
建始红椿种群结构及时间序列研究
2016-05-14 03:09冷艳芝谭艮山苏长江程德华汪洋
绿色科技 2016年6期
冷艳芝 谭艮山 苏长江 程德华 汪洋
摘要:对建始县2个不同人为干扰的红椿天然群落进行了样地调查,采用“空间替代时间法”,将林木依胸径大小分级,对种群的龄级结构、动态指数和时间序列进行了分析,结果表明:2个种群结构除第2龄级外,趋势基本一致;T1种群在4和11龄级波动,T2种群在4和8龄级出现波动,Vpi均大于50%,种群为增长型。随时间推移,低龄级存活数量虽然在相应龄级有所增加,但总体趋势递减,老龄个体逐渐增多。对现存种群及生境保护,采取适当的人工林窗措施是红椿种群恢复的关键。
关键词:建始;红椿;种群结构;数量动态
中图分类号:S718.5 文献标识码:A 文章编号:1674-9944(2016)06-0094-03
1 引言
红椿(Toona ciliata Roem)为楝科(Meliaceae)香椿属,国家二级保护濒危种,素有“中国桃花心木”之称,天然分布范围从印度经缅甸、老挝、巴基斯坦、泰国、马来西亚、印度尼西亚等国至澳大利亚。红椿主要分布于我国华南地区的低山丘陵区,东经100°16′~119°40′,北纬24°21′~30°31′。由于多年过度利用和砍伐,多数红椿天然林已经退化,有些地区已经消失。
红椿的濒危状况引起国内不少学者关注,研究领域包括红椿种质资源、遗传结构、植物生理、育种造林、化学及药理等方面,但红椿生态学研究相对不足。本文通过样地调查和相关统计分析,对红椿不同生境的种群结构动态进行了分析,并对种群未来发展做了时间序列预测,目的是探索红椿种群龄级动态特征与环境竞争的关系,为种群恢复和有效保护提供依据。
2 研究地概况
建始县属亚热带季风湿润型气候,雨量充沛,雨热同季,立体气候特征十分显著。年均气温16.O℃,年均降雨量1250mm,年均相对湿度78%,无霜期270d,年日照时数1500h。土壤为质地疏松的山地黄壤,土壤自然含水率较高。2015年在建始县青龙河村首次发现红椿天然群落。青龙河村地形复杂,红椿种群零星分布,多以16~200m2为分布规模,尺度在20m×20m的极少。选择的两个典型研究样地:夹沟(T1)样地为40年以上次生林;桃树坡(T2)样地位于山边公路旁,群落虽然被公路切断,但恢复部分保存良好。样地物种多样性丰富,除人为干扰因素以外,群落结构较为完整。两样地红椿为各自群落优势种与建群种,已经占据群落上层。
3 研究方法
3.1 样地调查
经充分踏查后,考虑不同群落类型和生境条件的代表性,尤其是人为干扰强度对红椿种群的影响程度,在不同典型群落中共设置夹沟(T1)和桃树坡(T2)两个的样地,每样地取样20m×20m。每块样地按10m×10m的大小分成4个样方,对乔木物种进行调查统计;分别在每个20m×20m样地的四角和中心位置,按5m×5m设置5个灌木层样方,再在每个灌木样方区域内按1m×1m设置5个相应草本层样方。调查内容:地形地貌、人为干扰强度、土壤、气象、郁闭度等,见表1。群落学和种群学特征:包括群落组成、高度、盖度等;对胸径(DBH)≤2.5cm的植株测定地径、高度;对胸径≥2.5cm的植株记录树高、胸径、冠幅、枝下高、分枝角等指标。
3.2 龄级划分及动态特征
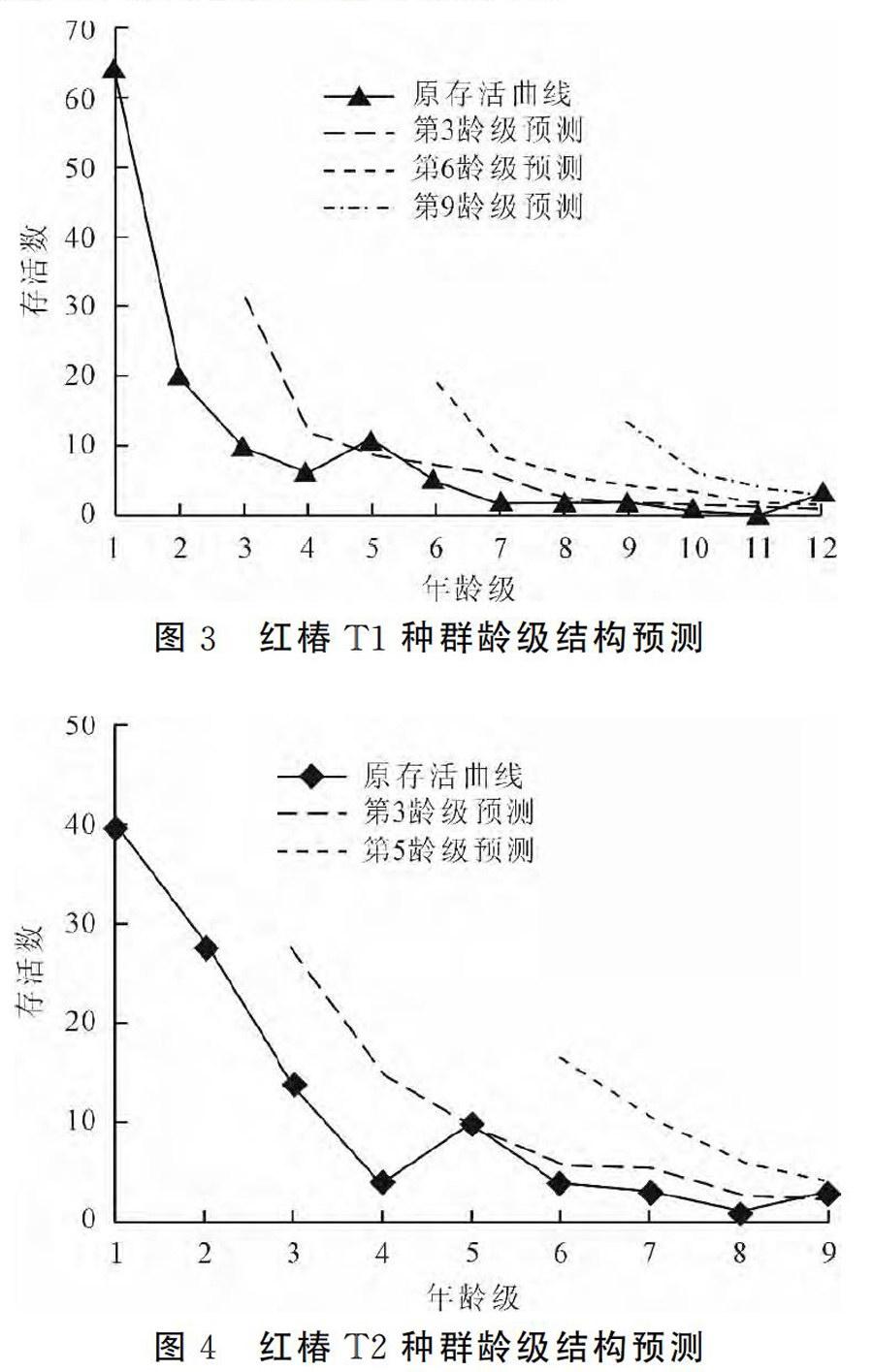
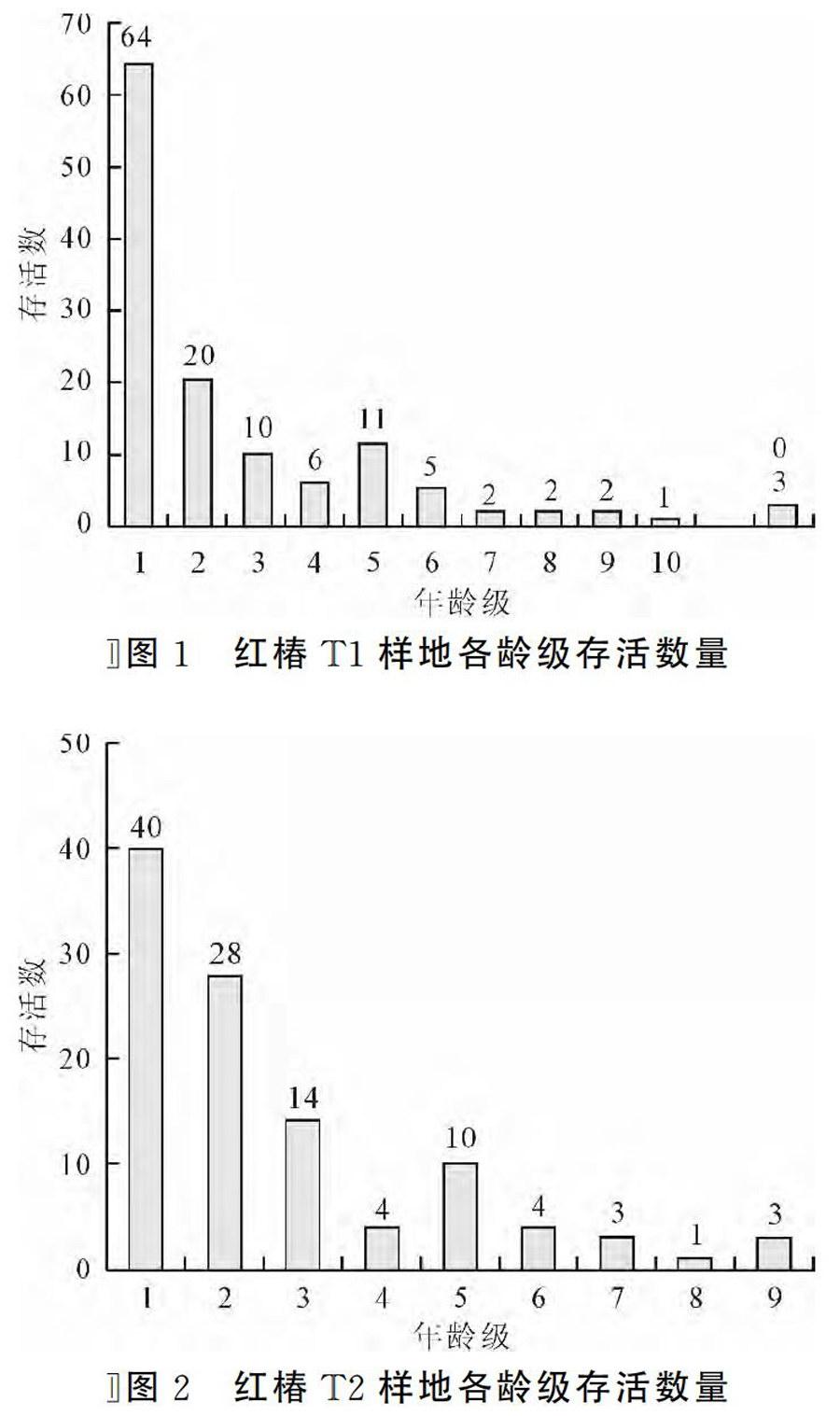
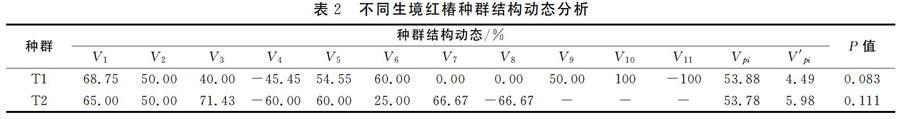
参照红椿胸径与年龄的相关性方面的研究结果,结合恩施州各地红椿实际调查数据,采用“空间替代时间法”,将林木依胸径大小分级,以立木级结构代替种群年龄结构分析种群动态,从第2径级开始,以胸径5cm为步长增加一级,即幼苗级Ⅰ(2.5cm 式中,Vn表示种群从n到n+1级的个体数量变化动态指数;Vpi表示整个种群结构的数量变化动态指数;Sn和Sn+1,分别表示第n到第n+1径级种群个体数,当考虑外部干扰时: 式中,K为种群径级数量,VpiVn取正、负、零值的意义分别反映种群或相邻径级个体数量的增长、稳定、衰退的动态关系。P为种群对外界干扰所承担的风险概率,当P的值为最大时对种群动态Vpi构成最大的影响。 4 结果与分析 4.1 红椿种群年龄结构及动态分析 图1和图2分别表示青龙河村T1和T2不同红桔群落天然种群的年龄结构。从图1和图2可以看出,该地区红椿种群年龄结构从整体上看属于增长型种群,不同的群落类型中的红椿种群的年龄结构有差别。 体现在T1种群的完整龄级结构为12级,T2种群结构仅为9级;种群T1中Ⅰ龄级个体数高于T2龄级Ⅰ龄级个体数;T1第Ⅱ龄级存活数量明显低于T2种群Ⅱ龄级数量,说明T2种群幼苗个体存活率更好,T2种群的环境条件更适应Ⅱ龄级红椿通过环境筛选和竞争。2种群其他龄级数量结构趋势基本相同。 表2为种群相邻级间个体数量的变化动态结果。T1种群在第4、8龄级出现衰退波动,其他龄级个体数量增长。整个种群年龄结构的数量变化动态指数Vpi=53.88%>0,种群属于增长型。考虑种群的外部干扰因素时,Vpi=4.49%>0,整体表现出稳定。T2种群在第4、11龄级出现衰退波动,其他龄级个体数量增长,Vpi=53.78%>0,种群属于增长型。Vpi=5.98%>O,整体表现出稳定。两个种群幼苗库较为丰富,且生殖阶段的大树占有较大比例,可以维持种群稳定。种群结构对干扰抗敏感性指数p值分别为0.083和0.111,表明红椿种群对外界干扰较为敏感,T2种群敏感度高于T1种群,在超出极限状态的干扰出现时,种群稳定存在风险。 4.2 红椿种群数量动态的时间序列预测分析 以红椿2个种群各龄级株数为原始数据,按照一次平均推移法计算出各龄级在未来12年,24年,36年后的株数(T2种群仅为24年),年龄与存活株数关系图3和图4。图中可以看出,红椿T1和T2种群各龄级株数峰值在预测序列中依次向后推移。随时间推移,低龄级存活数量虽然在相应龄级有所增加,但总体趋势递减,老龄个体逐渐增多。因此,幼龄株数的补充更显重要。最终老龄株数也急剧减少,两个种群呈现出衰老。因此,丰富的幼苗数量是维持种群更新的保证;创造红椿种群适生的林窗环境,保证第1和4龄级个体顺利通过环境筛,种群的健康发展才能持续。 5 结语 红椿龄级存活数量分析表明,存活植株与龄级之间变化关系存在着一种显著的波动规律,这种波动是种群与环境相互选择的过程。两个种群均在第4龄级表现为波动,说明红椿个体受环境制约较强,在第4龄级发展到第5龄级时,个体数量损失较大。 从群落特征分析,红椿所在群落属于常绿落叶阔叶林,红椿群落优势种。但群落以阔叶乔木为主,对空间、环境资源都具有一定的竞争要求。环境条件决定了红椿个体间以及与其它种群间对光照、水分、养分和空间生态位竞争,种群自疏作用增强,环境筛的选择强度增大,对红椿幼树及成年植株造成一定程度的损失。这与红椿龄级结构和数量动态分析结论一致。通过对建始红椿资源调查发现,与恩施州其他红椿种群相同,红椿种群基本处于在相应的群落边缘,2、4龄级的植株大多存在于群落边缘,能获得较强的光照,得以存活。因此,对现存种群及生境保护,适当的人工林窗措施是红椿种群恢复的关键。
