
嗜盐四联球菌的分离及其精氨酸代谢
2016-05-23 07:29廖淡宜堵国成江南大学工业生物技术教育部重点实验室江苏无锡214122江南大学生物工程学院江苏无锡214122江南大学食品安全与营养协同创新中心江苏无锡214122
食品与生物技术学报 2016年1期
廖淡宜, 方 芳*, 陈 坚, 堵国成, 刘 龙(1.江南大学工业生物技术教育部重点实验室,江苏无锡214122;2.江南大学生物工程学院,江苏无锡214122;3.江南大学食品安全与营养协同创新中心,江苏无锡214122)
嗜盐四联球菌的分离及其精氨酸代谢
廖淡宜1,2,3,方芳*1,2,3,陈坚1,2,3,堵国成1,2,刘龙1,2
(1.江南大学工业生物技术教育部重点实验室,江苏无锡214122;2.江南大学生物工程学院,江苏无锡214122;3.江南大学食品安全与营养协同创新中心,江苏无锡214122)
摘要:嗜盐四联球菌是存在于酱醪发酵中后期的嗜盐乳酸菌,其代谢精氨酸的特性与氨基甲酸乙酯前体物瓜氨酸的积累存在密切关系。为了降低酱油发酵过程中氨基甲酸乙酯前体物瓜氨酸的积累,研究采用选择性培养基以及groEL基因特异性对嗜盐四联球菌进行了分离和筛选,得到一株Tetragenococcus halophilus R23。对该菌株在不同培养温度和盐浓度下的精氨酸代谢特性进行了考察。结果表明:在30℃,180 g/L NaCl培养条件下,嗜盐四联球菌R23消耗精氨酸和瓜氨酸能力最强,消耗量分别为100.0%和64.3%。其中,代谢精氨酸的培养基中未检测到瓜氨酸的积累。这一特性对降低酱油发酵过程中氨基甲酸乙酯前体物瓜氨酸的积累具有重要意义。
关键词:嗜盐四联球菌;酱油;氨基甲酸乙酯;精氨酸;瓜氨酸
近年来,与我国传统发酵食品相关的食品安全事件的不断出现,引起了人们的广泛关注。影响传统食品安全最为重要的因素之一是由于各种发酵过程中含氮化合物的不完全代谢而生成的胺(氨)类物质,如氨基甲酸乙酯(Ethyl carbamate,EC),生物胺等。其中,氨基甲酸乙酯是影响范围最广的一种。国际癌症机构于2007年正式将氨基甲酸乙酯归为2A类致癌物(具有潜在致癌性)[1]。它的存在严重影响了我国传统发酵食品的安全性,使其出口贸易受到限制[2]。酱油是一种具有中国传统特色的调味品,近年来却检测出EC,这引起了专家的重视。目前,国内关于酱油中EC的报道仅集中在检测方法上[3-4];已有研究表明瓜氨酸是酱油酿造中形成EC的主要前体物质之一,并认为瓜氨酸积累与酱油发酵过程中的乳酸菌的精氨酸脱亚氨基(Arginine deiminase pathway,ADI)途径受到高盐环境抑制相关[5]。ADI途径中的3个关键酶分别是精氨酸脱亚氨酶、鸟氨酸氨甲酰基转移酶、氨基甲酸激酶。分别催化精氨酸转化成为瓜氨酸,瓜氨酸转化为鸟氨酸[6]得到的产物再进一步分解产生ATP、CO2和NH3。
嗜盐四联球菌属乳酸菌,以前被归类为嗜盐片球菌属,现在已单列为四联球菌属,广泛存在于如酱油、果酱等高盐或高糖环境中[7-8],是产生挥发性风味物质的主要微生物之一[9]。研究发现瓜氨酸主要是在酱醪生产前期积累,而嗜盐四联球菌主要存在于酱醪生产的中后期[5],已报道的嗜盐四联球菌全基因数据显示其具有完整的ADI代谢途径。因此,研究嗜盐四联球菌代谢精氨酸的能力和是否积累瓜氨酸对研究酱油EC前体物质瓜氨酸的积累机制具有重要意义。
作者从酱醪中分离嗜盐四联球菌,研究其精氨酸、瓜氨酸代谢能力,成功筛选到一株能将精氨酸和瓜氨酸转化为风味氨基酸鸟氨酸,且无瓜氨酸积累的嗜盐四联球菌,并初步研究了该菌的培养条件,为降低酱油中氨基甲酸乙酯的含量提供了一条有效途经。
1 材料与方法
1.1材料
1.1.1样品来源分离菌株的样品取自我国南方某酱油厂发酵180 d的酱醪。
1.1.2培养基分离培养基(g/L):蛋白胨10.0,牛肉膏10.0,酵母膏5.0,柠檬酸氢二铵2.0,葡萄糖20.0,吐温80 1.0,乙酸钠5.0,三水合磷酸氢二钾2.0,七水合硫酸镁0.58,一水合硫酸锰0.25,琼脂18.0,氯化钠100.0,溴甲酚紫0.06,生酱油100.0,制霉菌素50 U/mL,pH 6.0;种子培养基:MRS液体培养基,氯化钠100.0 g/L;精氨酸利用检测培养基(g/L):酵母膏5.0,牛肉膏5.0,胰蛋白胨5.0,氯化钠180.0,葡萄糖0.5,吐温80 1.0,七水合硫酸镁0.2,一水合硫酸锰0.05,硫酸铁0.4,柠檬酸三铵2.0,碳酸钙0.1,吡哆醛-5-磷酸0.05,磷酸氢二钾2.0,精氨酸5.0,pH 6.0[10];瓜氨酸利用检测培养基(g/L):酵母膏5.0,牛肉膏5.0,胰蛋白胨5.0,氯化钠180.0,葡萄糖0.5,吐温80 1.0,七水合硫酸镁0.2,一水合硫酸锰0.05,硫酸铁0.4,柠檬酸三铵2.0,碳酸钙0.1,吡哆醛-5-磷酸0.05,磷酸氢二钾2.0,瓜氨酸3.0,pH 6.0[10]。
1.1.3主要试剂和仪器基因组提取试剂盒:E.Z. N.A Bacterial DNA Kit D3350-01:购自上海生物工程有限公司;MRS培养基:购自OXIOD公司;HPLC(安捷伦1200):购自安捷伦科技公司;DNA maker:购自TaKaRa公司;PCR仪:购自美国Bio-Rad公司;厌氧培养箱:购于日本三洋公司。
1.2菌株分离
从发酵第180 d的酱醪中取样,酱醪悬液经过梯度稀释后涂布于分离平板上,30℃培养7 d。挑选表面光滑、不透明、乳白色小单菌落。将单菌落在斜面上划线分离两次后接单菌落到种子培养基中培养4 d,提取基因组,根据嗜盐四联球菌groEL基因序列,利用Primer软件设计引物扩增groEL基因,引物序列如下:groELF:5’- CGTCGTTCAATGCTTA ATGG -3’;groELR:5’- TGCTGCCAGAAGAAACTT CA -3’
以嗜盐四联球菌基因组为模板,PCR扩增条件:95℃预变性5 min,30个循环,95℃变性30 s,54℃退火30 s,72℃延伸30 s。2%琼脂糖凝胶电泳检测扩增结果。
1.3菌株种属鉴定及电镜样品制备
将groEL基因扩增结果显阳性的菌株进行划线分离两次后接单菌落到种子培养基中培养4 d,提取基因组,利用细菌16S rDNA基因通用引物进行PCR扩增16S rDNA全基因序列。引物序列如下:27F:5’- AGAGTTTGATCCTGGCTCAG -3’;1492R:5’- GGTTACCTTGTTACGACTT -3’
以嗜盐四联球菌基因组为模板,PCR扩增条件:95℃预变性5 min,30个循环,95℃变性30 s,54℃退火30 s,72℃延伸1.5 min。得到的PCR产物送到上海生物工程有限公司测序,得到序列后通过NCBI数据库比对分析,确定菌株种属。
将培养好的菌体用2.5 g/dL戊二醛溶液固定,0.1 mol/L的磷酸缓冲液漂洗,滴样在透射电镜的3 mm铜网上,磷钨酸染色后在透射电镜下观察菌株的形态。
1.4精氨酸、瓜氨酸代谢
将-80℃保藏的嗜盐四联球菌划线复苏两次后挑单菌落接种于种子培养基中,30℃静置培养3 d,取4 mL菌液离心10 min(4 000 r/min,4℃)。弃掉上清后加入1 mL磷酸盐缓冲液(pH 7.0)悬浮菌体,离心10 min(4 000 r/min,4℃),弃掉上清,向离心管中加入4 mL氨基酸利用检测培养基。30℃静置培养5天,检测精氨酸、瓜氨酸和鸟氨酸含量。
1.5氨基酸测定
取1 mL相应培养液,离心10 min(4 000 r/ min,4℃),取上清液。用体积分数5%的三氯乙酸以1∶5比例稀释上清液,用孔径为0.22 μm的滤膜过滤取滤液。用高效液相色谱法(HPLC)测定样品中精氨酸、瓜氨酸和鸟氨酸含量。测定条件:色谱柱ODS-2 HYPERSIL。流动相A相(1 L):无水乙酸钠5 g,四氢呋喃5 mL,三乙胺200 μL,pH为7.2。流动相B相(1 L):无水乙酸钠5 g,超纯水200 mL,甲醇400 mL,乙腈400 mL,pH为7.2。检测器为VWD紫外检测器,检测波长338 nm,柱温40℃[11]。
1.6嗜盐四联球菌3 L发酵罐扩培
将-80℃保藏的嗜盐四联球菌划线复苏后挑单菌落接种于种子培养基活化两次,以体积分数4%的接种量接种到3 L发酵罐中,发酵培养基:MRS,NaCl 100 g/L,pH 7.0。培养条件:转速100 r/min,温度30℃。发酵70 h,平均每6 h取样测OD600、pH。
1.7嗜盐四联球菌低温生长和精氨酸、瓜氨酸代谢
将-80℃保藏的嗜盐四联球菌划线复苏后挑单菌落接种于种子培养基中,30℃静置培养3天,活菌计数为2.3×108CFU/mL,将其放入低温(15~20℃)下每隔5 d更换一次培养基并进行活菌计数,同时以30℃作为对照,研究嗜盐四联球菌在低温与常温下生长情况。分别取置于低温下培养5 d和20 d的菌液离心10 min(4 000 r/min,4℃)。弃掉上清后向菌体加入1 mL磷酸盐缓冲液(pH 7.0)悬浮菌体,离心10 min(4 000 r/min,4℃),弃掉上清,向离心管中加入4 mL氨基酸利用检测培养基。30℃静置培养5 d,检测精氨酸、瓜氨酸和鸟氨酸含量。
2 结果与分析
2.1嗜盐四联球菌的分离与鉴定
热休克蛋白GroEL是一种广泛存在于细菌中的分子伴侣蛋白,其行使的功能相似,但不同物种间基因序列相差较大,因此本研究根据Tetragenococcus halophilus的groEL基因序列设计特异性引物,通过PCR扩增来筛选Tetragenococcus halophilus。挑选groEL基因扩增结果为阳性的菌株进行16S rDNA基因序列测定,将测得的16S rDNA全基因序列与NCBI GenBank中的数据比对,结果显示与Tetragenococcus halophilus相似度大于99%的都是嗜盐四联球菌。用MEGA5.2.1软件以Neighbor-Joining法(Bootstrap值设定为1000,其他值为软件默认)构建系统发育树,筛选到一株菌与Tetragenococcus halophilus NBRC 12172聚到了同一个分支,亲缘关系最近。命名为Tetragenococcus halophilus R23。系统进化树如图1,Tetragenococcus halophilus R23电镜图如图2。
2.2嗜盐四联球菌利用精氨酸能力分析
研究表明在酱醪发酵过程中,微生物通过精氨酸脱亚氨基途径利用精氨酸转化为瓜氨酸,造成瓜氨酸的积累[5]。为了考察嗜盐四联球菌代谢精氨酸的能力,将T. halophilus R23接种于精氨酸利用检测培养基中培养,然后检测精氨酸、瓜氨酸和鸟氨酸的含量,结果见表1。T. halophilus R23在低盐条件下(100 g/L NaCl)利用精氨酸的量为5.0 g/L(利用率100.0%),高盐条件下(180 g/L NaCl)利用精氨酸的量为4.88 g/L(利用率97.6%),低于低盐条件下利用精氨酸的量,低盐和高盐条件下精氨酸转换成鸟氨酸的转化率分别为79.3%和92.9%,且均不积累瓜氨酸。
2.3嗜盐四联球菌利用瓜氨酸能力分析
瓜氨酸是酱醪发酵过程中形成氨基甲酸乙酯的主要前体物质之一,降低瓜氨酸的含量是减少氨基甲酸乙酯的有效措施之一。为了考察嗜盐四联球菌能否代谢瓜氨酸以及代谢能力,将T. halophilus R23接种于含有瓜氨酸的培养基中培养,然后检测瓜氨酸和其终产物鸟氨酸的含量,结果见表2。T. halophilus R23在低盐(100 g/L NaCl)和高盐(180 g/ L NaCl)条件下用瓜氨酸的利用率64.3%和17.7%,说明T. halophilus R23对瓜氨酸具有一定的代谢能力。且在低盐和高盐条件下瓜氨酸转换成鸟氨酸的转化率均达到100%,这一特性对降低酱醪发酵过程中瓜氨酸的含量具有重要意义。
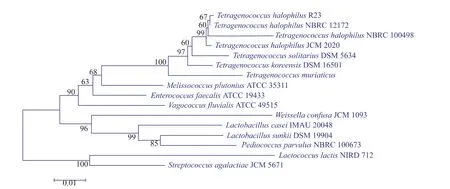
图1 R23基于16S rDNA序列的系统发育分析Fig. 1 Phylogenetic analysis of T. halophilus R23 based on its 16S rDNA sequence
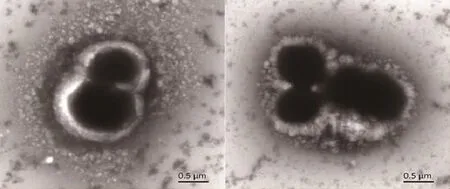
图2 T. halophilus R23电镜图Fig. 2 TEM observation of T. halophilus R23
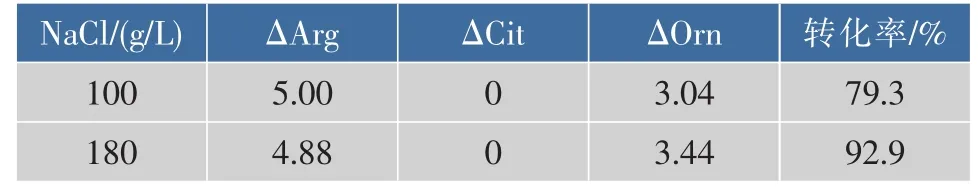
表1 NaCl质量浓度对嗜盐四联球菌R23精氨酸代谢的影响Table 1 Effect of salt on T. halophilus R23 arginine metabolism
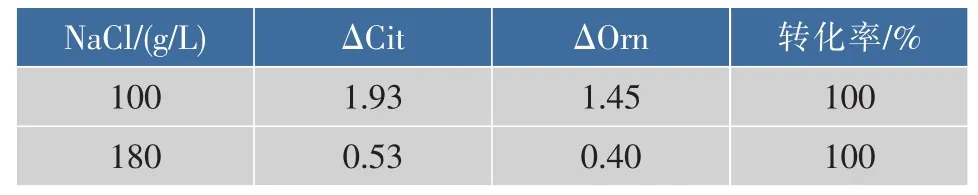
表2 嗜盐四联球菌R23消耗瓜氨酸能力Table 2 Detection of citrulline degradation by T. halophilus R23
2.4嗜盐四联球菌3 L发酵罐扩培
Natteewan等人已经采用添加嗜盐四联球菌作为一种工艺手段达到增加发酵食品风味的目的[8]。T. halophilus R23对培养条件要求较高,其生长受pH、溶氧、生长因子等限制。为获得适合高浓度酱醪发酵的T. halophilus R23菌液并验证扩培的可行性,将T. halophilus R23接种于合适的培养基进行纯种培养,测定OD600生长曲线和pH变化曲线。T. halophilus R23单菌落经两次摇瓶活化培养后接种于3 L发酵罐中扩大培养,生长曲线如图3。如图所示,0~8 h,菌生长处于延滞期,生长缓慢。8~30 h,菌体处于对数生长期,菌体大量繁殖,pH降低。30 h后菌体生长进入稳定期,此时菌体数目不再增加,pH也趋于稳定。35 h时检测到最大A600 nm=3.21,活菌计数为3.2×108CFU/mL。
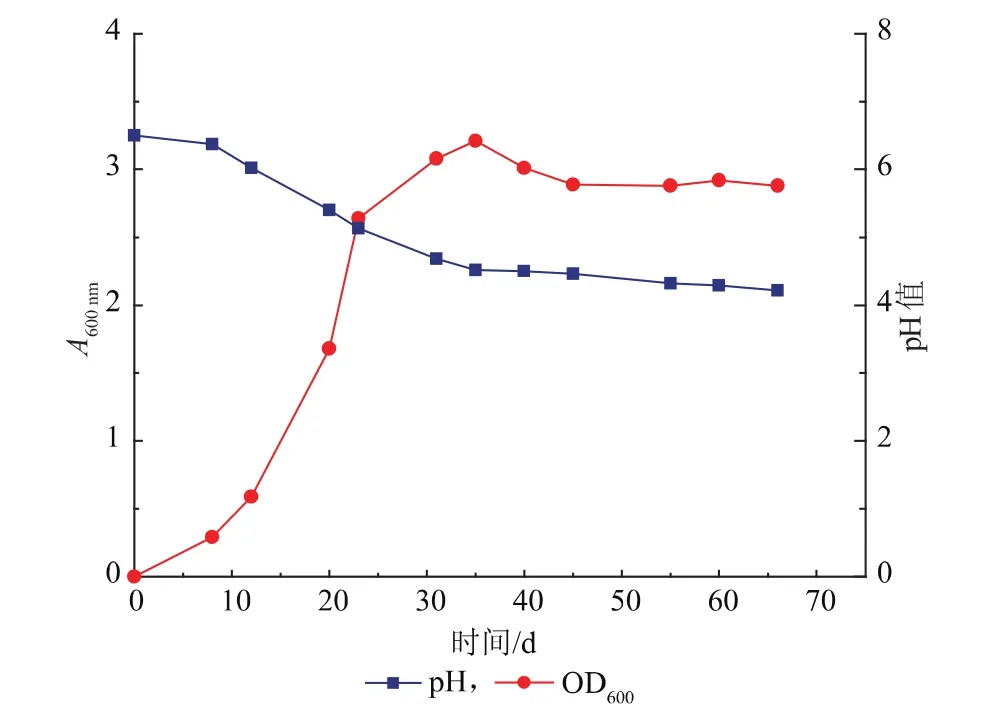
图3 T. halophilus R23 3 L发酵罐扩培生长曲线Fig. 3 Growth curve of T. halophilus R23 cultivated in a 3 L fermentor
2.5嗜盐四联球菌低温生长和精氨酸、瓜氨酸代谢
酱醪发酵前期控温包括低温(16℃)和常温(30℃)两个阶段。为了研究温度对T. halophilus R23生长和氨基酸代谢能力的影响,将T. halophilus R23在低温和常温下进行了连续培养,并测定了低温培养(15~20℃)后T. halophilus R23对精氨酸和瓜氨酸的利用量。T. halophilus R23在低温下随着培养时间的延长,活菌数下降,在常温下活菌数量随着培养时间的延长先上升后下降,第10 d达到最大值,为3.0×108CFU/mL。两个温度下T. halophilus R23活菌数均在第25 d到1×106CFU/mL以下。将低温培养一段时间后的T. halophilus R23接种于氨基酸利用检测培养基中培养,然后检测精氨酸、瓜氨酸和鸟氨酸的含量,结果见表3。低温放置20 d 后T. halophilus R23利用精氨酸和瓜氨酸的能力仍然存在,但明显低于低温放置5 d后T. halophilus R23利用精氨酸和瓜氨酸的能力。
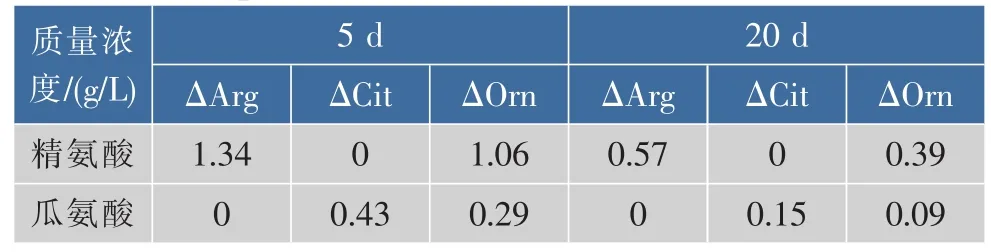
表3 低温培养对嗜盐四联球菌R23利用精氨酸、瓜氨酸能力的影响Table 3 Effect of cultivation of T. halophilus R23 on its degradation of arginine and citrulline at low temperature
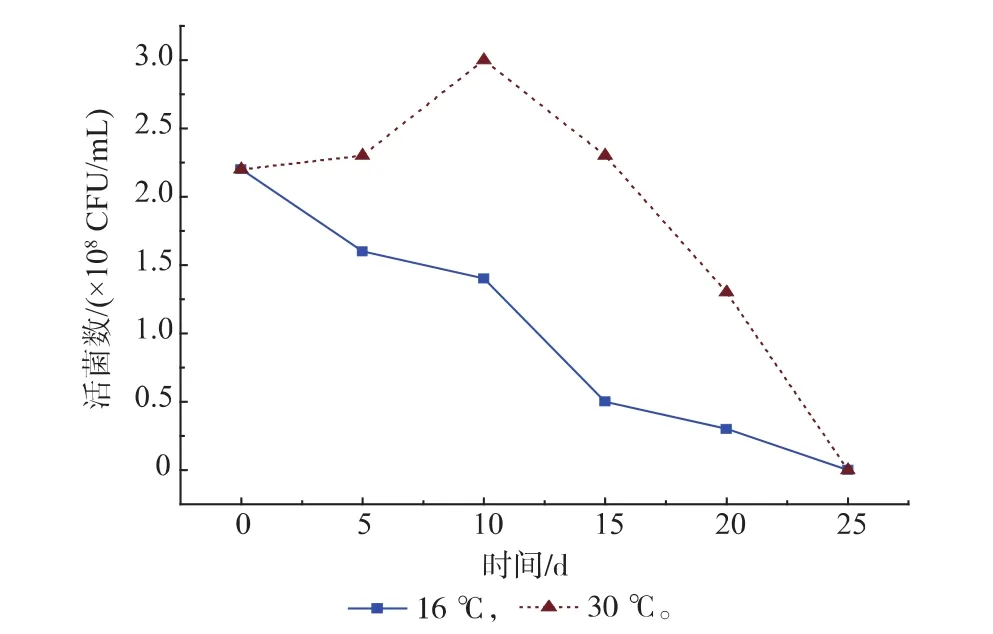
图4 T. halophilus R23不同温度下生长曲线Fig. 4 Growth curve of T. halophilus R23 cultivated at different temperature
3 结语
20世纪40年代,Nettleship实验证明了氨基甲酸乙酯具有致癌作用[12],随后,在酱油等大豆发酵制品中检测出了氨基甲酸乙酯。已有研究表明瓜氨酸是酱油发酵过程中形成氨基甲酸乙酯的主要前体物质之一,且温度和盐浓度等环境因素会影响乳酸菌的精氨酸脱亚氨基途径,从而影响环境中瓜氨酸的含量[13]。
作者从我国南方某酱油厂发酵180 d的酱醪中筛选到一株嗜盐四联球菌株T. halophilus R23,通过对它在高盐和低盐条件下精氨酸和瓜氨酸代谢能力的考察,发现T. halophilus R23在高盐环境中能将精氨酸和瓜氨酸转化为鸟氨酸,不仅消耗了瓜氨酸,同时增加了风味氨基酸鸟氨酸,对酱醪发酵起到增香的效果。T. halophilus R23在高盐条件下不积累瓜氨酸的特性与Pediococcus acidilactici在高盐条件下代谢受到抑制不同[5]。为了得到适合高浓度酱醪发酵的T. halophilus R23菌体,本研究进行了3 L发酵罐扩培,发现T. halophilus R23延滞期较长。为了缩短发酵时间,下一步可以对T. halophilus R23进行发酵条件优化。作者考察了酱醪发酵的最适添加T. halophilus R23时期以及温度对T. halophilus R23代谢精氨酸和瓜氨酸能力的影响,结合低温处理菌体细胞研究了T. halophilus R23生长情况。发现低温处理后的T. halophilus R23生长速率以及代谢能力下降。因此可以考虑在酱油发酵前期的常温发酵阶段添加T. halophilus R23来控制瓜氨酸的积累,从而达到降低酱油中氨基甲酸乙酯的目的。
参考文献:
[1]Canas B J,Joe F L,Diachenko G W,et al. Determination of ethyl carbamate in alcoholic beverages and soy sauce by gas chromatography with massselective detection:collaborative study[J]. Journal of AOAC International,1993,77(6):1530-1536.
[2]周景文,堵国成,陈坚.发酵食品有害氨(胺)类代谢物-形成机制和消除策略[J].中国食品学报,2011,11(9):8-18. ZHOU Jinwen,DU Guocheng,CHEN Jian. Harmful amines(ammonia)metabolites in fermented foods:formation mechanism and the elimination strategy[J]. Chinese Institute of Food Science,2011,11(9):8-18.(in Chinese)
[3]Huang Z,Pan XD,Wu PG,et al. Validation(in-house and collaboratory)of the quantification method for ethyl carbamate in alcoholic beverages and soy sauce by GC-MS[J]. Food Chemistry,2013,141(4):4161-4165.
[4]张燕,丰帆,姜小梅,等.顶空固相微萃取气相色谱质谱法检测酱油中氨基甲酸乙酯含量[J].食品与发酵工业,2010,1(5):34-40. ZHANG Yan,FENG Fan,JIANG Xiaomei,et al. The development of head space solid-phase microextraction and gas chromatography-tandem mass spectrometry for the detection of ethyl carbamate in soy sauce[J]. Food and Fermentation Industries Editorial Staff,2010,1(5):34-40.(in Chinese)
[5]Zhang J,Fang F,Chen J,et al. The arginine deiminase pathway of koji bacteria is involved in ethyl carbamate precursor production in soy sauce[J]. FEMS Microbiology Letters,2014,358(1):91-97.
[6]Kameya M,Asano Y. Rapid enzymatic assays for L-citrulline and L-arginine based on the platform of pyrophosphate detection[J]. Enzyme and Microbial Technology,2014,136(9):506-570.
[7]Juste A,Lievens B,Frans I,et al. Genetic and physiological diversity of Tetragenococcus halophilus strains isolated from sugarand salt-rich environments[J]. Microbiology,2008,154(9):2600-2610.
[8]Udomsil N,Rodtong S,Choi YJ,et al. Use of Tetragenococcus halophilus as a starter culture for flavor improvement in fish sauce fermentation[J]. Journal of Agricultural and Food Chemistry,2011,59(15):8401-8408.
[9]黄丹,梁源,左勇,等.酱油发酵酱醅中耐盐乳酸菌的分离筛选及产酸特性[J].食品与生物技术学报,2014,33(6):652-656. HUANG Dan,LIANG Yuan,ZUO Yong,et al. Isolation and screening of salt-tolerance lactic acid bacteria strains from soy sauce mash and study on its characteristic producing lactic acid[J]. Journal of Food Science and Biotechnology,2014,33(6):652-656.(in Chinese)
[10]Bover C S,Holzapfel W H. Improved screening procedure for biogenic amine production by lactic acid bacteria[J]. International Journal of Food Microbiology,1999,53(1):33-41.
[11]陈兵,王雅珍,黄徽微,等.反相高效液相色谱-质谱联用法测定抑菌肽功能单元的氨基酸序列[J].食品与生物技术学报,2013,9(5):522-527. CHEN Bin,WANG Yazheng,HUANG Huiwei,et al. Determination of amino acid sequence of antibacterial peptide function units by using RPHPLC-MS method[J]. Journal of Food Science and Biotechnology,2013,9(5):522-527.(in Chinese)
[12]耿予欢,李国基.关于酿造酱油中氨基甲酸乙酯的探讨[J].中国酿造,2003,2(1):31-33. GENG Yuhuan,LI Guoji. Research on the amino ethyl formate in brewing soy sauce[J]. China Brewing,2003,2(1):31-33.(in Chinese)
[13]Vrancken G,Rimaux T,Wouters D,et al. The arginine deiminase pathway of Lactobacillus fermentum IMDO 130101 responds to growth under stress conditions of both temperature and salt[J]. Food Microbiology,2009,26(7):720-727.
Isolation and Arginine Metabolism Detection of a Tetragenococcus halophilus Strain
LIAO Danyi1,2,3,FANG Fang*1,2,3,CHEN Jian1,2,3,DU Guocheng1,2,LIU Long1,2
(1. Key Laboratory of Industrial Biotechnology,Ministry of Education,Jiangnan University,Wuxi214122,China;2. School of Biotechnology,Jiangnan University,Wuxi 214122,China;3. Collaborative Innovation Center of Food Safety and Nutrition,Wuxi 214122,China)
Abstract:The Tetragenococcus halophilus species is a halophilic lactic acid bacteria(LAB)exhibiting in the later fermentation of soy sauce moromi. The arginine metabolism of this species is closely related to the accumulation of citrulline which is the main ethyl carbamate precursor in soy sauce. A Tetragenococcus halophilus strain R23 was isolated from moromi by selective medium cultivation and PCR(targeting on the groEL gene)screening. The arginine metabolism of the strain was investigated under different cultivation temperature and salt concentration. A highest utilization of arginine and citrulline for T. halophilus R23 was observed at 30℃in the presence of 180 g/L NaCl,with degradation rate of 100.0% and 64.3%,respectively. In addition,there was no citrulline detected with in arginine consumption of R23,which provided a potential application of citrulline reduction in soy sauce during fermentation process.
Keywords:Tetragenococcus halophilus,soy sauce,ethyl carbamate,arginine,citrulline
*通信作者:方芳(1976—),女,安徽枞阳人,哲学博士,副教授,主要从事微生物与食品生物技术研究。E-mail:ffang@jiangnan.edu.cn
基金项目:国家自然科学基金项目(3137821);国家“973”计划项目(2012CB720802)。
收稿日期:2014-11-24
中图分类号:Q 93-3
文献标志码:A
文章编号:1673—1689(2016)01—0048—06
猜你喜欢
小猕猴学习画刊(2018年4期)2018-01-25
现代检验医学杂志(2016年1期)2016-11-12
故事大王(2016年10期)2016-11-07
文理导航·趣味课堂(2016年6期)2016-09-09
中国免疫学杂志(2016年2期)2016-01-30
中外医疗(2015年11期)2016-01-04
中国当代医药(2015年30期)2015-03-01
疑难病杂志(2014年12期)2014-04-16
药学实践杂志(2011年3期)2011-12-08
植物营养与肥料学报(2011年6期)2011-10-24
