
红树植物角果木属的分子系统发育研究
2016-06-15 09:08:55朱春超谈凤笑刘莹强寅梦施苏华黄椰林中山大学生命科学学院广东省植物资源重点实验室广东广州5075华南农业大学资源环境学院广东广州5064
海洋通报 2016年2期
朱春超,谈凤笑,刘莹,强寅梦,施苏华,黄椰林(.中山大学 生命科学学院 广东省植物资源重点实验室,广东 广州 5075;.华南农业大学 资源环境学院,广东 广州 5064)
红树植物角果木属的分子系统发育研究
朱春超1,谈凤笑2,刘莹1,强寅梦1,施苏华1,黄椰林1
(1.中山大学生命科学学院广东省植物资源重点实验室,广东广州510275;2.华南农业大学资源环境学院,广东广州510642)
摘要:运用PCR产物直接测序法测定了红树科(Rhizophoraceae)角果木属(Ceriops)5个物种24个个体加上外类群木榄共125个序列,包括rpl20-rps12,atpB-rbcL,trnL intron,trnL-trnF,trnT(UGU)-trnL(UAA)共5段叶绿体非编码区,序列总长度为2716 bp。基于以上合并序列,采用最大简约法、贝叶斯法和邻接法所构建的系统发育树呈现出高度一致的系统发育关系,结果显示:角果木属包含两个大的分支:(1)角果木C. tagal和澳洲角果木C. australis构成一个分支;(2)十雄角果木复合体内的3个物种(十雄角果木C. decandra、齐氏角果木C. zippeliana和拟十雄角果木C. pseudodecandra)组成另一分支,其中齐氏角果木和拟十雄角果木为姊妹种,该分支3个物种相互之间的遗传距离均显著大于传统物种角果木和澳洲角果木之间的遗传距离,并且每个物种的单系性在系统树上均得到强烈支持。研究结果支持了Sheue在最近研究中关于该属物种地位的处理,进一步探讨了角果木属系统发育关系与该属生物地理分布模式之间的相关性。
关键词:红树植物;角果木属;分子系统发育
红树科(Rhizophoraceae)角果木属(Ceriops)是热带亚热带海岸红树林植物类群中的重要树种,是以印度-西太平洋地区为分布中心的代表性真红树植物。该属植物叶片呈光滑偏黄绿色,故被称为“yellow mangrove”(Tomlinson,1986);传统上该属分为3种:十雄角果木C. decandra、角果木C. tagal、澳洲角果木C. australis,其中以角果木和十雄角果木分布最广,其分布范围从非洲东岸、马达加斯加向东经热带亚洲和澳大利亚直至美拉尼西亚(Tomlinson,1986);澳洲角果木分布范围很窄,仅限于澳大利亚、新几内亚及其邻近岛屿。中国仅有角果木1种,其南部沿海是该种世界分布的北缘(Duke,1992)。最新的调查表明其分布范围由原来的广西、海南、广东直至台湾南部缩减为仅有的海南、广东两省(陈小勇等,2000;梁士楚,2000;王文卿等,2007)。近年来,随着印度西太平洋地区海岸红树植物实地调查的不断深入和植物材料的重新整理和审视,角果木属的系统发育研究有了新的进展,有研究显示传统上所认为的十雄角果木可能为一个物种复合体(species complex),能进一步根据形态特征划分为3个物种(十雄角果木C. decandra,齐氏角果木C. zippeliana,拟十雄角果木C. pseudodecandra)(Sheue et al,2009a;Sheue et al,2010)。但这种物种划分尚未得到进一步的验证,各物种之间的关系也存在矛盾和疑问(Tsai et al,2012)。此外,该属植物物种之间形态差异相对较小,各种性状往往相互交叉,种间界限难以区分,分类和鉴定非常困难。因此利用新的证据进一步的开展角果木属的系统发育研究无疑是该属研究的重要方向。
随着分子生物学技术的发展,合理的分子标记被认为能较为客观地提供植物类群中的系统发育信息并解决形态特征上的趋同进化干扰。其中叶绿体基因组中的内含子和基因间隔区因为均属非编码序列,进化速率快,常被用于探讨属间或属下类群的系统发育关系。常用的叶绿体非编码区域包括rpl20-rps12,atpB-rbcL,trnLintron,trnL-trnF,trnT (UGU)-trnL(UAA)等。本研究旨在利用上述序列探讨角果木属植物种间亲缘关系,构建系统发育树,并为相关物种的界定提供分子水平上的证据。
1 材料和方法
1.1实验材料
本研究所用植物材料包括角果木属24个个体,按Sheue等(2010)的形态特征描述分别属于5个不同的物种(表1),并基于前人的研究结果(Huang et al,2008)采用木榄作为本研究的外类群。采样时间跨度为2004-2013年,系本文作者所在实验室在多年的针对印度-西太平洋沿岸地区各种红树植物的实地采样过程中不断采集得到。角果木属内每种植物均包含3-6个分布在不同地理群体里的个体,目的是初步估计并比较物种内的遗传多态性水平。野外采集时将所采植物的新鲜叶片材料用硅胶快速干燥,带回实验室干燥柜中储存备用。凭证标本存于中山大学标本馆。
1.2 DNA提取
采用CTAB法提取总DNA(Doyle et al,1987)。所提样品DNA均存放在零下八十度超低温冰箱冻存。
1.3叶绿体序列的扩增、纯化及测序
叶绿体片段PCR扩增的引物来自Taberlet等(1991),Hodges等(1994)和Hamilton(1999)。反应体系为20 μl,包括DNA 20-40 ng,引物0.15 μmol/L,MgCl221.5 mol/L,dNTP 0.1 mmol/L,Taq DNA聚合酶1.5 U。扩增程序为:94℃,5 min→94℃,45 s→56℃,50 S→72℃,80 s→32个循环→72℃,6 min。PCR产物经琼脂糖电泳检测后直接测序,测序引物与PCR反应引物相同,采用双向测序。
1.4序列分析和系统树的构建
所测序列经人工校正后提交至GenBank(登录号见表1,所有125个序列均为本研究所测),进一步采用Clustal X 1.8软件进行排序和格式转换后,采用PAUP(Phylogenetic Analysis Using Parsimony)Version 4.0进行最大简约法(Maximum Parsimony,MP)构树,空位(gap)处理为缺失状态。采用启发式搜寻,TBR枝长交换以及500次随机序列加入寻找最大简约树。采用自展分析(bootstrap)检验简约树中各分支的置信度,自展数据集为1 000次。贝叶斯树(Bayesian inference,BI)的构建则采用Mrbayes V3.0 b4软件包完成。在构树前先采用Modeltest V 3.06获取最佳核苷酸替代模型,并估计相关参数。贝叶斯法所用Markov Chain Monte Carlo(马尔可夫链蒙特卡洛,MCMC)参数如下:以随机树为起始树,number of generations(代数)= 1000000,number of chains(链数)= 4,sample frequency(取样频率)= 100,MCMC达到平衡后,舍弃老化样本数(burnin)占总数的25%,系统树各分支的置信度以后验概率(Posterior Probability,PP)的形式来体现。此外,应用MEGA(Molecular Evolutionary Genetics Analysis)Version 5来计算种间遗传距离以及种内的遗传多态性水平,遗传距离模型选择Kimura 2-parameter模型,并进行邻接法构树(Neighbor-Joining,NJ)。NJ树同样采用自展值检验分支的置信度,自展数据集为1000次。

表1 实验材料来源及GenBank序列号

表2 基于5个叶绿体非编码区合并序列的角果木属物种间遗传距离和种内多态性
2 结果
所分析样品的5个叶绿体非编码区段的序列矩阵经Clustal X 1.8排列后总长为2716 bp。由于叶绿体在遗传上是作为一个整体(单倍型)。因此,我们将同一样品的所有5个叶绿体非编码区段的序列数据合并进行分析,以最大程度获取系统发育的信息。利用Kimura 2-parameter模型计算得到的物种间的遗传距离见表2,角果木属植物间的遗传距离介于0.003~0.027,遗传距离最近的两个物种是角果木与澳洲角果木(0.003),遗传距离最远的两个物种是角果木(澳洲角果木)与十雄角果木C. decandra之间(均为0.027)。十雄角果木物种复合体所包含的3个物种相互之间的遗传距离介于0.010~0.019之间,均大于角果木与澳洲角果木之间的遗传距离。物种内的遗传多态性检测结果见表2,分析结果表明5个物种的遗传多样性水平均较低。角果木、澳洲角果木与拟十雄角果木没有检测到多态性,而十雄角果木C. decandra和齐氏角果木C. zippeliana的种内多态性水平均为千分之一。
最大简约分析共有109个信息位点,采用启发式搜寻最终得到26棵最简约树,步长为183,一致性指数(CI)和维持性指数(RI)分别为0.956 和0.986。图2为其严格一致树并给出了重复1000次计算所得到的各分支的Bootstrap值。应用Modeltest进行模型选择所选的最适替代模型为GTR + G,由该程序所获得的替代模型参数运用在下一步的贝叶斯构树分析中。贝叶斯法构建系统发育树的结果见图3,各分支的后验概率值(PP)也同时列出。比较最大简约法、贝叶斯法和邻接法(未列出)的系统发育结果发现,3种方法所分别构建的系统发育树在拓扑结构上高度一致,角果木属植物均分为两个大的分支:(1)角果木和澳洲角果木构成一个分支;(2)属内其余植物,即十雄角果木复合体内的3个物种组成另一分支,该分支中齐氏角果木和拟十雄角果木为姊妹种,而十雄角果木与两者的关系相对较远(图2,3),在分支的支持率上,属内各物种的对应分支均得到极高的支持,其单系性均得到验证。
3 讨论
角果木属作为红树科海岸红树林植物4属之一,具有典型的红树植物形态特征,例如胎萌和发达的根系等等,以适应于海岸潮间带生境(Tomlinson,1986)。但该属的系统发育和分类研究则一直存在争议。Hou(1958)认为该属存在两个种:十雄角果木C. decandra(Griff.)Ding Hou和角果木C. tagal(Perr.)C. B. Robinson。White (1926)和Tomlinson(1996)则进一步在澳大利亚、新几内亚及其邻近岛屿鉴定出一个新的变种C. tagal(Perr.)C. B. Rob. var. australis C. T. White。Ballment等(1988)通过进一步的等位酶分析,将该亚种由亚种地位上升为种,命名为澳洲角果木C. australis(C. T. White)E. R. Ballment,T. J. Smith & J. A. Stoddart。目前澳洲角果木的种的地位已得到较多研究结果的支持(Duke,2006;Huang et al,2008;Sheue et al,2009b)。在形态特征上,角果木与澳洲角果木非常相似,最显著的区别在于胎生苗(胚轴)的形态,前者的胎生苗有棱角,而后者为圆柱状(Sheue et al,2009b)。本研究的分子系统发育研究结果同样支持了角果木与澳洲角果木各自的单系类群地位(100%支持率,图2,3),以及两种之间的姊妹群关系。这两个种组成了角果木属的第一大分支(100%支持率)。另外值得注意的是,本研究中的6个角果木个体分别来自不同地理区域的群体(包括东印度洋沿岸,西太平洋沿岸和澳大利亚),在一定程度上反映了角果木的分布范围。而种内多态性分析的结果表明,角果木作为该属分布最广的种,与分布区狭窄的澳洲角果木相比其种内多样性水平却都接近于零(表2),尽管这只是基于少数个体水平上的初步估计,但这一结果同样印证了Huang等(2012)对角果木群体遗传研究的结果,表明该物种存在一定程度上的遗传瓶颈效应。
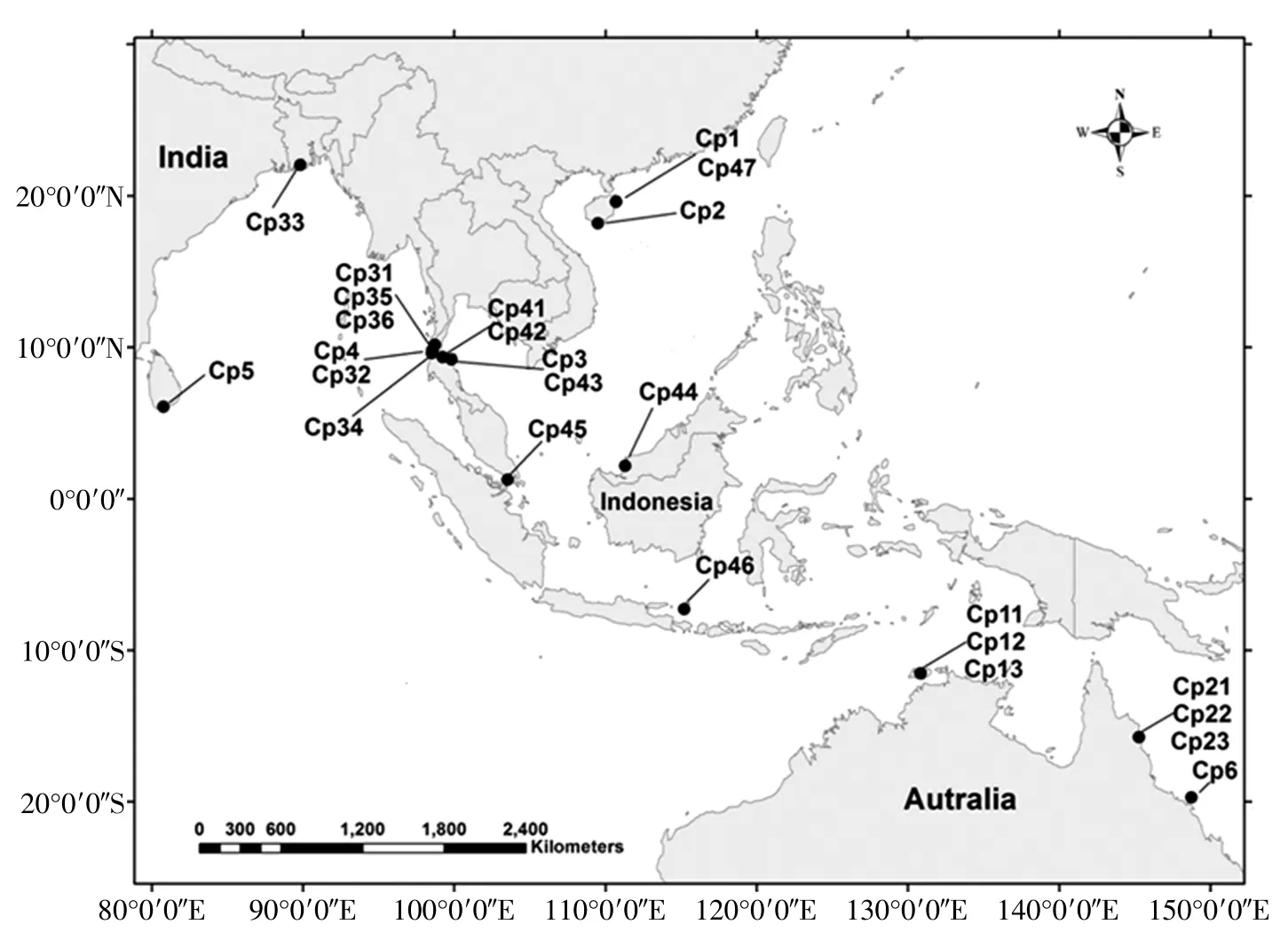
图1 角果木属物种和外类群的样品地理分布图

图2 基于叶绿体合并序列的角果木属最大简约树(MP)

图3 基于叶绿体合并序列的角果木属贝叶斯树(BI)
在本研究的分子系统发育树中角果木属的第2大分支对应着传统意义上的角果木属的第3个种:十雄角果木C. decandra(Griff.)Ding Hou。其支持率也达到了100%。但该分支进一步分化成3个亚分支;分别对应着3个经重新界定和命名的物种(Sheue et al,2010):这3个种的重新界定和命名主要是基于形态学上的考量。Sheue等(2009a;2010)在印度西太平洋地区海岸红树植物实地调查过程中对角果木属的植物材料进行了重新整理审视,认为基于萼筒形状,花瓣被毛特征和花的数目等特征可以把传统意义上的十雄角果木划分为3个种,十雄角果木C. decandra(Griff.)Ding Hou,齐氏角果木C. zippeliana Blume和拟十雄角果木C. pseudodecandra Sheue,Liu,Tsai & Yang。具体来说,十雄角果木的花萼呈现半球型,5~9 mm高;子叶下胚轴长8~13 cm、宽0.5~0.7 cm,顶端较圆。而齐氏角果木的花萼则呈较浅的圆盘型,2-3mm高;子叶下胚轴长9~17 cm、宽0.7~0.8 cm,顶端尖锐。拟十雄角果木花萼也是半球形,高度介于十雄角果木和齐氏角果木之间,4~5 mm;子叶下胚轴长10~16 cm、宽0.5~0.8 cm,顶端极其尖锐(Sheue et al,2010)。但在进一步的实地采样中发现这些特征上的差异仍然相对较小,各种性状往往相互交叉。因此利用分子序列上的证据来进一步验证这些物种是非常有必要的,尽管在划分这些物种时,Sheue等(2009a;2010)也采用了分子数据,但均基于一个基因片段,分辨率和可靠性存在疑问,也没有足够的支持率回答这3个种哪两个为姊妹种的问题。现在重建的分子系统发育树基于5个叶绿体分子片段,其所蕴含的分子系统发育信息非常清晰的将这3个种分开成独立的3个分支,其中齐氏角果木和拟十雄角果木为姊妹种。而且这些分支之间的遗传距离取值都在0.010以上,均大于两个广泛接受的物种即角果木与澳洲角果木之间的遗传距离0.003,这进一步支持了这些分支所代表的种的物种地位。
将本研究所获得分子系统发育树与各物种现有的地理分布相结合,会发现该复合体中存在一个明显的生物地理模式:即异域式物种形成模式。在印度-西太平洋地区,由于红树植物(尤其是红树科植物)的胎生苗是通过海洋进行传播的,因此普遍认为该地区由于海平面升降所形成的几处陆地障碍会影响红树植物的分化乃至物种形成(Huang et al,2008;Benzie et al,1998;Benzie et al,1999;Liao et al,2007)。其中马来半岛和印度尼西亚群岛作为历史上在该地区曾出现的基因流地理障碍对该地区的红树植物的遗传分化可能有着显著的影响(Huang et al,2008;Tan et al,2005)。在本研究中所揭示的分子系统发育树中,十雄角果木,齐氏角果木和拟十雄角果木这3个种的地理分布恰好对应着这两个历史地理障碍所分割的区域:十雄角果木的分布范围是印度,孟加拉,缅甸和泰国西海岸;齐氏角果木的分布范围大致是马来半岛东岸和南岸,南中国海周边,菲律宾,以及印尼的北部和中部地区;拟十雄角果木的分布范围限于澳大利亚,巴布亚新几内亚和印尼的南部部分地区(Sheue et al,2010)。这一分布模式暗示着由历史上的几次隔离分化事件驱动了相应的物种形成,并且进一步可由在系统树上3个物种的系统发育关系推论出马来半岛所引起的隔离分化事件相对更早。历史上,由马来半岛所引起的异域式物种形成在该地区已有记录和研究(Tomlinson,1986)。例如红树植物阿吉木属Aegialitis的一个种A. annulata仅分布在马来半岛的印度洋一侧,而它的姊妹种A. rotundifolia则分布在马来半岛的太平洋一侧(Duke,2006)。是否该地区有更多的红树植物具有类似的物种形成模式?同时,本属另一个广泛分布的物种(角果木)为何在这些区域之间没有显著的遗传隔离模式?对这些问题的探索是进一步研究的方向,而采用多基因的序列在统计学意义上进行物种界定与物种形成分析的研究是回答这些问题所必需的,进一步的研究正在进行中。
参考文献
Ballment E R,Smith T J,Stoddart J A,1988. Sibling species in the mangrove genus Ceriops(Rhizophoraceae),Detected using Biochemical Genetics. Australian Systematic Botany,1:391-397.
Benzie J A H,1998. Genetic structure of Marine Organisms and SE Asian Biogeography. In:Hall R,Holloway J D.(Eds.),Biogeography and Geological Evolution of SE. Asia. The Netherlands:Backhuys Publishers:197-209.
Benzie J A H,1999. Major genetic differences between Crown-of-Thorns Starfish(Acanthaster planci)Populations in the Indian and Pacific Ocean. Evolution,53:1782-1795.
Duke N C.1992. Mangrove Floristics and Biogeography. In:Robertson A,Alongi D.(Eds.),Tropical mangrove ecosystems. Washing DC:American Geophysical Union:63-100.
Duke N C,2006. Australia's Mangroves:the authoritative guide to Australia's mangrove plants. Brisbane:University of Queensland.
Doyle J J,Doyle J L,1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemistry Bulletin,19:11-15.
Hamilton M B,1999. Four primer pairs for the amplification of chloroplast intergenic regions with intraspecific variation. Molecular Ecology,8:521-523.
Hodges S A,Arnold M L,1994. Columbines:a geographically widespread species flock. Proceedings of the National Academy of Sciences USA,91:5129-5132
Hou D. 1958. Rhizophoraceae. In van Steenis C G G J.(Eds.),Flora Malesiana,ser. 1,vol. 5. Djakarta:Noordhoff- Kolff N V:429-473.
Huang Y L,Tan F X,Su G H,et al,2008. Population genetic structure ofthreetreespeciesin themangrovegenus Ceriops(Rhizophoraceae)from the Indo West Pacific. Genetica,133:47-56.
Huang Y L,Zhu C C,Li X,et al,2012. Differentiated population structure of a genetically depauperate mangrove species Ceriops tagal revealed by both sanger and deep sequencing. Aquatic Botany,101:46-54.
Liao P C,Havanod S,Huang S,2007. Phylogeography of Ceriops tagal (Rhizophoraceae)in Southeast Asia,The Land Barrier of The Malay Peninsula has caused population differentiation between The Indian Ocean and South China Sea. Conservation Genetics,8:89-98.
Sheue C R,Liu H Y,Tsai C C,et al,2009a. On the morphology and molecular basis of segregation of two species Ceriops zippeliana Blume and C. decandra(Griff.)Ding Hou(Rhizophoraceae)from Southeastern Asia. Blumea,54:220-227.
Sheue C R,Yang Y P,Liu H Y,et al. 2009b. Reevaluating the taxonomic status of Ceriops Australis(Rhizophoraceae)based on morphological and molecular evidence. Botanical Studies,50:89-100.
Sheue C R,Liu H Y,Tsai C C,et al,2010. Comparison of Ceriops pseudodecandasp. Nov.(Rhizophoraceae),a new mangrove species in Australasia,with related species. Botanical Studies,51:237-248.
Tan F X,Huang Y L,Ge X J,et al,2005. Population genetic structure and conservation implications of Ceriops decandra in Malay Peninsula and North Australia. Aquatic Botany,81(2):175-188.
Taberlet P,Gielly L,Pautou G,et al,1991,Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Molecular Biology,17:1105-1109.
Tomlinson P B,1986. The Botany of Mangroves. Cambridge:Cambridge University Press.
Tsai C C,Li S J,Su Y Y,et al,2012. Evidence of natural hybridization and gene introgression among Ceriops species(Rhizophoraceae)based on nuclear and plastid DNA. Biochemical Systematics and E-cology,43:178-191.
White C T,1926. A variety of Ceriops tagal C. B. Rob.(= C. candollean W. and A.). The London journal of botany,64:220-221.
陈小勇,林鹏,2000.我国红树植物分布的空间自相关分析.华东师范大学学报,3:104-109.
梁士楚,2000.广西红树植物群落特征的初步研究.广西科学,7 (3):210-216.
王文卿,王瑁,2007.中国红树林.北京:科学出版社.
(本文编辑:李晓光)
Molecular phylogeny of mangrove genus Ceriops(Rhizophoraceae)
ZHU Chun-chao1,TAN Feng-xiao2,LIU Ying1,QIANG Yin-meng1,SHI Su-hua1,HUANG Ye-lin1
(1. Guangdong Provincial Key Laboratory of Plant Resources,School of Life Sciences,Sun Yat-Sen University,Guangzhou 510275,China;2. South China Agricultural University,Guangzhou 510642,China)
Abstract:Five non-coding regions of chloroplast,including rpl20-rps12,atpB-rbcL,trnL intron,trnL-trnF and trnT (UGU)-trnL(UAA),from 24 individuals of five species in Ceriops and one outgroup species Bruguiera gymnorrhiza in Rhizophoraceae were determined by sequencing their PCR products directly. All 125 sequences of the five non-coding regions from different species were aligned and resulted in 2716 bp. The maximum parsimony,Bayesian inference and neighbor-joining methods were adopted to reconstruct molecular phylogeny of Ceriops based on the combined sequences,respectively. All phylogenetic trees showed consistent topology which revealed two clades in Ceriops,including one clade consisting of C. tagal and C. australis,and the other clade corresponding to C. decandra complex(C. decandra,C. pseudodecandraand C. zippeliana). The pairwise genetic distances among species of C. decandracomplex were significantly higher than those between two well-recognized species C. tagal and C. australis. And the monophyly of different species and the sistership of C. pseudodecandraand C. zippelianawere well supported. Our result supported Sheue's recent studies on the treatment of these species status in Ceriops. The article further discussed the association between the molecular phylogeny and the biogeographical model in Ceriops.
Keywords:mangrove;Ceriops;molecular phylogeny
中图分类号:P735;Q941
文献标识码:A
文章编号:1001-6932(2016)02-0209-07
Doi:10.11840/j.issn.1001-6392.2016.02.012
收稿日期:2015-01-25;
修订日期:2015-06-30
基金项目:国家自然科学基金(41276107;31200466;J1210074;J1310025);广东省自然科学基金(2015A030313136)。
作者简介:朱春超(1986-),女,博士研究生,主要从事分子生态学研究。电子邮箱:zhuchunchao11@163.com。
通讯作者:黄椰林,博士,副教授。主要从事红树植物分子生态与群体遗传研究。电子邮箱:lsshyl@mail.sysu.edu.cn。
