
棕榈蓟马对番茄斑萎病毒的传毒特征及其在日光温室的发生关系研究
2016-05-14 10:38张安盛毛晓红于毅周仙红庄乾营
山东农业科学 2016年5期
张安盛 毛晓红 于毅 周仙红 庄乾营
摘 要:通过室内PCR检测和日光温室定点调查,研究了棕榈蓟马对番茄斑萎病毒的传毒特征、番茄斑萎病毒病发生与棕榈蓟马种群动态的关系。结果表明,用带毒菜椒植株饲喂健康棕榈蓟马若虫24~96 h,棕榈蓟马带毒率达15.00%~18.33%,带毒棕榈蓟马成虫取食健康菜椒植株12~48 h,其理论传毒效率可达36.37%~59.10%,表明棕榈蓟马若虫具有一定获毒能力,其成虫具有一定传毒能力; 4月下旬之前,棕榈蓟马成虫数量较低,病毒病没有发生;4月下旬~5月中旬,棕榈蓟马成虫数量略有提高,病毒病开始出现,但菜椒植株带毒率较低;5月下旬~6月初,棕榈蓟马成虫数量增长迅速,菜椒植株带毒率亦呈快速增长状态,表明菜椒植株带毒率与棕榈蓟马成虫密度密切相关,且病毒病的发生具有一定滞后性。
关键词:棕榈蓟马;番茄斑萎病毒;传毒特征;发生关系
中图分类号:S436.412.2+9文献标识号:A文章编号:1001-4942(2016)05-0098-04
Abstract The transmission characteristics of Tomato spotted wilt virus (TSWV) by Thrips palmi and the relationship between the occurrence of TSWV and the population dynamics of T. palmi were studied by point-survey in solar greenhouse and PCR detection in the lab. The results showed that the virus carrying rate of T. palmi was 15.00%~18.33% when the healthy T. palmi nymphs were fed on viruliferous sweet pepper plants for 24~96 hours,and the theoretical virus transmission efficiency of T. palmi adults were 36.37%~59.10% when the viruliferous T. palmi adults were fed on healthy sweet pepper plants for 12~48 hours, which indicated that the T. palmi nymphs had certain virus acquisition ability and the adults had certain virus transmission ability. The number of T. palmi adults was low and the TSWV had not occurred before the last ten days of April. The number of T. palmi adults increased slowly and the TSWV appeared but the viruliferous percentage of sweet pepper plant was low from the last ten days of April to the middle ten days of May. The number of T. palmi adults increased rapidly and the viruliferous percentage of sweet pepper plant enhanced rapidly as well from the last ten days of May to the earlier ten days of June. Therefore, the viruliferous percentage of sweet pepper plant was closely related to the population density of T. palmi adults and the occurrence of TSWV lagged behind the the appearance of T. palmi adults.
Key words Thrips palmi; Tomato spotted wilt virus; Transmission characteristics; Occurrence relationship
番茄斑萎病毒(Tomato spotted wilt virus,TSWV)属于布尼亚病毒科(Bunyaviridae)番茄斑萎病毒属(Tospovirus),可侵染蔬菜、花生、大豆等多种农作物,是世界十大危险性植物病毒之一[1]。番茄斑萎病毒病1915年在澳洲被首次发现,而后在欧洲、美洲和非洲等地暴发,造成辣椒、番茄等蔬菜重大损失,甚至绝产[2]。在我国,番茄斑萎病毒最早于1984年在广州花生植株上被发现[3],而后在四川烟草上被报道[4]。自2000年起,该病毒病在云南省大面积危害,对该地区菜椒、番茄、烟草、花卉等作物生产造成重大损失[5~8]。2012年,番茄斑萎病毒病在北京市海淀区日光温室辣椒上发生[9],2014~2015年课题组在山东省济南等地菜椒植株上发现疑似病株。
番茄斑萎病毒传毒介体是蓟马[10],蓟马若虫期获毒,病毒在蓟马体内复制,成虫期将病毒传播到寄主植物[11]。棕榈蓟马(Thrips palmi)是番茄斑萎病毒重要的传毒介体,也是我国北方设施蔬菜蓟马类害虫的优势种,其发生区域、种群数量、危害程度远远高于花蓟马、烟蓟马和西花蓟马等其它蓟马类害虫。番茄斑萎病毒病已经在我国北方设施蔬菜上发生,其传播蔓延速度和危害程度,与棕榈蓟马的发生为害和传毒特征密切相关。本研究初步测定了棕榈蓟马对番茄斑萎病毒的传毒特征,调查了日光温室条件下番茄斑萎病毒病发生与棕榈蓟马种群动态的关系,以期为番茄斑萎病毒病的科学防控提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 主要仪器设备 RXZ-280D-LED光照培养箱(宁波江南仪器厂产); TP600PCR仪(TaKaRa公司产);凝胶成像仪(Alpha Innotech公司产); DYY-6C电泳仪(北京市六一仪器厂产);5804R低温冷冻离心机(Eppendorf公司产); MDF-U4086S -80℃冰箱(SANYO公司产);MLS-3020高压灭菌锅(SANYO公司产); SHZ-82型水浴恒温振荡器(江苏太仓医疗器械厂产);2.5~1 000 μL移液器(Eppendorf公司产);120目防虫网(浙江省台州市路桥区德农防虫网厂生产);养虫瓶(高10.3 cm,瓶口半径为3.1 cm的圆柱形糖果罐);吸虫器(由20 mL一次性塑料注射器改造而成);PVC网罩(将PVC膜做成直径12 cm、高度40 cm圆柱体,一端粘120目防虫网);微虫笼(将2个内径2 cm、高0.5 cm的有机玻璃环固定在条形发夹两翼,外部粘120目防虫网,发夹两翼闭合时两个有机玻璃环紧扣在一起,形成封闭的微虫笼,通过发夹将微虫笼固定在菜椒叶片上);塑料花盆(直径10 cm);培养皿(直径9.0 cm);封口膜;小毛笔。
1.1.2 试验试剂 TRIzol Reagent(天根生化科技有限公司产,目录号:15596026);TIANscript cDNA第一链合成试剂盒(天根生化科技有限公司产,目录号:KR104);DL2000 DNA Marker(北京全式金生物公司产); Pfu PCR MasterMix(天根生化科技有限公司产,目录号:KP201);RNAprep Pure植物总RNA提取试剂盒(天根生化科技有限公司产,目录号:DP432);dd H2O(上海生工生物有限公司产);氯仿、异戊醇、异丙醇、无水乙醇、Na2SO3、PVP、Na2HPO4·12H2O、NaH2PO4·2H2O(天津市大茂化学试剂厂产)。
1.1.3 其它试验材料 供试番茄斑萎病毒毒源:由云南农业大学提供,一部分置于-80℃冰箱中保存,另一部分用于菜椒植株接种;
健康菜椒植株:品种为“常丰大菜椒”,在以120目防虫网隔离的育苗盘中培养至四叶期,除去弱苗,单株移栽在花盆中作为健康试验植株;
带毒菜椒植株:选取四叶期的健康菜椒植株,采用人工摩擦接种方法接种番茄斑萎病毒,接种后置于温度(26±1)℃、相对湿度(70±5)%、光照L∶D=14∶8的光照培养箱中,15 d后做PCR检测,选取带毒植株作为试验植株;
健康棕榈蓟马:采自济南市商河县玉皇庙街道办事处西甄村日光温室菜椒植株,室内用芸豆连续多代饲养,建立稳定种群;
带毒棕榈蓟马:在以120目防虫网隔离的带毒菜椒苗上接健康棕榈蓟马,持续饲养2个月,选取初羽化棕榈蓟马成虫,一部分做PCR检测,明确其带毒率,另一部分用于棕榈蓟马传毒试验。
1.2 试验方法
1.2.1 棕榈蓟马若虫对番茄斑萎病毒的获毒能力 选取带毒菜椒植株上的嫩叶,置于培养皿中,下垫湿润滤纸保湿。在叶片上接健康棕榈蓟马1龄若虫(孵化6 h内),用封口膜将培养皿封口后置于温度(26±1)℃、相对湿度(70±5)%、光照L∶D=14∶8的光照培养箱中。分别在接虫后0.5、1、3、6、12、24、48、96 h将棕榈蓟马若虫转移到养虫瓶中,在相同温湿度条件下用芸豆分别饲养,每个时间处理重复3次。待棕榈蓟马成虫羽化后,用PCR方法单头检测棕榈蓟马雌成虫带毒情况,每重复检测棕榈蓟马雌成虫20头,计算带毒率。
1.2.2 棕榈蓟马成虫对番茄斑萎病毒的传毒能力 将带毒棕榈蓟马雌成虫置于微虫笼中,每微虫笼放1头;将微虫笼固定在健康菜椒叶片上,每株菜椒植株固定微虫笼1个,菜椒植株接虫后罩PVC网罩。分别在接虫后0.5、1、3、6、12、24、48、96 h去除微虫笼并杀死笼中蓟马,使菜椒苗在PVC网罩中继续生长,每个时间处理重复3次,每重复菜椒苗40株。20 d后用PCR方法单株检测菜椒苗带毒情况,用菜椒苗带毒株率除以棕榈蓟马带毒率,即棕榈蓟马理论传毒效率。
1.2.3 日光温室中番茄斑萎病毒病的发生与棕榈蓟马种群动态的关系 试验在山东省农业科学院植物保护研究所日光温室中进行,日光温室面积约200 m2。菜椒定植时间为2015年3月10日,每666.7m2栽2 400株左右,长势良好;定植时在菜椒行间均匀放入4株盆栽带毒菜椒,作为毒源植株。4月初~6月初调查日光温室中棕榈蓟马和番茄斑萎病毒病发生情况,每隔10 d左右调查一次,共调查7次。调查方法:在日光温室东、西两侧分别保留1垅菜椒做保护行,将日光温室自东向西平均分为5小区,在每个小区的中间位置选取菜椒2行,在每行菜椒的南、中、北各标记5株代表性菜椒植株,每株菜椒上选取4朵花,调查棕榈蓟马成虫数量,同时采集标记菜椒上的叶片,室内PCR检测菜椒植株带毒率。
2 结果与分析
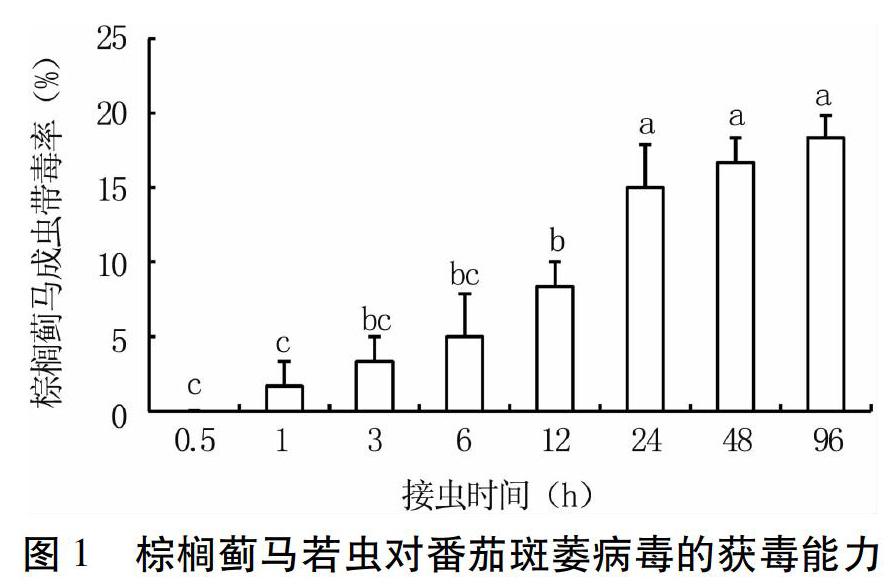
2.1 棕榈蓟马若虫对番茄斑萎病毒的获毒能力
棕榈蓟马若虫一旦获毒会终生带毒,其成虫带毒率可以反映若虫获毒能力,成虫带毒率越高,若虫获毒率越高,反之亦然。试验结果(图1)表明,棕榈蓟马若虫的获毒能力随着饲喂带毒菜椒植株时间的延长逐渐提高。健康棕榈蓟马若虫在带毒菜椒植株上饲喂0.5 h,棕榈蓟马成虫不带毒;饲喂1 h后棕榈蓟马成虫开始带毒,但带毒率极低,只有1.67%;饲喂3~12 h,棕榈蓟马成虫带毒率较低,仅为3.33%~8.33%,处理间无显著差异;饲喂24~96 h,棕榈蓟马成虫带毒率略有提高,达15.00%~18.33%,该时间段不同处理间无显著差异,但显著高于饲喂12 h处理。
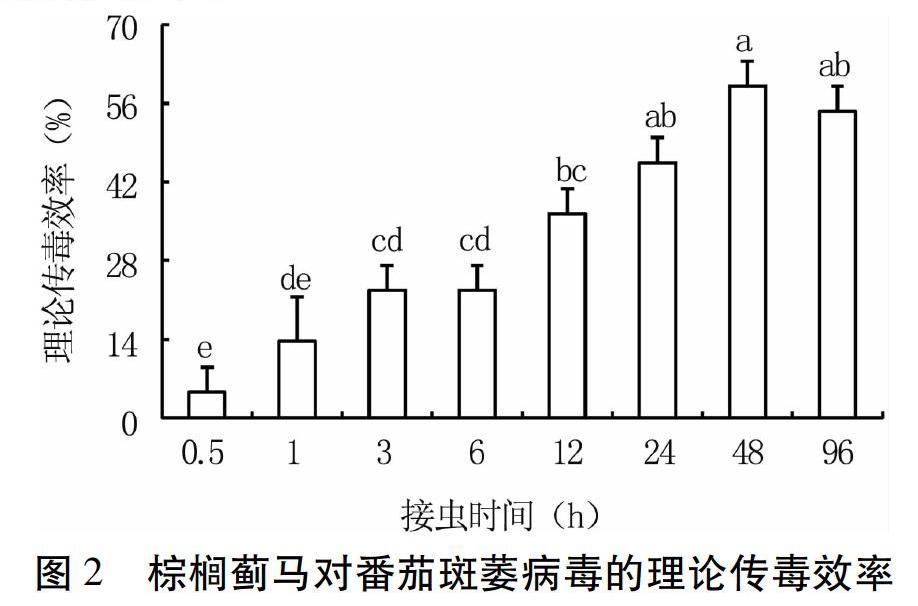
2.2 棕榈蓟马成虫对番茄斑萎病毒的传毒能力
试验结果(图2)显示,棕榈蓟马成虫的理论传毒效率随着饲喂健康菜椒植株时间的延长先逐渐提高最后降低。带毒棕榈蓟马成虫在健康菜椒植株上饲喂0.5 h,其理论传毒效率较低(4.55%);饲喂1~6 h,其理论传毒效率略有增长,为13.64%~22.73%,处理间无显著差异;饲喂12~48 h,其理论传毒效率进一步提高,在饲喂48 h时达最高(59.10%),处理间无显著差异;饲喂96 h,其理论传毒效率有所下降,但和48 h时无显著差异。
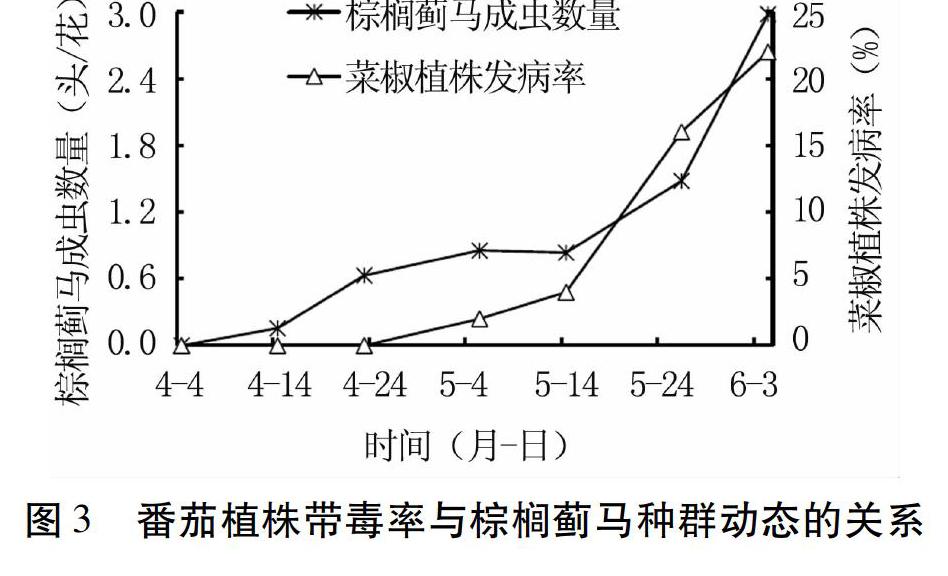
2.3 日光温室中番茄斑萎病毒病的发生与棕榈蓟马种群动态的关系
调查结果(图3)显示,调查期间日光温室菜椒植株上棕榈蓟马成虫数量呈逐渐增长趋势,4月初第一次调查时未见棕榈蓟马成虫,至5月中旬(平均单花虫量为0.86头),棕榈蓟马成虫缓慢增加,5月中旬以后棕榈蓟马成虫数量迅速增加,6月初时平均单花虫量达2.98头。从番茄斑萎病毒病发生情况来看,至4月24日调查时日光温室无带毒番茄植株,5月上旬带毒植株出现,但植株带毒率较低,至5月中旬前(4.00%)缓慢增加,而后迅速增加,至6月初番茄植株带毒率达22.00%。
从番茄斑萎病毒病发生与棕榈蓟马种群动态之间的关系来看,菜椒植株带毒率与棕榈蓟马成虫密度密切相关,且病毒病的发生具有一定滞后性:4月下旬之前,棕榈蓟马成虫数量较低,菜椒植株不带毒;4月下旬~5月中旬,棕榈蓟马成虫数量略有提高,菜椒带毒植株开始出现,但带毒率较低;5月下旬~6月初,棕榈蓟马成虫数量增长迅速,菜椒植株带毒率亦呈快速增长状态。
3 结论与讨论
番茄斑萎病毒病是我国蔬菜上的重要病毒病,继我国云南等地猖獗发生危害以来,现已在我国北方部分蔬菜产区发生。番茄斑萎病毒不能通过寄主植物自身垂直或水平传播[12, 13],而是以蓟马作为传毒介体传播。本研究结果显示,用带毒菜椒植株饲喂健康棕榈蓟马若虫24 h后,棕榈蓟马带毒率达15.00%以上,带毒棕榈蓟马成虫取食健康菜椒植株12~96 h,其理论传毒效率可达36.37%~59.10%,表明棕榈蓟马若虫具有一定获毒能力,其成虫具有一定传毒能力。本试验中虽然棕榈蓟马个体的获毒和传毒能力有限,但棕榈蓟马是菜椒、茄子、黄瓜等设施蔬菜主要害虫,该害虫在设施蔬菜上种群密度极高,成虫活动性较强,容易导致番茄斑萎病毒病的传播和蔓延[14],给我国蔬菜产业安全生产造成重大威胁。
日光温室调查结果表明,番茄斑萎病毒病的发生危害与棕榈蓟马种群动态密切相关,日光温室中番茄斑萎病毒病的发生危害程度取决于棕榈蓟马的发生数量。日光温室无棕榈蓟马或有棕榈蓟马但数量极低时,菜椒植株不带番茄斑萎病毒;当棕榈蓟马种群达到一定数量后,带毒菜椒植株开始出现;棕榈蓟马种群密度增长较快时期则是菜椒病株率迅速增长时期。因此,有效控制日光温室中棕榈蓟马的种群数量,是防治番茄斑萎病毒病的重要措施之一。
本研究初步测定了棕榈蓟马对番茄斑萎病毒的传毒特征和日光温室中番茄斑萎病毒病发生危害与棕榈蓟马种群动态的关系,但本试验是在特定环境条件下进行的,棕榈蓟马对番茄斑萎病毒获毒、传毒能力会因温度、湿度、寄主种类等因素不同而有较大差异,这些因素对棕榈蓟马传毒特征的影响有待进一步研究。另外,本研究中番茄斑萎病毒病的发生与棕榈蓟马种群动态关系是在日光温室中相对稳定条件下进行的,在生产实践中,番茄斑萎病毒病的发生还会受蔬菜种植模式、管理水平、农事操作等多种因素的影响,这些因素对番茄斑萎病毒病的危害亦有待进一步研究。
参 考 文 献:
[1] Scholtof K B G,Adkins S C, Czosnek H, et al. Top 10 plant viruses in molecular plant pathology[J]. Molecular Plant Pathology, 2011, 12(9): 938-954.
[2] 洪霓, 高必达主编. 植物病害检疫学[M]. 北京: 科学技术出版社, 2005: 120-123.
[3] 许永泽, 张宗义, 陈金香.番茄斑萎病毒TSWV广州分离物生物学特性研究[J].植物病理学报, 1989,19(4):198.
[4] 姚革. 四川晒烟上发现番茄斑萎病毒TSWV[J]. 中国烟草, 1992(4):2-4.
[5] 张仲凯, 丁铭,方琦,等. 番茄斑萎病毒属病毒(Tospovirus)在云南的发生分布研究初报[J].西南农业学报, 2004, 17(增刊): 163-168.
[6] 张仲凯, 方琦, 丁铭, 等. 侵染烟草的番茄斑萎病毒(TSWV)电镜诊断鉴定[J].电子显微镜学报, 2000, 19(3): 339-340.
[7] Dong J H, Cheng X F, Yin Y Y, et al. Characterizationg of tomato zonate spot virus, a new tosporius in China[J]. Archives of Virology, 2008, 153(5): 855-861.
[8] 程晓非, 董家红, 方琦,等.从云南蝴蝶兰上检测到番茄斑点萎蔫病毒属病毒[J].植物病理学报, 2008, 38(1):312-314.
[9] 李飞, 吴青君, 徐宝云, 等. 北京地区发现番茄斑萎病毒[J].植物保护, 2012, 38(6):186-188.
[10]Hu Z Z, Feng Z K, Zhang Z J, et.al. Complete genome sequence of a tomato spotted wilt virus isolate from China and comparison to other TSWV isolates of different geographic origin[J]. ArchVird., 2011,156(10):1905-1908.
[11]Whitfield A E, Ullman D E,German T L. Tospovirus-thrips interactions[J]. Annual Review of Phytopathology, 2005, 43:459-489.
[12]Pappu H R, Jones R A C, Jain R K.Global status of tospovirus epidemics in diverse cropping systems: successes achieved and challenges ahead[J].Virus Research,2009,141(2):219-236.
[13]Nagata T. Tissue tropism related to vector competence of Frankliniella occidentalis for tomato spotted wilt tospovirus[J].Journal of General Virology,1999,80 :507-515.
[14]Jones D R.Plant viruses transmitted by thrips[J].European Journal of Plant Pathology,2005,113: 119-157.
