细胞连续传代后E0基因变异分析
2016-07-16 02:30孙永科胡超英林明星陆欣然杨玉艾
动物医学进展 2016年6期
关键词:变异
孙永科,胡超英,林明星,陆欣然,杨玉艾
(云南农业大学动物科学技术学院,云南昆明 650201)
猪瘟病毒石门株在ST细胞连续传代后E0基因变异分析
孙永科,胡超英,林明星,陆欣然,杨玉艾*
(云南农业大学动物科学技术学院,云南昆明 650201)
摘要:为了解猪瘟病毒(CSFV)石门(Shimen)株连续传代后变异情况,根据已发表的CSFV E0基因序列(AF092448.2),设计合成引物,以在ST细胞中连续传代培养的CSFV石门株细胞毒总RNA为模板,每5代通过RT-PCR方法扩增病毒的E0基因,测定其核苷酸序列和氨基酸序列,用DNA Star软件分析比较原代、5代、10代、15代、20代、25代接毒细胞中CSFV E0基因的变异性。结果显示,接种病毒第10代的病毒E0基因在630核苷酸位点出现变异,第15代病毒在632核苷酸位点出现变异。本研究通过分子生物学方法发现CSFV Shimen株在ST细胞连续传代过程中E0基因能保持遗传的稳定性。
关键词:猪瘟病毒;连续传代;ST细胞;E0基因;变异
猪瘟(Classical swine fever,CSF)是由猪瘟病毒(Classical swine fever virus,CSFV)引起的猪的一种急性、热性和高度接触性传染病,具有流行广泛、发病率和病死率高的特点,我国将其列为一类动物疫病[1]。近年来,该病出现许多新特点,以非典型温和型为主,给猪病防控带来许多新的问题。CSFV属于黄病毒科(Flaviviridae)瘟病毒属(Pestivirus),是具有囊膜的单股正链RNA病毒,全基因组长约12.3 kb[2-3],包括4个结构蛋白(Erns/E0、C、E1和E2)和8个非结构蛋白[4],其中E0和E2能够刺激机体产生中和抗体,可诱导免疫猪产生对致死量CSFV的保护性免疫,同时E0和E2也是病毒吸附进入敏感细胞的必需蛋白[5]。E0蛋白是一种多功能性蛋白,由于编码E0的核酸序列保守程度高,分子质量约为 44 ku~50 ku,其同源二聚体分子质量为97 ku,无疏水的膜锚定结构,能直接从被感染的细胞内分泌到细胞外,为可分泌性蛋白并可诱导机体产生中和抗体抵抗致死剂量CSFV的攻击,因而E0也可作为防治猪瘟的一种靶蛋白[6]。E0蛋白具有神经细胞毒性和抗凝集活性,CSFV感染动物后所产生的E0蛋白在致病过程中起重要作用[7]。还参与病毒粒子黏附和侵入宿主细胞的过程,用E0中和单抗可有效阻断CSFV对易感动物细胞的感染[8]。根据E0蛋白对淋巴细胞有直接的毒性效应,能引起淋巴细胞的凋亡,同时猪瘟病毒感染是以白细胞减少和免疫抑制为特征,因此认为E0很可能与CSFV的致病性有关[9]。E0还具有RNase活性[10],E0的这种RNase活性在病毒复制及病毒感染的致病机制中的作用一直备受关注。E0蛋白以同源二聚体形式存在于病毒粒子中,含有227个氨基酸,其中191-227 氨基酸具有猪瘟病毒特异性[11]。本试验通过对CSFV E0基因进行扩增、核苷酸序列及氨基酸序列分析,用分子生物学的手段发现E0遗传变异特点,从而对研究该区域蛋白的结构与功能、鉴别诊断技术和猪瘟基因工程疫苗研究奠定基础,对进一步了解CSFV的分子流行病学、遗传变异及做好猪瘟的防控工作等方面的研究奠定基础。
1材料与方法
1.1材料
1.1.1细胞和病毒ST细胞、猪瘟病毒石门株由云南农业大学农业生物多样性应用技术国家工程研究中心保存。
1.1.2引物参考GenBank中已公布的 CSFV石门株全基因组序列(AF092448.2)中的E0基因设计特异性引物,预计扩增长度为699 bp,引物由上海生工生物工程技术服务有限公司合成。引物E0 U 5′-GGGCGGTAATAGCAATTATGT-3′,E0 L 5′-GGGGTGCAGTTGTTAGTATAC-3′,扩增片段长度699 bp。
1.1.3主要试剂DMEM培养基、血清、胰酶为Gibico公司产品;Trizol Reagent为Invitrogen公司产品;DEPC水、异丙醇、氯仿、无水乙醇、Random引物、RNA抑制剂、dNTP Mixture、AMV反转录酶、exTaq聚合酶均为宝生物工程(大连)有限公司产品;DNA Marker DL 2 000为上海生工生物工程技术服务有限公司产品。
1.2方法
1.2.1ST细胞复苏与培养由液氮罐中迅速取出冻存的ST细胞(1.5 mL冻存管),立即放入37℃水浴中,融化后缓慢放入盛有9 mL DMEM液的无菌离心管中,1 500 r/min室温离心5 min,弃上清,将沉淀放入盛有10 mL含100 mL/L犊牛血清及10 mL/L双抗的DMEM培养液的无菌培养瓶中,37℃培养36 h~48 h,倒置显微镜下观察细胞生长情况。待ST细胞长成单层后进行细胞传代,倾去培养液,用无菌PBS洗2次,加入2.5 g/L胰酶-EDTA 1 mL,轻摇数次,平放消化数分钟,边消化边在显微镜下观察,直至细胞分散、变圆,即将脱落时,弃胰酶,加入含有犊牛血清100 mL/L的DMEM培养液,37℃继续培养,备用。
1.2.2病毒培养取生长良好的ST单层细胞,弃去培养液,用PBS液清洗2次,按照1×103TCID50接种CSFV Shimen株,在37℃培养箱内吸附1 h,其间轻摇2次,之后加入含终浓度20 mL/L~30 mL/L血清的DMEM,37℃培养,逐日观察,5 d为一个传代周期。传代前细胞培养物反复冻融3次,同样方法进行传代。
1.2.3病毒总RNA的提取取细胞液400 μL于1.5 mL EP管中,加入600 μL TrizoL混匀,冰浴5 min;加200 μL氯仿,振荡15 s~30 s,冰浴5 min,12 000 r/min、4℃离心15 min;小心吸取上清于1.5 mL EP管,加入等体积的异丙醇,缓慢颠倒数次,冰浴5 min,12 000 r/min、4℃离心10 min;弃上清,加750 mL/L乙醇1 mL,9 000 r/min、4℃离心5 min;弃上清,自然风干后,DEPC水溶解。所有的EP管及枪头均经1 mL/L的DEPC水处理去除RNA酶。
1.2.4病毒cDNA的合成参照AMV反转录酶的使用说明,进行两步法RT-PCR获得病毒的cDNA1.8E0基因片段的PCR扩增,在一个体系中分别加入超纯水、ExTaqbuffer、dNTP、TaqDNA 聚合酶、反转录反应产物和引物。PCR反应条件为:95℃ 5 min;94℃ 1 min,60℃ 45 s,72℃ 1 min,5个循环;94℃ 1min,53℃ 45 s,72℃ 1 min,30个循环;最后在72℃延伸10 min。扩增结束后取上述产物5 μL混合适量的buffer和花青素在7 g/L 琼脂糖凝胶电泳检测。
1.2.5核酸序列测定及分析将所得的PCR反应产物送往上海生工生物工程技术服务有限公司测序。利用DNA Star软件分析比较多次细胞传代的猪瘟病毒全基因组的核苷酸序列和氨基酸序列的同源性、变异情况,并绘制系统发育进化树。
2结果
2.1E0基因的克隆
扩增产物经7 g/L琼脂糖凝胶电泳,在约800 bp处出现扩增条带,其大小与预期的相符。
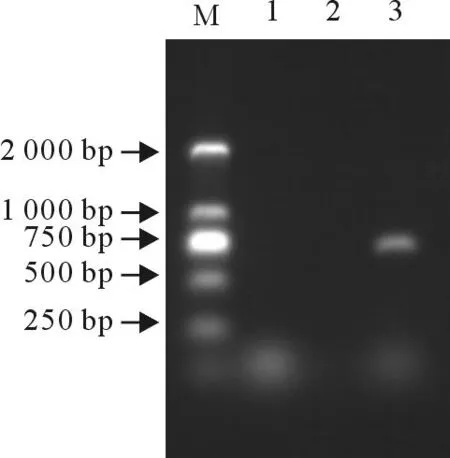
M.DNA 标准 DL 2 000;1.阴性细胞对照;2.水对照;3.E0基因
M.DNA Marker DL 2 000; 1.Negative cell control; 2.H2O control; 3.CSFV E0 gene
图1CSFV E0基因PCR扩增
Fig.1PCR amplification of CSFV E0 gene
2.2基因核苷酸序列比较
克隆测序后,用DNA Star 软件将猪瘟毒Shimen株原代与5代、10代、15代、20代、25代E0基因的核苷酸序列进行分析比较,结果如图2所示,从图2可以看出10代第630核苷酸位点开始出现变异,15代第632核苷酸位点开始出现变异,图3显示核苷酸序列的同源性为99.6%~100%,将核苷酸同源性分析结果绘制系统发育进化树(图4)。
2.3氨基酸序列分析
利用DNA Star软件根据核苷酸序列推导氨基酸序列,可见发生核苷酸变异的位点氨基酸也发生改变(图5),氨基酸同源性为99.1%~100%(图6)。
3讨论
猪瘟病毒是单股正链RNA病毒,由于RNA聚合酶的校正活性,RNA病毒较易发生变异[12-13]。已有研究证实RNA病毒基因组在群落传代过程中能够产生广泛的分子和抗原性改变[14],通过进化树分析也已经证明猪瘟病毒野毒株之间存在着基因组的变异[15]。目前我国防控猪瘟的主要措施是疫苗免疫,猪瘟细胞苗则是其中一种,但在现行猪瘟细胞苗的使用过程中,往往出现免疫失败诸多问题,究其原因有报道认为猪瘟病毒的抗原变异可能是导致该病毒逃逸免疫保护的主要原因。E0蛋白是猪瘟病毒的主要保护性抗原,同时也是一个相当保守的蛋白,对编码该蛋白的基因序列进行变异分析,对研究CSFV抗原的变异性和多样性有重要意义,同时有利于从分子水平上揭示近年来猪瘟病毒毒力变异及非典型猪瘟广泛传播的原因。
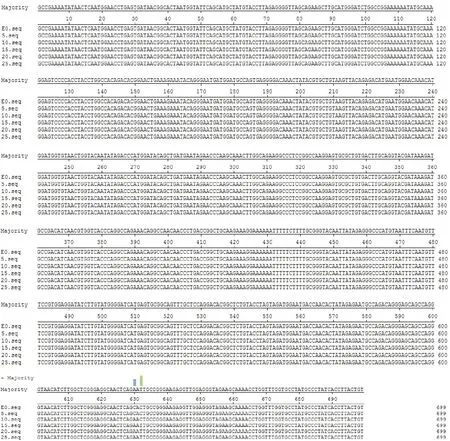
图2 不同代次间E0基因核苷酸序列比较
图3 不同代次间E0基因核苷酸序列同源性比较
图4 不同代次间系统进化树
本研究采用PCR技术,扩增了在ST细胞中连续传代培养的猪瘟病毒石门株细胞毒E0基因,运用DNA Star软件分析比较多次细胞传代后猪瘟病毒E0基因的核苷酸序列和氨基酸序列的变异情况,从结果我们可以看出仅2个位点发生变异,分别为630和632核苷酸位点,同源性高达99.6%~100%。从推导的氨基酸序列来看发生核苷酸变异的位点氨基酸也发生改变,虽然只有2个氨基酸发生变异,但都是亲水性氨基酸残基,运用DNA Star软件进行蛋白分析发现,发生变异后该位置的α螺旋和β折叠均发生变化,氨基酸残基在特定结构的位置由内部转到外部,潜在的蛋白质抗原决定簇也下降。E0基因作为病毒诱导机体产生中和抗体的一个主要保护性抗原和防治猪瘟的一种靶蛋白,具有神经细胞毒性和抗凝集活性,与CSFV的致病性和病毒的宿主嗜性有关,而且E0的RNase活性在病毒复制及病毒感染的致病机制有关联。因此,关于猪瘟病毒石门珠在ST细胞中连续传代后E0基因的变化应进行深入的研究。

图5 不同代次间E0基因氨基酸序列比较
图6 不同代次间E0基因氨基酸序列同源性比较
参考文献:
[1]李学伍,张改平,郭启祥.猪瘟病毒生物学特性研究概述[J].河南农业科学,2007(10):11-15.
[2]冶贵生,张彦明,徐浩.载体介导的shRNA抑制猪瘟病毒在PK-15细胞中增殖特性的研究[J].畜牧兽医学报,2007,38(5):500-505.
[3]Li L N,Maritza B.A DNA vaccine expressing the E2 protein of classical swine fever virus elicits T cell responses that can prime for rapid antibody production and confer total protection upon viral challenge[J].Vaccine,2005,23:3741-3752.
[4]李国新,李娜,仇华吉,等.猪瘟病毒石门株基因组全长cDNA的克隆与序列分析[J].中国预防兽医学报,2006,3(28):275-278.
[5]Riedel C,Lamp B,Heimann M,et al.Characterization of essential domains and plasticity of the classical swine fever virus core protein[J].J Virol,2010,84(21):11523-11531.
[6]Frey C F,Bauhofer O,Ruggli N,et al.Classical swine fever virus replicon particles lacking the Erns gene:a potential marker vaccine for intradermal application[J].Vet Res,2006,37(5):655-670.
[7]孙永科,杨玉艾,王养会,等.表达猪瘟病毒石门株E0抗原的重组腺病毒构建及免疫性的研究[J].畜牧兽医学报,2007,38(5):482-487.
[8]刘坤,兰邹然,姜平,等.猪瘟病毒分子生物学及检测技术研究进展[J].动物医学进展,2012,33(10):99-104.
[9]夏燕华,赵天生,张楚瑜.猪瘟病毒全长突变型cDNA克隆的构建及其在细胞水平的恢复 [J].南昌大学学报(理科版),2009,4(32):374-379.
[10]Lin M,McRae H,Dan H,et al.High-resolution epitope mapping for monoclonal antibodies to the structural protein E0 of classical swine fever virus using peptide array and random peptide phage display approaches[J].J Gen Virol,2010,91(Pt 12):2928-2940.
[11]Ahuja A,Sen A,Yogisharadhya R,et al.Prokaryotic expression and purification of highly soluble partial glycoprotein erns of Indian strain of classical swine fever virus[J].Indian J Virol,2012,23(3):397-401.
[12]Kumar R,Barman N N,Khatoon E,et al.Molecular characterization of E2 glycoprotein of classical swine fever virus:adaptation and propagation in porcine kidney cells[J].InVitroCell Dev Biol Anim,2015,51(5):441-446.
[13]曾咏芳,曹佳媛,杨可妍,等.猪瘟病毒广西流行毒株E0基因的克隆与分析[J].动物医学进展,2015,36(1):31-35.
[14]郝飞,汤德元,曾智勇,等.我国猪瘟病毒基因流行变异研究[J].猪病防控,2013(3):42-44.
[15]吴旭锦.咸阳地区猪瘟病毒E2基因扩增与序列分析[J].动物医学进展,2014,35(12):49-53.
Variation Analysis of E0 Gene of Classical Swine Fever Virus Shimen Strain in Continuously Passed ST Cells
SUN Yong-ke,HU Chao-ying,LIN Ming-xing,LU Xing-ran,YANG Yu-ai
(YunnanAgricultureUniversity,Kunming,Yunnan,650201,China)
Abstract:In order to study genetic variation of classical swine fever virus Shimen strains,the primers of CSFV E0 gene of Shimen strain were designed and synthesized according to the published sequence(AF092448.2). The total RNA of classical swine fever virus Shimen strain continuouly passed in ST cells used as a template,and E0 gene was amplified by RT-PCR every five passages of cultured swine fever virus.E0 nucleotide and amino acid sequences were determined.Variabilities of primary,5,10,15,20 and 25 passage viruses were analyzed by using DNA Star software.The results showed that variation appeared in 630 nucleotide point of 10 passage virus,and 632 nucleotide point of 15 passage virus.By molecular biological methods,this research revealed that the E0 genes of CSFV Shimen strain were genetically stable after passages in the ST cells.
Key words:Classical swine fever virus; continuous passage; ST cell; E0 gene; variation
收稿日期:2015-11-19
基金项目:国家自然科学基金项目(31560704)
作者简介:孙永科(1977-),男,江苏盐城人,博士,主要从事预防兽医学研究。*通讯作者
中图分类号:S852.651
文献标识码:A
文章编号:1007-5038(2016)06-0013-05
猜你喜欢
传染病信息(2022年6期)2023-01-12
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
水利规划与设计(2020年1期)2020-05-25
摄影之友(影像视觉)(2019年3期)2019-03-30
现代计算机(2018年7期)2018-04-24
中学生理科应试(2017年2期)2017-04-01
试题与研究·高考理综生物(2016年4期)2017-03-28
百科知识(2015年18期)2015-09-10
小星星·阅读100分(高年级)(2015年4期)2015-05-26
